- Дыхание растений

Содержание
- 2. Аэробная фаза дыхания Вторая фаза дыхания — аэробная — локализована в митохондриях и требует присутствия кислорода.
- 3. Процесс можно разделить на три основные стадии: 1) окислительное декарбоксилирование пировиноградной кислоты; (превращение поступивших из цитоплазмы1)
- 4. Первая стадия — окислительное декарбоксилирование ПВК В аэробных условиях пируват симпортом с ионами Н+, движущимися по
- 5. Ацетил-SKoA Ацетил-SKoA образуется в процессе расщепления сахаров, жирных кислот и некоторых аминокислот и используется при их
- 6. Окисление ацетата дает много энергии Образующийся в ПВК-дегидрогеназной реакции ацетил-SКоА далее вступает в цикл трикарбоновых кислот
- 7. Вторая стадия - ЦТК Открытие ЦТК. В 1937 году Ганс Кребс. Эта идея родилась у него,
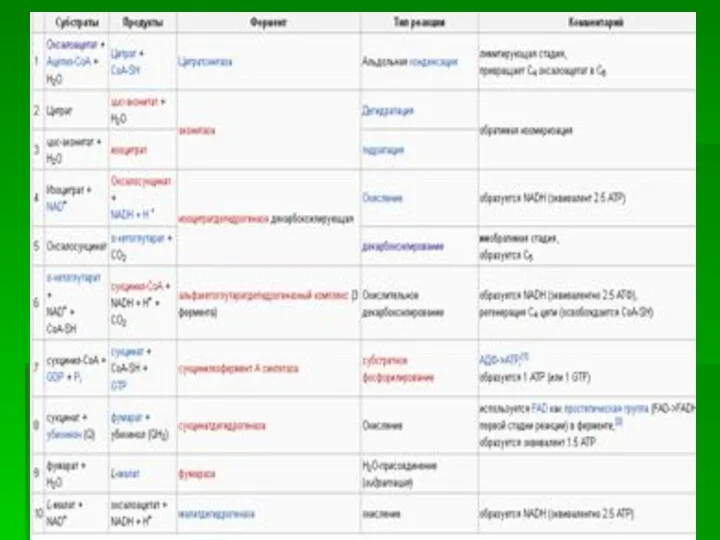
- 8. Цикл трикарбоновых кислот Цикл протекает в матриксе митохондрий и представляет собой восемь последовательных реакций. 1. Цикл
- 9. 2. Лимонная кислота превращается в изолимонную. Превращение цитрата в изоцитрат катализируется аконитазой, содержащей железо в двухвалентном
- 10. 3. Образовавшаяся а-кетоглутаровая кислота подвергается окислительному декарбоксилированию. Этот процесс также катализируется мультиферментным комплексом кетоглутаратдегидрогеназой, содержащим тиаминпирофосфат,
- 11. 3. Образовавшаяся а-кетоглутаровая кислота подвергается окислительному декарбоксилированию. Этот процесс также катализируется мультиферментным комплексом кетоглутаратдегидрогеназой, содержащим тиаминпирофосфат,
- 12. 4. Одновременно происходит восстановление еще одной молекулы НАД до НАДН и образуется сукцинил-КоА. 5. На следующем
- 13. 6. Образовавшаяся янтарная кислота окисляется до фумаровой кислоты. Реакция катализируется ферментом сукцинатдегидрогеназой, простетической группой которого является
- 14. Таким образом, ЩУК регенерирует в прежнем виде и может реагировать со следующей молекулой активного ацетата. Одновременно
- 16. Цикл трикарбоновых кислот ПВК АцКоА Цитрат Изоцитрат 2СО2 ЩЯК КГК 2СО2 2СО2 СукцинилКоА Активная янтарная к-та
- 18. Суммарное уравнение ЦТК 2СН3СОСООН + 6Н2О +8НАД +2ФАД +2АДФ +2Фн = 6СО2 +8НАДН2 +2ФАДН2 + 2АТФ
- 19. Основная роль ЦТК Основная роль ЦТК заключается в генерации атомов водорода для работы дыхательной цепи, а
- 20. Третья стадия — электронтранспортная цепь (ЭТЦ) Для чего образуются восстановленные НАД и ФАД? Являются донорами водородов
- 21. Окислительное фосфорилирование Окислительное фосфорилирование –процесс, происходящий во внутренней мембране митохондрий и заключающийся в окислении восстановленных эквивалентов
- 22. Окислительное фосфорилирование Это создает градиент концентрации ионов Н+ между цитозолем и замкнутым внутримитохондриальным пространством. Ионы водорода
- 23. РАСПОЛОЖЕНИЕ ДЫХАТЕЛЬНЫХ ПЕРЕНОСЧИКОВ В МЕМБРАНЕ МИТОХОНДРИЙ Всего цепь переноса электронов включает в себя около 40 разнообразных
- 24. Дыхательная цепь включает множество белков-переносчиков Комплекс I осуществляет перенос электронов от НАДН к убихинону. Его субстрат-
- 25. Дыхательная цепь включает множество белков-переносчиков Комплекс III переносит электроны от восстановленного убихинона к цитохрому с, т.е
- 26. СХЕМА ПЕРЕНОСА ЭЛЕКТРОНОВ ПО ДЫХАТЕЛЬНОЙ ЦЕПИ Комплекс I – НАДН-Убихинон Оксидоредуктаза Комплекс II – СукцинатДегидрогеназа Комплекс
- 28. Принцип работы дыхательной цепи Образующиеся в реакциях НАДН и ФАДН2 передают атомы водорода на ферменты дыхательной
- 29. Принцип работы дыхательной цепи Протоны Н+ стремятся обратно в матрикс и проходят через АТФ-синтазу. При этом
- 30. Строение ферментативных комплексов 1 комплекс. НАДН-КоQ-оксидоредуктаза Этот комплекс также имеет рабочее название НАДН-дегидрогеназа, содержит ФМН, 22
- 31. 3 комплекс. КоQ-цитохром с-оксидоредуктаза Данный комплекс включает цитохромы b и c1. Кроме цитохромов в нем имеются
- 32. V комплекс это фермент АТФ-синтаза, состоящий из множества белковых цепей, подразделенных на две большие группы: одна
- 33. Каким образом энергия водорода преобразуется в энергию АТФ? Механизм окислительного осфорилирования 1 Ферменты дыхательной цепи расположены
- 34. Механизм окислительного фосфорилирования 3. Здесь атомы водорода (от НАДН и ФАДН2) передают свои электроны в дыхательную
- 35. Участки сопряжения(фосфорилирования) 5 Перенос ионов водорода через мембрану (выкачивание) происходит не случайно, а в строго определенных
- 36. Участки сопряжения(фосфорилирования) В рез-те работы этих комплексов формируется градиент ионов водорода между внутренней и наружной поверхностями
- 37. Энергетические соотношения в дыхательной цепи митохондрий и участки переноса ионов Н+ через мембрану Движущей силой транспорта
- 38. Энергоноситель - АТФ АТФ образуется при дыхании путем окислительного фосфорилирования – затрачивается энергия электрона по ЭТЦ
- 39. Молекулярная модель АТФ-синтазы Структура АТФ-синтазы. Протонный канал FO и вращающаяся часть показаны синим, компонент F1 —
- 40. Механизм действия АТФ-синтазы Кристалл F1 состоит из перемежающихся α- и β-субъединиц (по 3 каждого вида), расположенных
- 41. Механизм действия АТФ-синтазы Часть компонента FO вращается, когда протоны проходят через мембрану. Это c-кольцо жестко связано
- 42. СХЕМА СИНТЕЗА АТФ ПО ХЕМИОСМОТИЧЕСКОЙ ТЕОРИИ МИТЧЕЛА
- 43. Протонная АТФ-синтаза 3 субъединицы способствуют целостности АТФ-синтазы β субъединица является основной функциональной единицей. Она имеет 3
- 44. Суммарное уравнение ЦТК 2СН3СОСООН + 6Н2О +8НАД +2ФАД +2АДФ +2Фн = 6СО2 +8НАДН2 +2ФАДН2 + 2АТФ
- 45. Значение ЦТК Цикл лимонной кислоты является механизмом, обеспечивающим улавливание большей части свободной энергии, освобождаемой в процессе
- 46. Энергетический баланс процесса дыхания суммарное уравнение гликолиза: глюкоза + 2АТФ+ 2НАД+ + 2Фн + 4АДФ→ 2
- 47. Энергетический баланс процесса дыхания Окисление одной молекулы пировиноградной кислоты сопровождается образованием ЗСО2 15 молекул АТФ. Однако
- 48. Выделение энергии: 2600 кДж - на 2 моля С3Н6О3 45% Рассеивается в виде тепла Сберегается в
- 49. Использование метаболитов ЦТК в синтезе различных соединений: 1 – 3 – синтез заменимых аминокислот; 4 –
- 51. Скачать презентацию

Аэробная фаза дыхания
Вторая фаза дыхания — аэробная — локализована в
Аэробная фаза дыхания
Вторая фаза дыхания — аэробная — локализована в

Процесс можно разделить на три основные стадии:
1) окислительное декарбоксилирование пировиноградной
Процесс можно разделить на три основные стадии:
1) окислительное декарбоксилирование пировиноградной

Первая стадия — окислительное декарбоксилирование ПВК
В аэробных условиях пируват симпортом с
Первая стадия — окислительное декарбоксилирование ПВК
В аэробных условиях пируват симпортом с
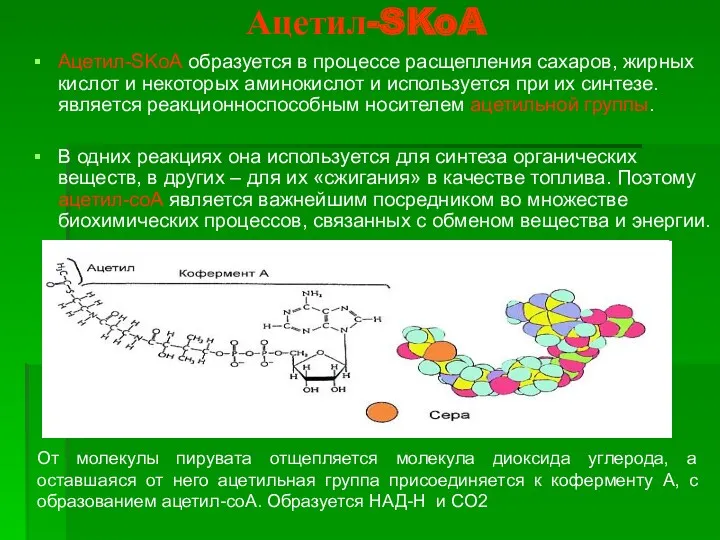
Ацетил-SKoA
Ацетил-SKoA образуется в процессе расщепления сахаров, жирных кислот и некоторых аминокислот
Ацетил-SKoA
Ацетил-SKoA образуется в процессе расщепления сахаров, жирных кислот и некоторых аминокислот

Окисление ацетата дает много энергии
Образующийся в ПВК-дегидрогеназной реакции ацетил-SКоА далее вступает
Окисление ацетата дает много энергии
Образующийся в ПВК-дегидрогеназной реакции ацетил-SКоА далее вступает

Вторая стадия - ЦТК
Открытие ЦТК.
В 1937 году Ганс Кребс. Эта
Вторая стадия - ЦТК
Открытие ЦТК.
В 1937 году Ганс Кребс. Эта

Цикл трикарбоновых кислот
Цикл протекает в матриксе митохондрий и представляет собой восемь
Цикл трикарбоновых кислот
Цикл протекает в матриксе митохондрий и представляет собой восемь

2. Лимонная кислота превращается в изолимонную.
Превращение цитрата в изоцитрат катализируется аконитазой,
2. Лимонная кислота превращается в изолимонную.
Превращение цитрата в изоцитрат катализируется аконитазой,

3. Образовавшаяся а-кетоглутаровая кислота подвергается окислительному декарбоксилированию. Этот процесс также катализируется
3. Образовавшаяся а-кетоглутаровая кислота подвергается окислительному декарбоксилированию. Этот процесс также катализируется

3. Образовавшаяся а-кетоглутаровая кислота подвергается окислительному декарбоксилированию. Этот процесс также катализируется
3. Образовавшаяся а-кетоглутаровая кислота подвергается окислительному декарбоксилированию. Этот процесс также катализируется
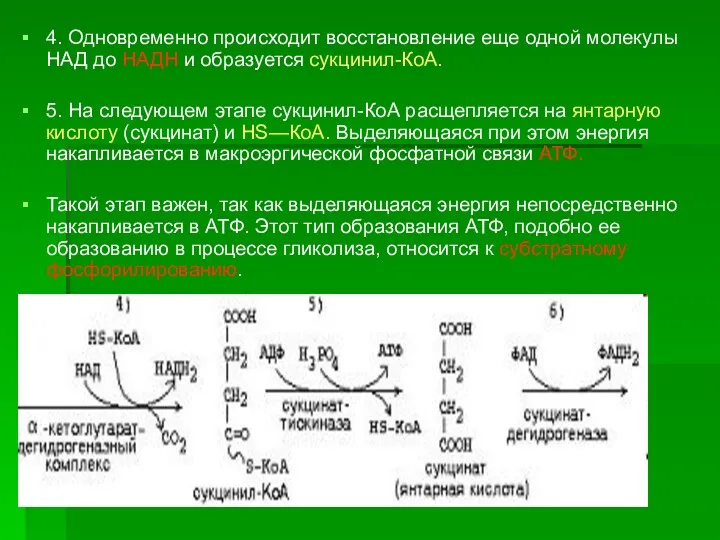
4. Одновременно происходит восстановление еще одной молекулы НАД до НАДН и
4. Одновременно происходит восстановление еще одной молекулы НАД до НАДН и

6. Образовавшаяся янтарная кислота окисляется до фумаровой кислоты. Реакция катализируется ферментом
6. Образовавшаяся янтарная кислота окисляется до фумаровой кислоты. Реакция катализируется ферментом

Таким образом, ЩУК регенерирует в прежнем виде и может реагировать со
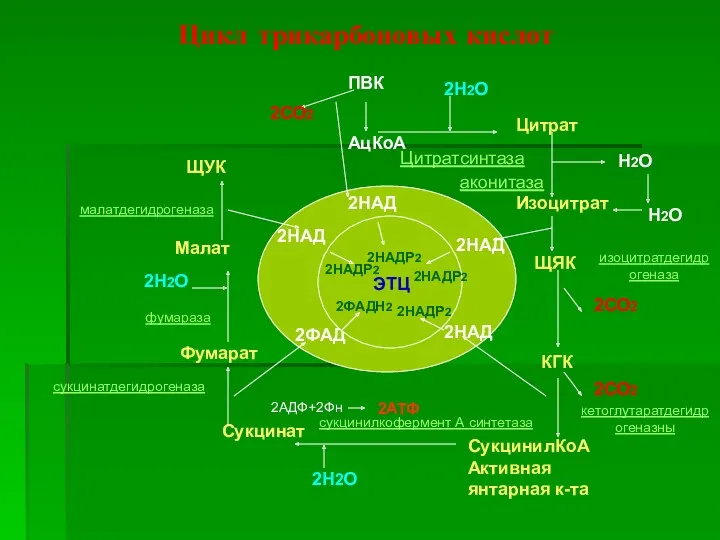
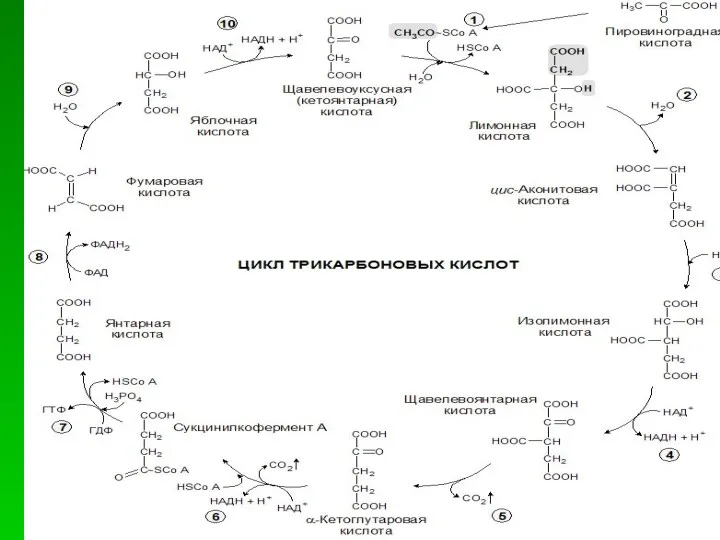
Цикл трикарбоновых кислот
ПВК
АцКоА
Цитрат
Изоцитрат
2СО2
ЩЯК
КГК
2СО2
2СО2
СукцинилКоА
Активная
янтарная к-та
Сукцинат
Фумарат
Малат
ЩУК
Н2О
Н2О
2Н2О
2Н2О
2НАД
2НАД
2НАД
2НАД
2ФАД
ЭТЦ
2НАДР2
2НАДР2
2НАДР2
2НАДР2
2ФАДН2
2Н2О
2АДФ+2Фн
2АТФ
Цитратсинтаза
аконитаза
изоцитратдегидрогеназа
кетоглутаратдегидрогеназны
сукцинилкофермент А синтетаза
Цикл трикарбоновых кислот
ПВК
АцКоА
Цитрат
Изоцитрат
2СО2
ЩЯК
КГК
2СО2
2СО2
СукцинилКоА
Активная
янтарная к-та
Сукцинат
Фумарат
Малат
ЩУК
Н2О
Н2О
2Н2О
2Н2О
2НАД
2НАД
2НАД
2НАД
2ФАД
ЭТЦ
2НАДР2
2НАДР2
2НАДР2
2НАДР2
2ФАДН2
2Н2О
2АДФ+2Фн
2АТФ
Цитратсинтаза
аконитаза
изоцитратдегидрогеназа
кетоглутаратдегидрогеназны
сукцинилкофермент А синтетаза

Суммарное уравнение ЦТК
2СН3СОСООН + 6Н2О +8НАД +2ФАД +2АДФ +2Фн =
6СО2
Суммарное уравнение ЦТК
2СН3СОСООН + 6Н2О +8НАД +2ФАД +2АДФ +2Фн =
6СО2

Основная роль ЦТК
Основная роль ЦТК заключается в
генерации атомов водорода для работы
Основная роль ЦТК
Основная роль ЦТК заключается в
генерации атомов водорода для работы

Третья стадия — электронтранспортная цепь (ЭТЦ)
Для чего образуются восстановленные НАД
Третья стадия — электронтранспортная цепь (ЭТЦ)
Для чего образуются восстановленные НАД

Окислительное фосфорилирование
Окислительное фосфорилирование –процесс, происходящий во внутренней мембране митохондрий и заключающийся
Окислительное фосфорилирование
Окислительное фосфорилирование –процесс, происходящий во внутренней мембране митохондрий и заключающийся

Окислительное фосфорилирование
Это создает градиент концентрации ионов Н+ между цитозолем и замкнутым
Окислительное фосфорилирование
Это создает градиент концентрации ионов Н+ между цитозолем и замкнутым
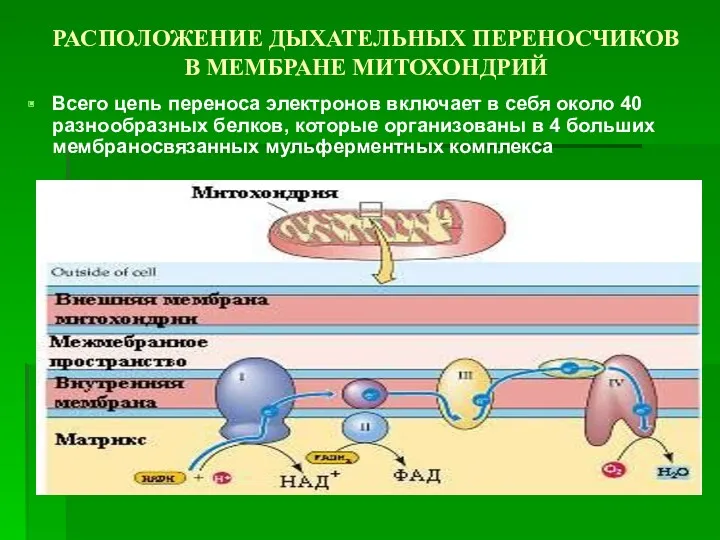
РАСПОЛОЖЕНИЕ ДЫХАТЕЛЬНЫХ ПЕРЕНОСЧИКОВ В МЕМБРАНЕ МИТОХОНДРИЙ
Всего цепь переноса электронов включает в
РАСПОЛОЖЕНИЕ ДЫХАТЕЛЬНЫХ ПЕРЕНОСЧИКОВ В МЕМБРАНЕ МИТОХОНДРИЙ
Всего цепь переноса электронов включает в
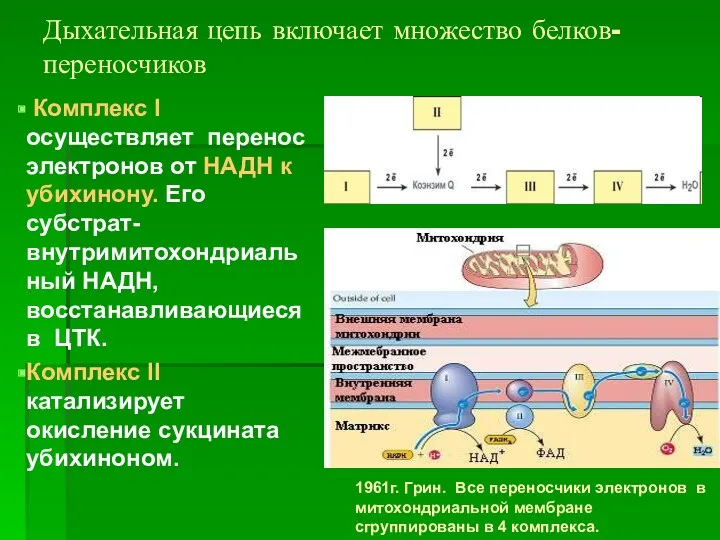
Дыхательная цепь включает множество белков-переносчиков
Комплекс I осуществляет перенос электронов от
Дыхательная цепь включает множество белков-переносчиков
Комплекс I осуществляет перенос электронов от
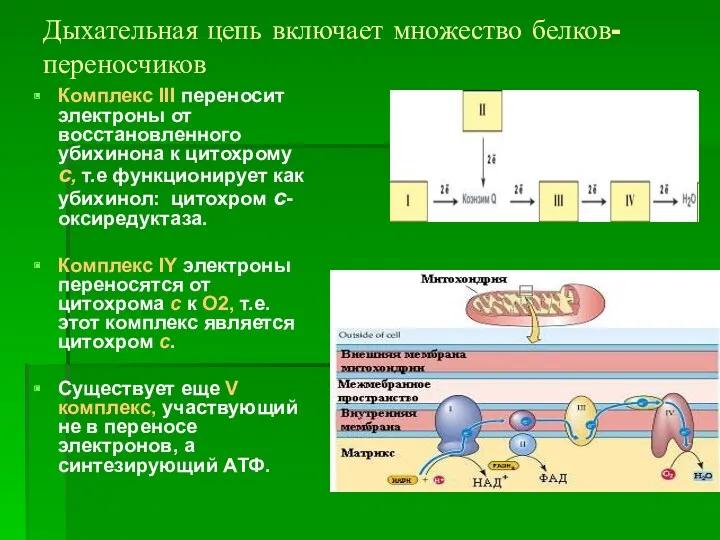
Дыхательная цепь включает множество белков-переносчиков
Комплекс III переносит электроны от восстановленного убихинона
Дыхательная цепь включает множество белков-переносчиков
Комплекс III переносит электроны от восстановленного убихинона
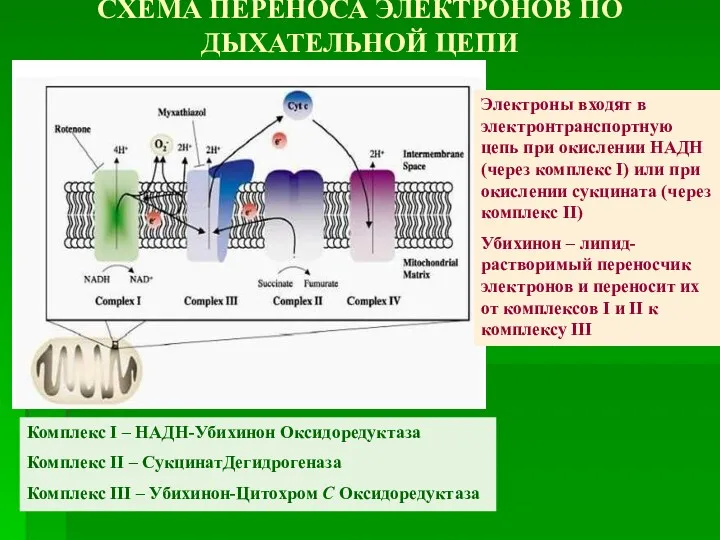
СХЕМА ПЕРЕНОСА ЭЛЕКТРОНОВ ПО ДЫХАТЕЛЬНОЙ ЦЕПИ
Комплекс I – НАДН-Убихинон Оксидоредуктаза
Комплекс II
СХЕМА ПЕРЕНОСА ЭЛЕКТРОНОВ ПО ДЫХАТЕЛЬНОЙ ЦЕПИ
Комплекс I – НАДН-Убихинон Оксидоредуктаза
Комплекс II
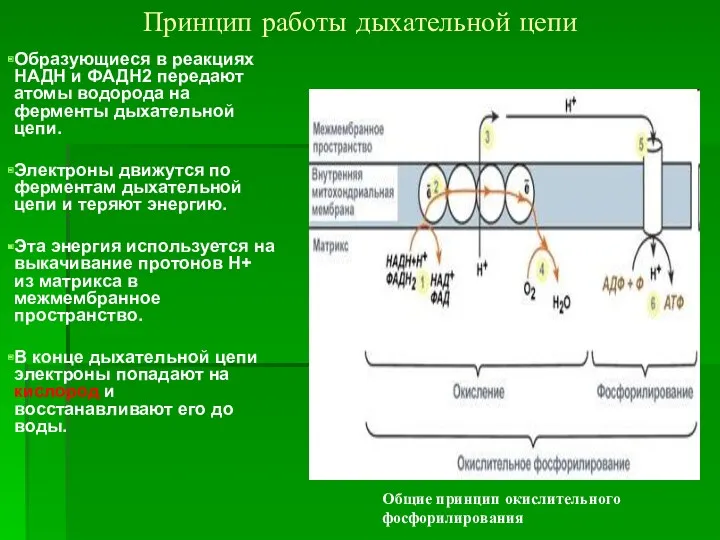
Принцип работы дыхательной цепи
Образующиеся в реакциях НАДН и ФАДН2 передают атомы
Принцип работы дыхательной цепи
Образующиеся в реакциях НАДН и ФАДН2 передают атомы

Принцип работы дыхательной цепи
Протоны Н+ стремятся обратно в матрикс и проходят
Принцип работы дыхательной цепи
Протоны Н+ стремятся обратно в матрикс и проходят

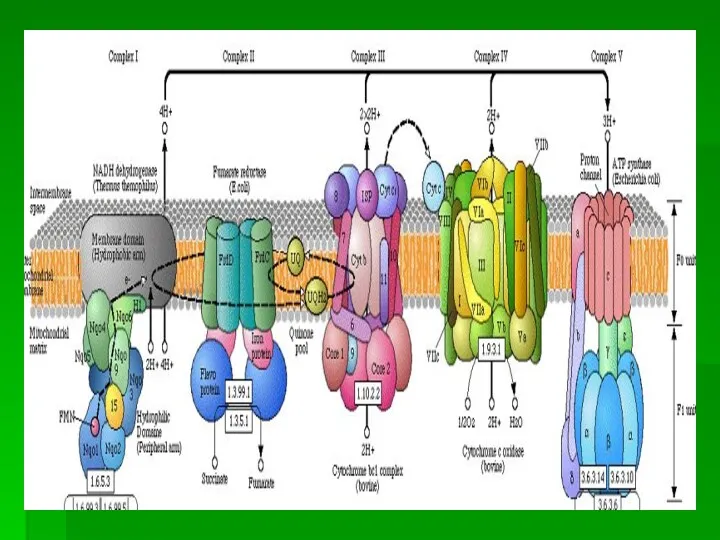
Строение ферментативных комплексов
1 комплекс. НАДН-КоQ-оксидоредуктаза
Этот комплекс также имеет рабочее название НАДН-дегидрогеназа, содержит ФМН,
Строение ферментативных комплексов
1 комплекс. НАДН-КоQ-оксидоредуктаза
Этот комплекс также имеет рабочее название НАДН-дегидрогеназа, содержит ФМН,

3 комплекс. КоQ-цитохром с-оксидоредуктаза
Данный комплекс включает цитохромы b и c1.
Кроме
3 комплекс. КоQ-цитохром с-оксидоредуктаза
Данный комплекс включает цитохромы b и c1.
Кроме

V комплекс
это фермент АТФ-синтаза, состоящий из множества белковых цепей, подразделенных на
V комплекс
это фермент АТФ-синтаза, состоящий из множества белковых цепей, подразделенных на

Каким образом энергия водорода преобразуется в энергию АТФ? Механизм окислительного
Каким образом энергия водорода преобразуется в энергию АТФ? Механизм окислительного

Механизм окислительного фосфорилирования
3. Здесь атомы водорода (от НАДН и ФАДН2) передают
Механизм окислительного фосфорилирования
3. Здесь атомы водорода (от НАДН и ФАДН2) передают
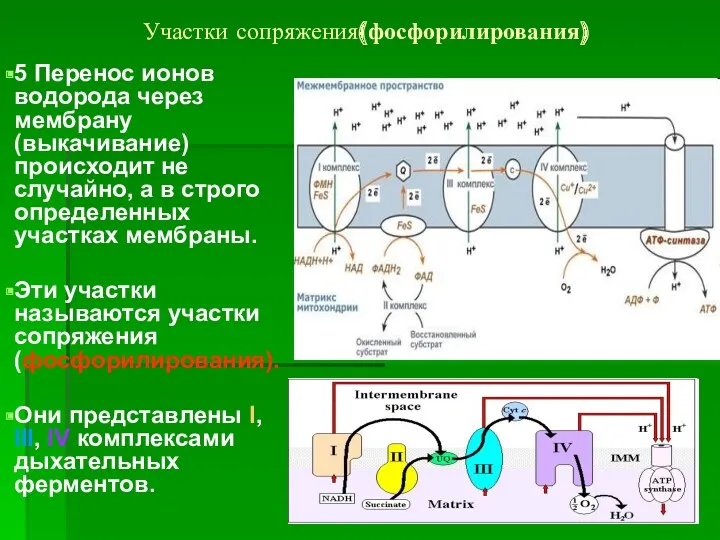
Участки сопряжения(фосфорилирования)
5 Перенос ионов водорода через мембрану (выкачивание) происходит не случайно,
Участки сопряжения(фосфорилирования)
5 Перенос ионов водорода через мембрану (выкачивание) происходит не случайно,
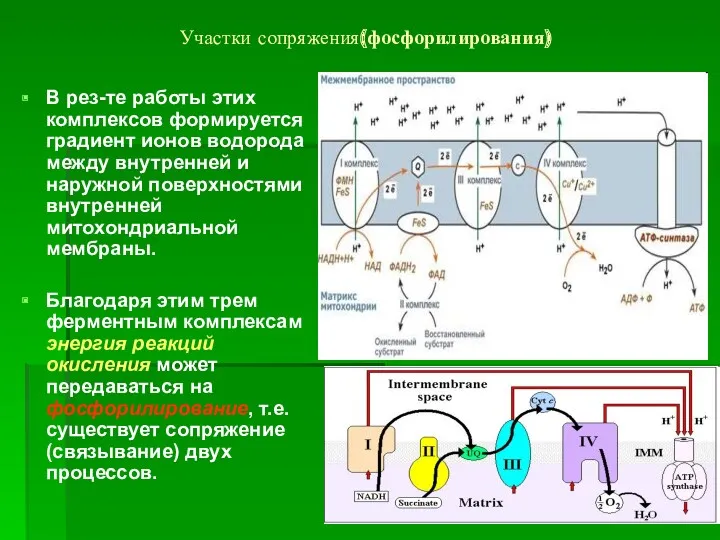
Участки сопряжения(фосфорилирования)
В рез-те работы этих комплексов формируется градиент ионов водорода между
Участки сопряжения(фосфорилирования)
В рез-те работы этих комплексов формируется градиент ионов водорода между
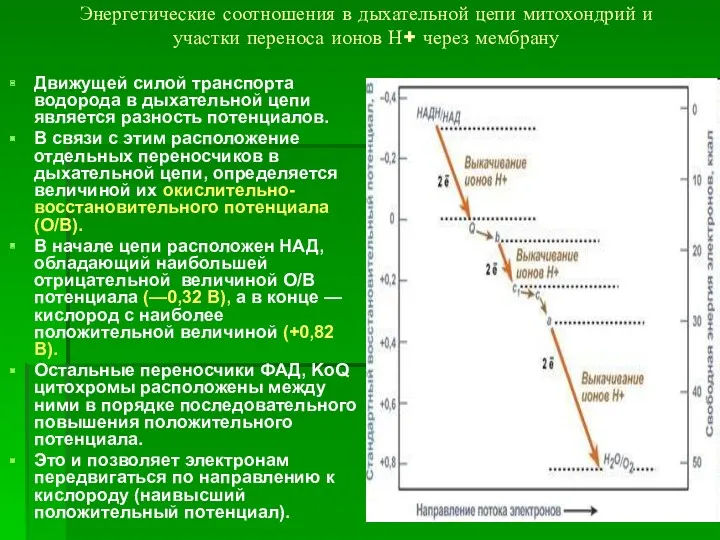
Энергетические соотношения в дыхательной цепи митохондрий и участки переноса ионов Н+
Энергетические соотношения в дыхательной цепи митохондрий и участки переноса ионов Н+
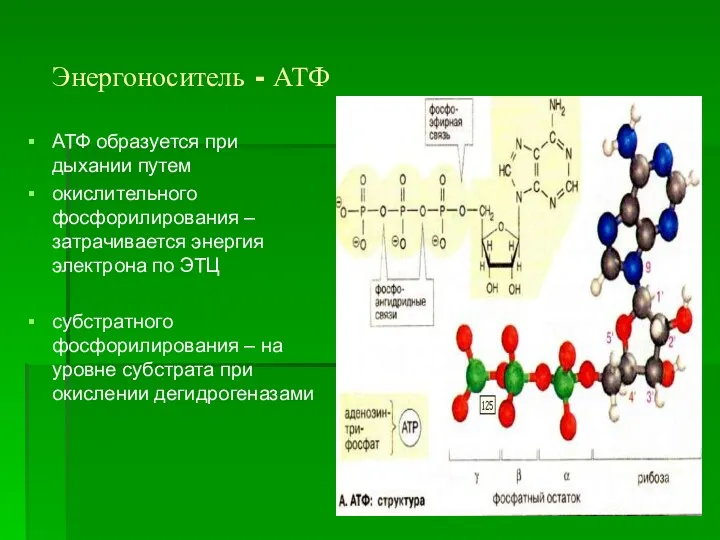
Энергоноситель - АТФ
АТФ образуется при дыхании путем
окислительного фосфорилирования –
Энергоноситель - АТФ
АТФ образуется при дыхании путем
окислительного фосфорилирования –
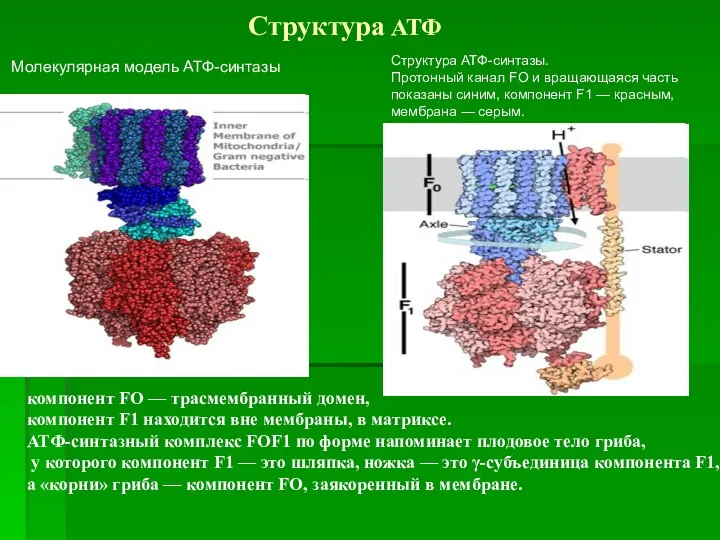
Молекулярная модель АТФ-синтазы
Структура АТФ-синтазы.
Протонный канал FO и вращающаяся часть
Молекулярная модель АТФ-синтазы
Структура АТФ-синтазы.
Протонный канал FO и вращающаяся часть
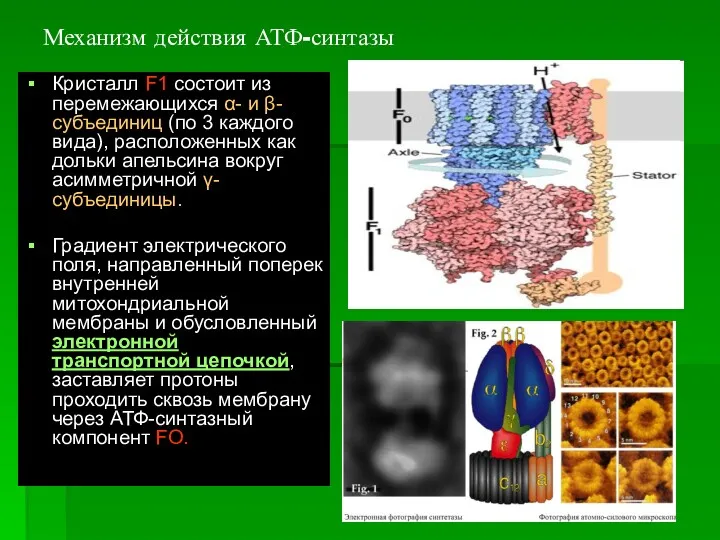
Механизм действия АТФ-синтазы
Кристалл F1 состоит из перемежающихся α- и β-субъединиц (по
Механизм действия АТФ-синтазы
Кристалл F1 состоит из перемежающихся α- и β-субъединиц (по

Механизм действия АТФ-синтазы
Часть компонента FO вращается, когда протоны проходят через мембрану.
Механизм действия АТФ-синтазы
Часть компонента FO вращается, когда протоны проходят через мембрану.
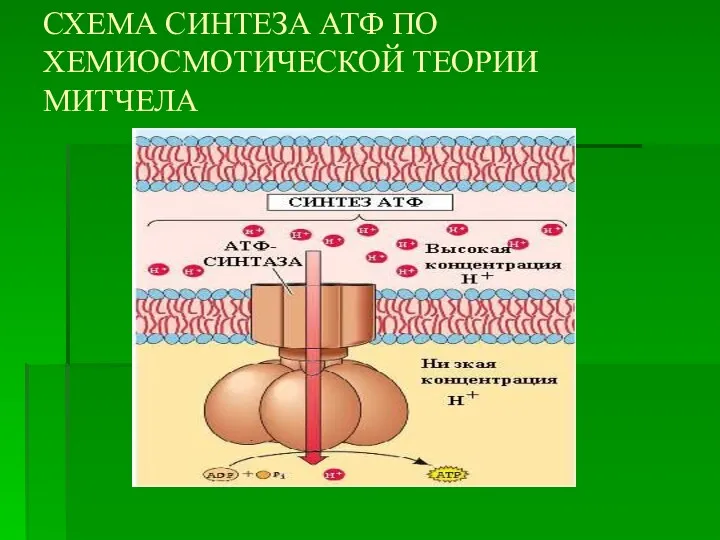
СХЕМА СИНТЕЗА АТФ ПО ХЕМИОСМОТИЧЕСКОЙ ТЕОРИИ МИТЧЕЛА
СХЕМА СИНТЕЗА АТФ ПО ХЕМИОСМОТИЧЕСКОЙ ТЕОРИИ МИТЧЕЛА

Протонная АТФ-синтаза
3 субъединицы способствуют целостности АТФ-синтазы
β субъединица является основной
Протонная АТФ-синтаза
3 субъединицы способствуют целостности АТФ-синтазы
β субъединица является основной

Суммарное уравнение ЦТК
2СН3СОСООН + 6Н2О +8НАД +2ФАД +2АДФ +2Фн =
6СО2
Суммарное уравнение ЦТК
2СН3СОСООН + 6Н2О +8НАД +2ФАД +2АДФ +2Фн =
6СО2

Значение ЦТК
Цикл лимонной кислоты является механизмом, обеспечивающим улавливание большей части свободной
Значение ЦТК
Цикл лимонной кислоты является механизмом, обеспечивающим улавливание большей части свободной

Энергетический баланс процесса дыхания
суммарное уравнение гликолиза:
глюкоза + 2АТФ+ 2НАД+ +
Энергетический баланс процесса дыхания
суммарное уравнение гликолиза:
глюкоза + 2АТФ+ 2НАД+ +

Энергетический баланс процесса дыхания
Окисление одной молекулы пировиноградной кислоты сопровождается образованием
Энергетический баланс процесса дыхания
Окисление одной молекулы пировиноградной кислоты сопровождается образованием
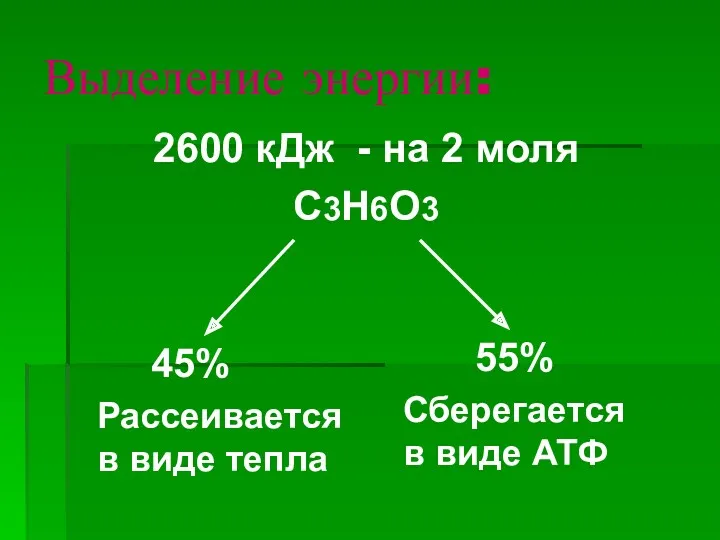
Выделение энергии:
2600 кДж - на 2 моля
С3Н6О3
45%
Рассеивается
в виде тепла
Сберегается
в
Выделение энергии:
2600 кДж - на 2 моля
С3Н6О3
45%
Рассеивается
в виде тепла
Сберегается
в
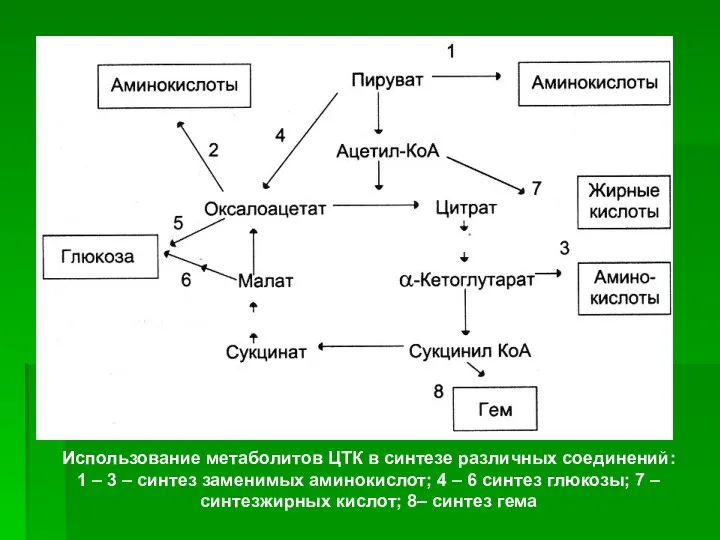
Использование метаболитов ЦТК в синтезе различных соединений:
1 – 3 – синтез
Использование метаболитов ЦТК в синтезе различных соединений:
1 – 3 – синтез
 Морфо-фізіологічні ознаки ссавців: кровоносна та нервова системи
Морфо-фізіологічні ознаки ссавців: кровоносна та нервова системи Национальный парк Зов Тигра
Национальный парк Зов Тигра Основы генетики
Основы генетики 20231204_35.obobshchenie_po_teme_teoriya_evolyutsii
20231204_35.obobshchenie_po_teme_teoriya_evolyutsii Размножение. Начало новой жизни
Размножение. Начало новой жизни Скелет верхних и нижних конечностей
Скелет верхних и нижних конечностей Окружающий мир
Окружающий мир Селекционные линии и перспективные гибриды картофеля
Селекционные линии и перспективные гибриды картофеля Как паук костюмчик менял
Как паук костюмчик менял Лекция № 1 Наука экология биосфера
Лекция № 1 Наука экология биосфера Умеют ли животные общаться
Умеют ли животные общаться Великий круговорот жизни. Урок окружающего мира
Великий круговорот жизни. Урок окружающего мира Витамины - кофакторы ферментативных реакций. Классификация витаминов. Гиповитаминозы. Гипервитаминозы. Антивитамины
Витамины - кофакторы ферментативных реакций. Классификация витаминов. Гиповитаминозы. Гипервитаминозы. Антивитамины Воробьиные (часть 1). Passeriformes
Воробьиные (часть 1). Passeriformes Весенние изменения в жизни зверей
Весенние изменения в жизни зверей Соматосенсорлы жүйенің физиологиясы
Соматосенсорлы жүйенің физиологиясы Карты генов наследственных заболеваний
Карты генов наследственных заболеваний Различия в строении клеток эукариот и прокариот
Различия в строении клеток эукариот и прокариот Катаболизмнің жалпы жолдары
Катаболизмнің жалпы жолдары Життя риб навесні. 2 клас
Життя риб навесні. 2 клас Жердегі биологиялық эволюцияның кезеңдері мен негіздері
Жердегі биологиялық эволюцияның кезеңдері мен негіздері Биологические основы физической культуры и спорта
Биологические основы физической культуры и спорта Класс брюхоногие моллюски или улитки
Класс брюхоногие моллюски или улитки Ткани растений
Ткани растений Міні-проект Рослини-мандрівники
Міні-проект Рослини-мандрівники Чудо-доктора. Лекарственные растения Лутугинского района
Чудо-доктора. Лекарственные растения Лутугинского района Цитоплазма и органеллы
Цитоплазма и органеллы Эндокринные железы
Эндокринные железы