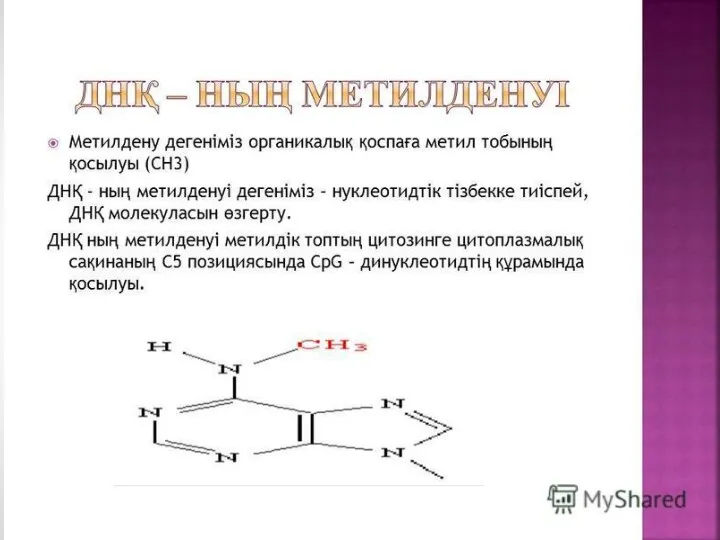
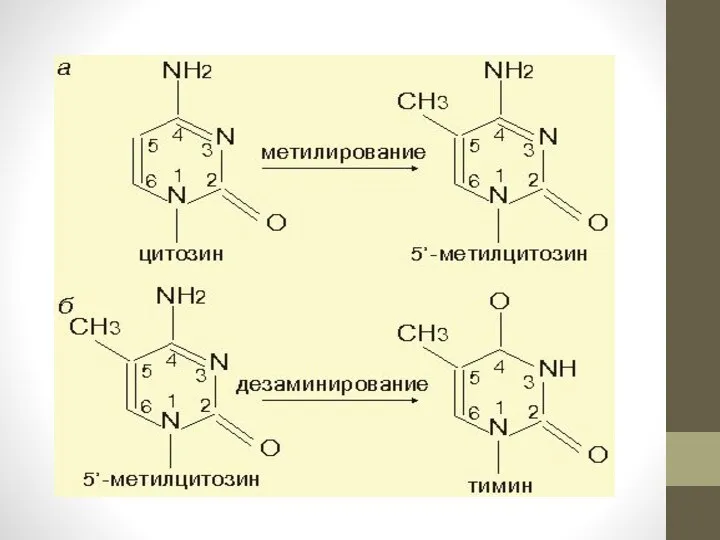
- ДНҚ-ның метилденуі

Содержание
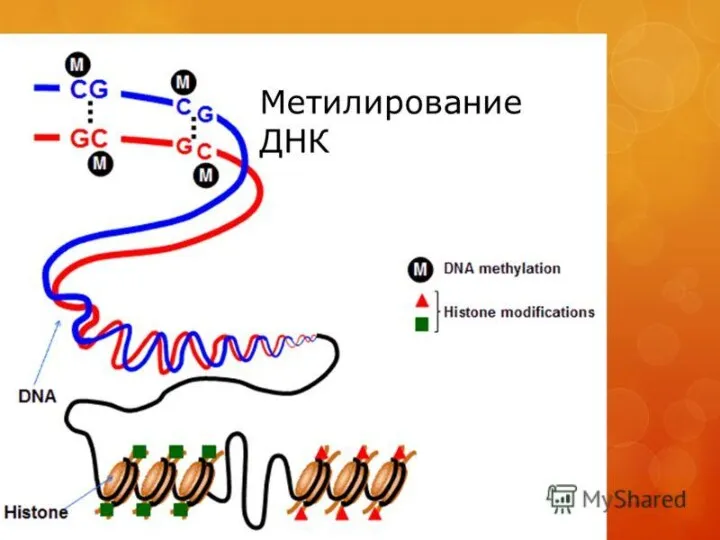
- 3. Метилденудің қызметі геннің белсенділігі мен өшірілуіне байланысты. Кейбір жағдайларда геннің промоторлық аймақтарының метилденуі ген белсенділігінің бәсеңдеуіне
- 5. У человека за процесс метилирования ДНК отвечают три фермента, называемые ДНК-метилтрансферазами 1, 3a и 3b (DNMT1,
- 7. Важными этапами в развитии злокачественных новообразований является предварительное гипометилирование ДНК[9] и последующая инактивация генов-супрессоров опухолевого роста[10].
- 9. Изменения метилирования ДНК при старении В настоящее время хорошо известно, что ландшафт метилирования геномной ДНК изменяется
- 10. Около трети всех солидных опухолей связано с мутацией гена KRAS или же с мутациями в путях
- 11. Роль метилирования в онкогенезе Сопоставление данных по генотипу людей, предрасположенных к онкологическим заболеваниям, с профилем метилирования
- 12. Пайдаланылған әдебиеттер 1.https://ru.wikipedia.org/wiki/%D0%9C%D0%B5%D1%82%D0%B8%D0%BB%D0%B8%D1%80%D0%BE%D0%B2%D0%B0%D0%BD%D0%B8%D0%B5_%D0%94%D0%9D%D0%9A 2.http://www.studfiles.ru/preview/3992254/page:24/ 3. http://medbiol.ru/medbiol/methilation/00021308.htm
- 14. Скачать презентацию

Метилденудің қызметі геннің белсенділігі мен өшірілуіне байланысты. Кейбір жағдайларда геннің промоторлық
Метилденудің қызметі геннің белсенділігі мен өшірілуіне байланысты. Кейбір жағдайларда геннің промоторлық

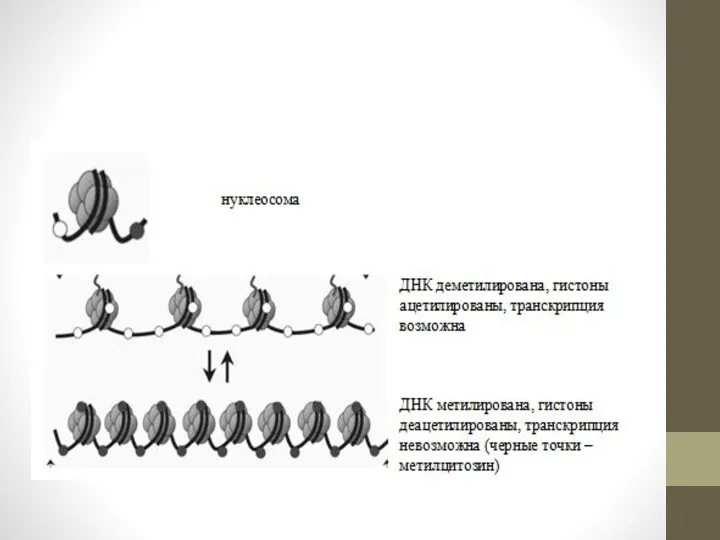
У человека за процесс метилирования ДНК отвечают три фермента, называемые ДНК-метилтрансферазами 1, 3a
У человека за процесс метилирования ДНК отвечают три фермента, называемые ДНК-метилтрансферазами 1, 3a
![Важными этапами в развитии злокачественных новообразований является предварительное гипометилирование ДНК[9]](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/228890/slide-6.jpg)
Важными этапами в развитии злокачественных новообразований является предварительное гипометилирование ДНК[9] и последующая
Важными этапами в развитии злокачественных новообразований является предварительное гипометилирование ДНК[9] и последующая

Изменения метилирования ДНК при старении
В настоящее время хорошо известно, что ландшафт
Изменения метилирования ДНК при старении
В настоящее время хорошо известно, что ландшафт

Около трети всех солидных опухолей связано с мутацией гена KRAS или
Около трети всех солидных опухолей связано с мутацией гена KRAS или

Роль метилирования в онкогенезе
Сопоставление данных по генотипу людей, предрасположенных к онкологическим
Роль метилирования в онкогенезе
Сопоставление данных по генотипу людей, предрасположенных к онкологическим

Пайдаланылған әдебиеттер
1.https://ru.wikipedia.org/wiki/%D0%9C%D0%B5%D1%82%D0%B8%D0%BB%D0%B8%D1%80%D0%BE%D0%B2%D0%B0%D0%BD%D0%B8%D0%B5_%D0%94%D0%9D%D0%9A
2.http://www.studfiles.ru/preview/3992254/page:24/
3. http://medbiol.ru/medbiol/methilation/00021308.htm
Пайдаланылған әдебиеттер
1.https://ru.wikipedia.org/wiki/%D0%9C%D0%B5%D1%82%D0%B8%D0%BB%D0%B8%D1%80%D0%BE%D0%B2%D0%B0%D0%BD%D0%B8%D0%B5_%D0%94%D0%9D%D0%9A
2.http://www.studfiles.ru/preview/3992254/page:24/
3. http://medbiol.ru/medbiol/methilation/00021308.htm
 Бүйрек физиологиясы. Несеп түзілуінің механизмі
Бүйрек физиологиясы. Несеп түзілуінің механизмі Основные загрязнители почвы
Основные загрязнители почвы Функции нейрона. Передача возбуждения в синапсах
Функции нейрона. Передача возбуждения в синапсах Строение, свойства и функции углеводов и липидов в клетке
Строение, свойства и функции углеводов и липидов в клетке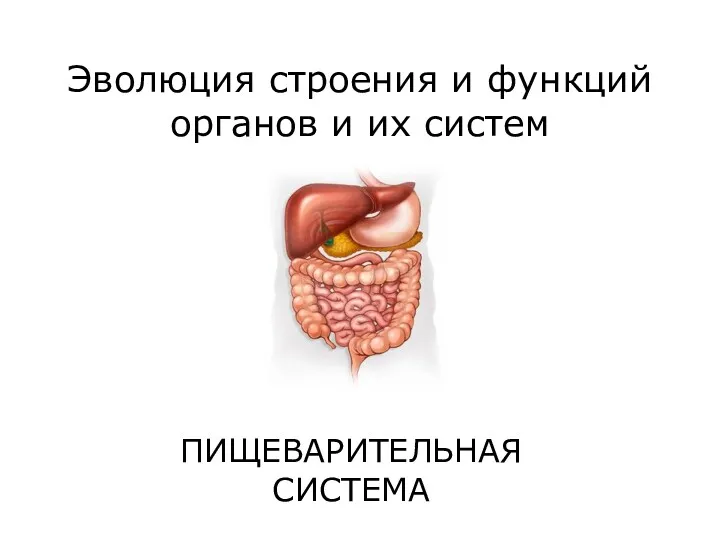 Пищеварительная система
Пищеварительная система Общее представление об организме
Общее представление об организме Презентация к уроку Эволюционное учение Ч. Дарвина
Презентация к уроку Эволюционное учение Ч. Дарвина Как звери готовятся к зиме
Как звери готовятся к зиме Приспособились ли потомки древних обитателей Земли – бактерии – к жизни на современной планете?
Приспособились ли потомки древних обитателей Земли – бактерии – к жизни на современной планете?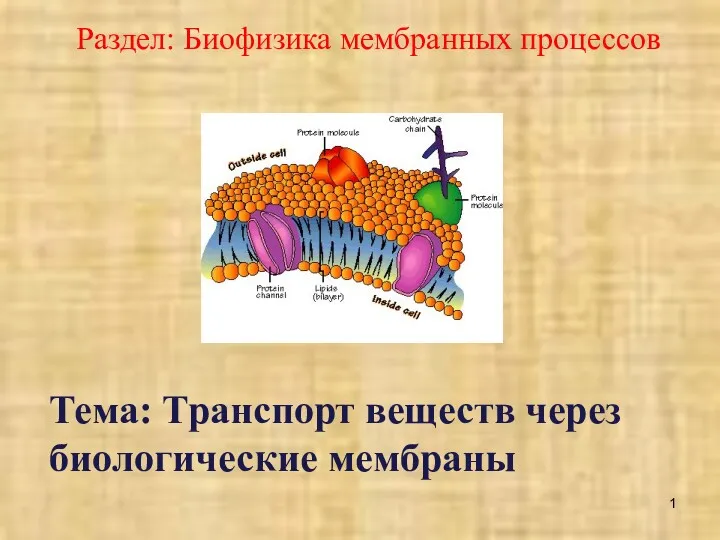 Транспорт веществ через биологические мембраны
Транспорт веществ через биологические мембраны Гидропонная система
Гидропонная система Анализирующее скрещивание. Дигибридное скрещивание. Третий закон Менделя
Анализирующее скрещивание. Дигибридное скрещивание. Третий закон Менделя Вплив електромагнітного випромінювання на рослини
Вплив електромагнітного випромінювання на рослини Презентация урока 6 класс ФГОС.
Презентация урока 6 класс ФГОС. Зимующие птицы. Подкармливание птиц зимой
Зимующие птицы. Подкармливание птиц зимой Дыхательная система человека
Дыхательная система человека Zoológico de Madrid
Zoológico de Madrid Игра Кто хочет стать отличником. Отборочный тур
Игра Кто хочет стать отличником. Отборочный тур Интересные факты о диких кошках
Интересные факты о диких кошках Тип хордовые
Тип хордовые Голонасінні. Загальна характеристика
Голонасінні. Загальна характеристика Клітинна радіобіологія. Теорія мішеней в клітинній радіобіології
Клітинна радіобіологія. Теорія мішеней в клітинній радіобіології Генетика, как наука. Взаимосвязь процессов наследственности и изменчивости
Генетика, как наука. Взаимосвязь процессов наследственности и изменчивости Информационный обзор по темам Ц. Бактерии, Ц. Грибы, Отделы растений, Вегетативные органы
Информационный обзор по темам Ц. Бактерии, Ц. Грибы, Отделы растений, Вегетативные органы Квантовые методы в медицине. Ядерный магнитный резонанс. Электронный парамагнитный резонанс. (Лекция 14)
Квантовые методы в медицине. Ядерный магнитный резонанс. Электронный парамагнитный резонанс. (Лекция 14) Органы чувств. Анализаторы
Органы чувств. Анализаторы Эмбриология (определение, виды)
Эмбриология (определение, виды) Семенные растения
Семенные растения