- Катаболизмнің жалпы жолдары

Содержание
- 2. Основные вопросы лекции: Макроэргтік қосылыстар.Тіндік тыныс алу боитотығу туралы түсінік Тотығып фосфорлану, Р/0 коэффициенті және АДФ/О.Тіндік
- 3. Специфические и общие пути катаболизма . Катаболитикалық процесстер екіге бөлінеді: - катаболизмнің өзіндік жолы, барлық класстарға
- 5. Пирожүзім қышқылының тотығып декарбоксильденуі Пирожүзім қышқылының тотығып декарбоксильденуі митохондрияның ішкі мембрананасында орналасқан мультиэнзимді пируват-дегидрогеназды комплекспен катализденеді.
- 6. катаболизмнің жалпы жолына кіреді: - Пирожүзім қышқылының тотығып декарбоксильденуі - Үш карбон қышқылы циклінде ацетил-КоА айналуы
- 7. Пируваттың тотығуы
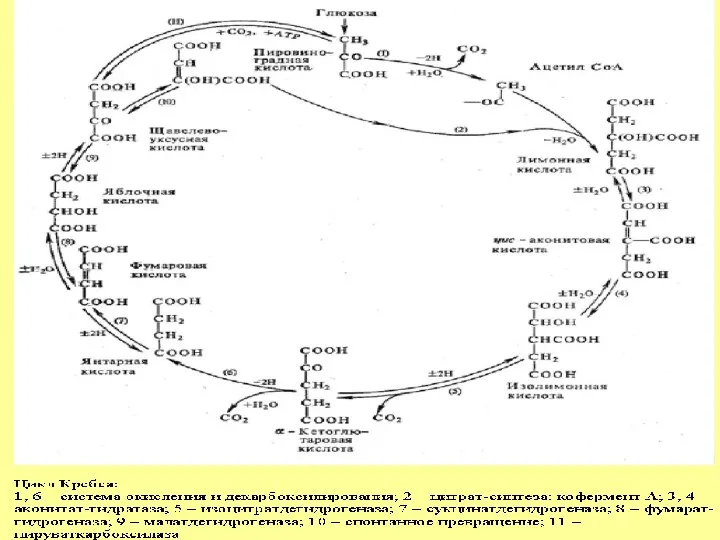
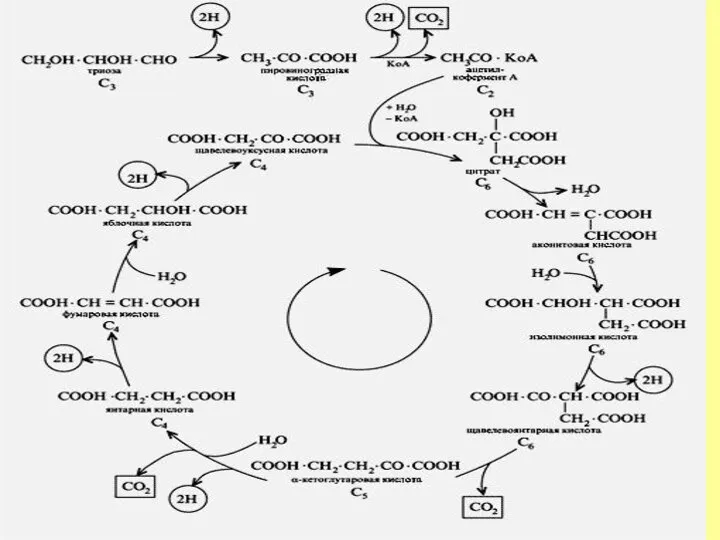
- 11. Үш карбон қышқылы циклі
- 12. Цикл трикарбоновых кислот
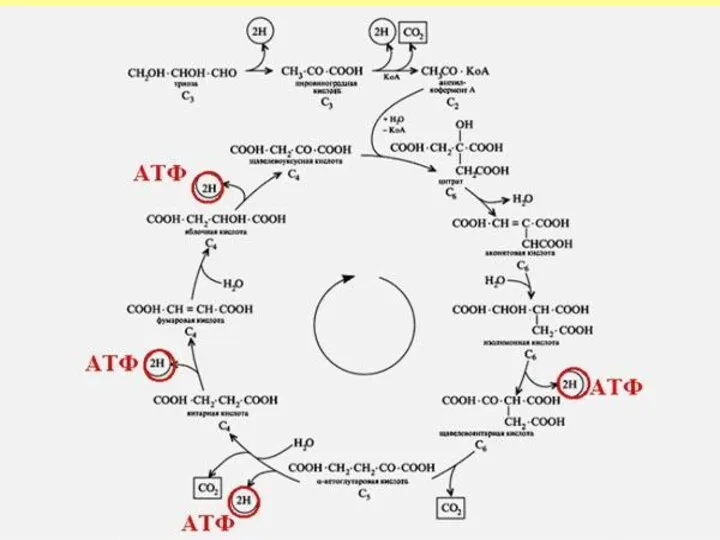
- 14. Энергия шығымы Дегидрогеназалармен төрт реакцияда субстрат дегидрирленуі жүреді изоцитратдегидрогеназа, кофермент НАД, альфа-кетоглутаратдегидрогеназалар, кофактор НАД сукцинатдегидрогеназа, кофермент
- 18. Челночные механизмы транспорта Своеобразным вариантом групповой транслокации является механизм переноса протонов (Н+) через митохондриальные мембраны. Мембраны
- 19. Глицерофосфатный челнок
- 20. Малатный челнок
- 21. Основные типы окисления Реакции катаболизма органических соединений (субстратов), сопряженные с высвобождением свободной энергии в биосистемах, в
- 22. Маркоэрги К энергетическим субстратам для клеток животного организма относятся углеводы, липиды и аминокислоты. Удельная калорийность углеводов
- 23. Механизмы трансформация энергии в клетке Высвобождение свободной энергии при катаболизме углеводов, липидов и аминокислот может происходить
- 25. Механизм биологического окисления
- 26. Основной функцией этого процесса является обеспечение организма энергией в доступной для использования форме (прежде всего в
- 27. Анаэробные дегидрогеназы Эти ферменты катализируют реакции переноса атомов водорода (протоны и электроны) на любой акцептор кроме
- 28. N o H OH OH H CH2 O P OH O O P OH O O
- 29. АЭРОБНЫЕ ДЕГИДРОГЕНАЗЫ Эти ферменты катализируют реакции переноса атомов водорода (протоны и электроны) на любой акцептор в
- 30. С С С С С С С С С С С С H H H3 H3
- 31. Кофермент Q Компонент дыхательной цепи – убихинон, производный бензохинона является коферментом ФАДН2 –дегидрогеназ, катализирующих перенос протонов
- 32. ЦИТОХРОМЫ Дальнейший перенос электронов от убихинона на кислород осуществляет система цитохромов: b, c1, c, a- a3.
- 33. Механизм действия цитохромов в окислительно-восстановительных реакциях Fe3+ + e- Fe2+ Fe2+ - e - Fe3+ Cu2+
- 34. СТРУКТУРА ДЫХАТЕЛЬНОЙ ЦЕПИ Перенос электронов и протонов в митохондриальной дыхательной цепи происходит по градиенту их окислительно-восстановительного
- 35. ДЫХАТЕЛЬНАЯ ЦЕПЬ SH2 НАД ФАД КоQ b c1 c a a3 1/2O2 O + H Q
- 36. ДЫХАТЕЛЬНАЯ ЦЕПЬ И РЕДОКС ПОТЕНЦИАЛ
- 37. ПУНКТЫ ФОСФОРИЛИРОВАНИЯ Первый пункт фосфорилирования находится между ферментами НАД и ФАД, где разность их потенциала равна
- 38. Ко-А-производные жирных кислот Изоцитрат Малат Пируват Глутамат НАД ФП b КоQ АТФ АДФ - Аминобарбитал, ротенон
- 39. Окислительное фосфорилирование При окислении одной молекулы субстрата на один атом поглощенного митохондриями кислорода может использоваться от
- 41. Согласно предложенной П. Митчелом гипотезе, движущей силой фосфорилирования АДФ служит энергия разности редокс-потенциалов, возникающая при переносе
- 43. Скачать презентацию

Основные вопросы лекции:
Макроэргтік қосылыстар.Тіндік тыныс алу боитотығу туралы түсінік
Тотығып фосфорлану,
Основные вопросы лекции:
Макроэргтік қосылыстар.Тіндік тыныс алу боитотығу туралы түсінік
Тотығып фосфорлану,

Специфические и общие пути катаболизма
. Катаболитикалық процесстер екіге бөлінеді:
-
Специфические и общие пути катаболизма
. Катаболитикалық процесстер екіге бөлінеді:
-

Пирожүзім қышқылының тотығып декарбоксильденуі
Пирожүзім қышқылының тотығып декарбоксильденуі митохондрияның ішкі
Пирожүзім қышқылының тотығып декарбоксильденуі
Пирожүзім қышқылының тотығып декарбоксильденуі митохондрияның ішкі

катаболизмнің жалпы жолына кіреді:
- Пирожүзім қышқылының тотығып декарбоксильденуі
-
катаболизмнің жалпы жолына кіреді:
- Пирожүзім қышқылының тотығып декарбоксильденуі
-
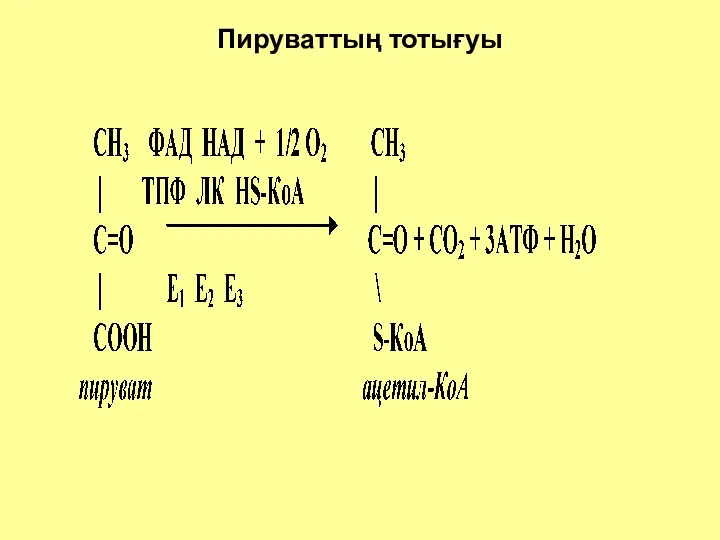
Пируваттың тотығуы
Пируваттың тотығуы




Үш карбон қышқылы циклі
Үш карбон қышқылы циклі

Цикл трикарбоновых кислот
Цикл трикарбоновых кислот


Энергия шығымы
Дегидрогеназалармен төрт реакцияда субстрат дегидрирленуі жүреді
изоцитратдегидрогеназа, кофермент НАД,
альфа-кетоглутаратдегидрогеназалар, кофактор НАД
сукцинатдегидрогеназа,
Энергия шығымы
Дегидрогеназалармен төрт реакцияда субстрат дегидрирленуі жүреді
изоцитратдегидрогеназа, кофермент НАД,
альфа-кетоглутаратдегидрогеназалар, кофактор НАД
сукцинатдегидрогеназа,
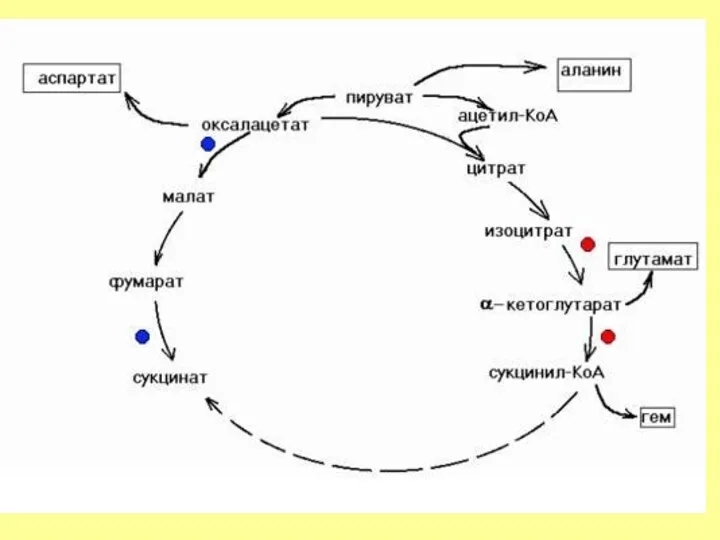

Челночные механизмы транспорта
Своеобразным вариантом групповой транслокации является механизм переноса протонов (Н+)
Челночные механизмы транспорта
Своеобразным вариантом групповой транслокации является механизм переноса протонов (Н+)
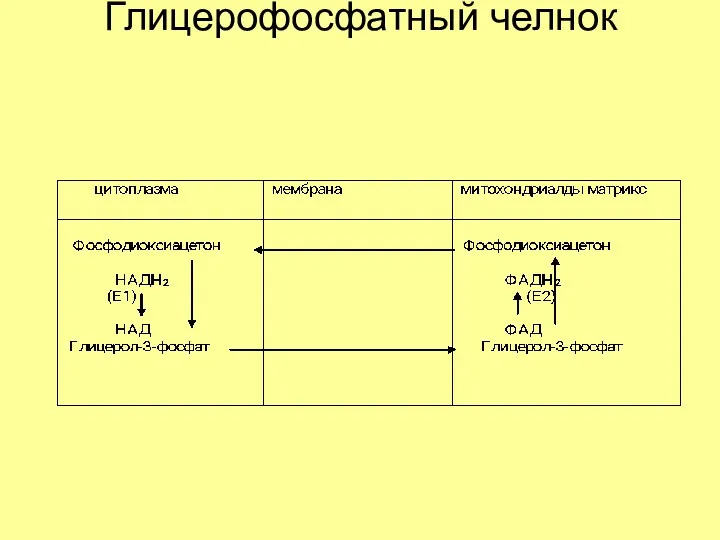
Глицерофосфатный челнок
Глицерофосфатный челнок
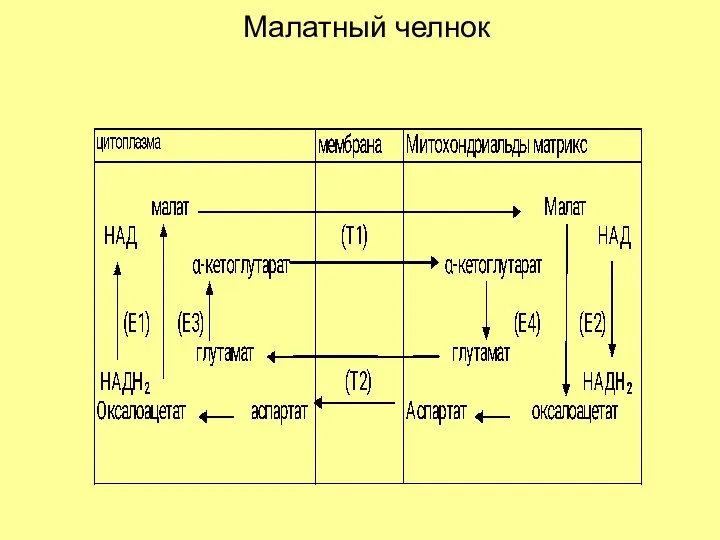
Малатный челнок
Малатный челнок

Основные типы окисления
Реакции катаболизма органических соединений (субстратов), сопряженные с высвобождением
Основные типы окисления
Реакции катаболизма органических соединений (субстратов), сопряженные с высвобождением

Маркоэрги
К энергетическим субстратам для клеток животного организма относятся углеводы, липиды и
Маркоэрги
К энергетическим субстратам для клеток животного организма относятся углеводы, липиды и

Механизмы трансформация энергии в клетке
Высвобождение свободной энергии при катаболизме углеводов,
Механизмы трансформация энергии в клетке
Высвобождение свободной энергии при катаболизме углеводов,
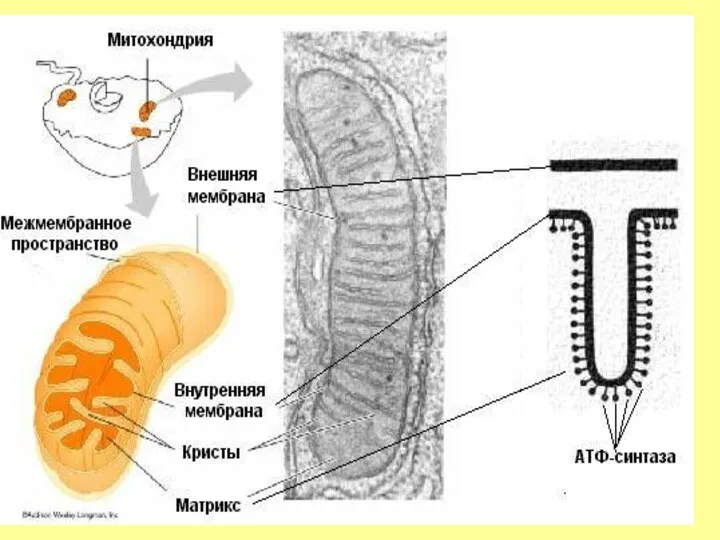

Механизм биологического окисления
Механизм биологического окисления

Основной функцией этого процесса является обеспечение организма энергией в доступной для
Основной функцией этого процесса является обеспечение организма энергией в доступной для

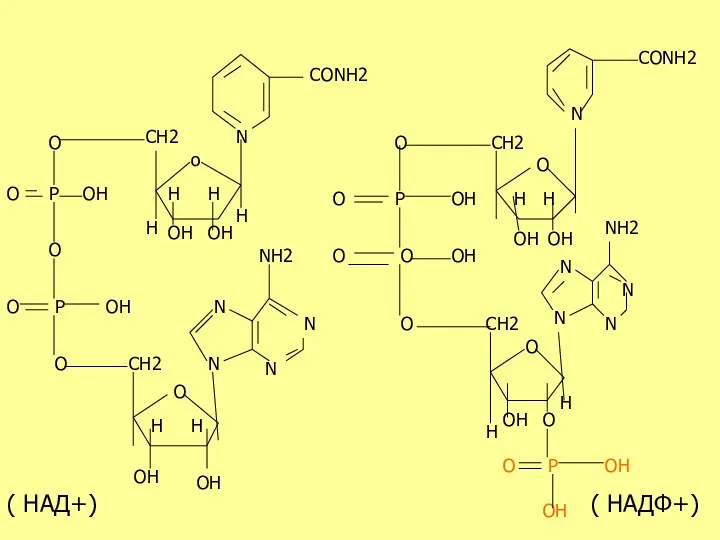
Анаэробные дегидрогеназы
Эти ферменты катализируют реакции переноса атомов водорода (протоны и электроны)
Анаэробные дегидрогеназы
Эти ферменты катализируют реакции переноса атомов водорода (протоны и электроны)
N
o
H
OH
OH
H
CH2
O
P
OH
O
O
P
OH
O
O
CH2
O
OH
OH
N
N
N
N
NH2
H
H
H
H
CONH2
( НАД+)
O
P
O
OH
O
O
OH
O
CH2
H
CH2
CONH2
N
O
NH2
N
N
N
N
H
OH
O
OH
OH
H
H
O
P
OH
O
OH
( НАДФ+)
N
o
H
OH
OH
H
CH2
O
P
OH
O
O
P
OH
O
O
CH2
O
OH
OH
N
N
N
N
NH2
H
H
H
H
CONH2
( НАД+)
O
P
O
OH
O
O
OH
O
CH2
H
CH2
CONH2
N
O
NH2
N
N
N
N
H
OH
O
OH
OH
H
H
O
P
OH
O
OH
( НАДФ+)

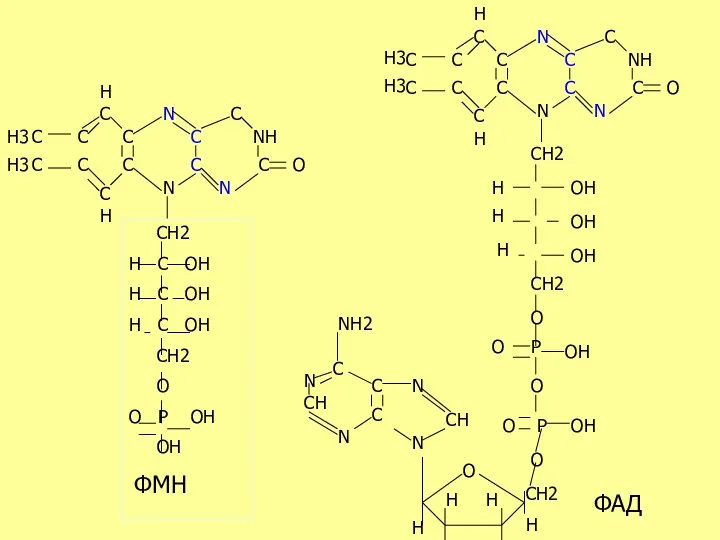
АЭРОБНЫЕ ДЕГИДРОГЕНАЗЫ
Эти ферменты катализируют реакции переноса атомов водорода (протоны и электроны)
АЭРОБНЫЕ ДЕГИДРОГЕНАЗЫ
Эти ферменты катализируют реакции переноса атомов водорода (протоны и электроны)
С
С
С
С
С
С
С
С
С
С
С
С
H
H
H3
H3
N
N
O
NH
N
CH2
H C OH
H C OH
H C OH
CH2
O
O P
С
С
С
С
С
С
С
С
С
С
С
С
H
H
H3
H3
N
N
O
NH
N
CH2
H C OH
H C OH
H C OH
CH2
O
O P
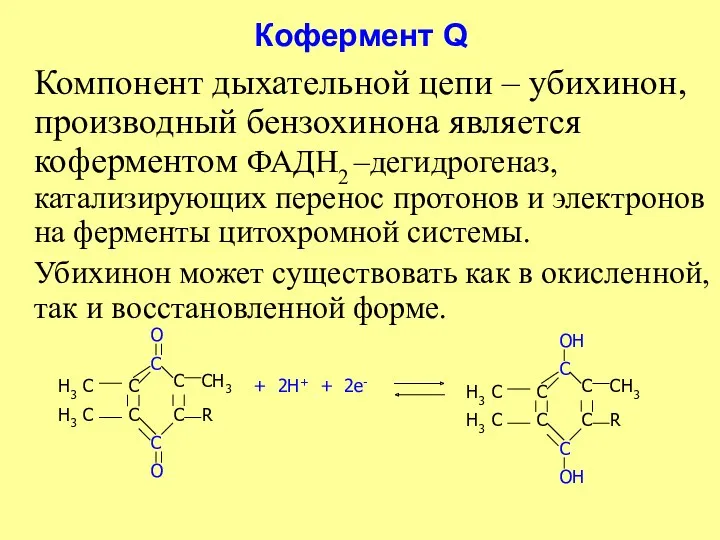
Кофермент Q
Компонент дыхательной цепи – убихинон, производный бензохинона является коферментом
Кофермент Q
Компонент дыхательной цепи – убихинон, производный бензохинона является коферментом

ЦИТОХРОМЫ
Дальнейший перенос электронов от убихинона на кислород осуществляет система цитохромов:
b,
ЦИТОХРОМЫ
Дальнейший перенос электронов от убихинона на кислород осуществляет система цитохромов:
b,

Механизм действия цитохромов в окислительно-восстановительных реакциях
Fe3+ + e- Fe2+ Fe2+ -
Механизм действия цитохромов в окислительно-восстановительных реакциях
Fe3+ + e- Fe2+ Fe2+ -

СТРУКТУРА ДЫХАТЕЛЬНОЙ ЦЕПИ
Перенос электронов и протонов в митохондриальной дыхательной цепи происходит
СТРУКТУРА ДЫХАТЕЛЬНОЙ ЦЕПИ
Перенос электронов и протонов в митохондриальной дыхательной цепи происходит
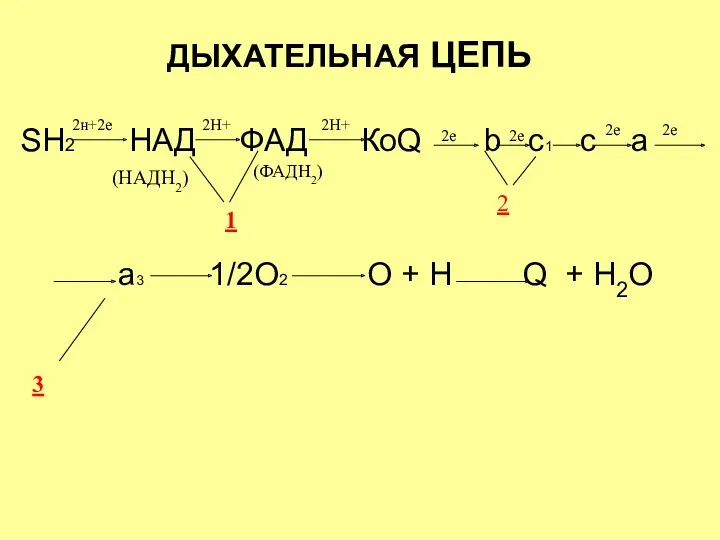
ДЫХАТЕЛЬНАЯ ЦЕПЬ
SH2 НАД ФАД КоQ b c1 c a
a3 1/2O2
ДЫХАТЕЛЬНАЯ ЦЕПЬ
SH2 НАД ФАД КоQ b c1 c a
a3 1/2O2
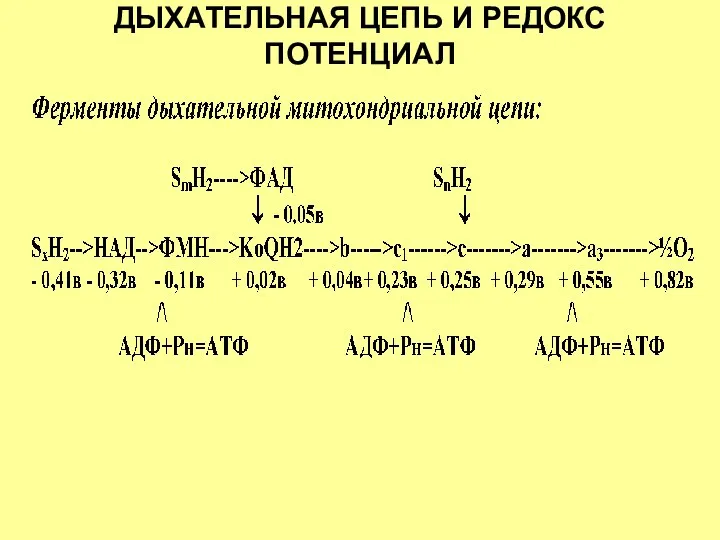
ДЫХАТЕЛЬНАЯ ЦЕПЬ И РЕДОКС ПОТЕНЦИАЛ
ДЫХАТЕЛЬНАЯ ЦЕПЬ И РЕДОКС ПОТЕНЦИАЛ

ПУНКТЫ ФОСФОРИЛИРОВАНИЯ
Первый пункт фосфорилирования находится между ферментами НАД и ФАД, где
ПУНКТЫ ФОСФОРИЛИРОВАНИЯ
Первый пункт фосфорилирования находится между ферментами НАД и ФАД, где
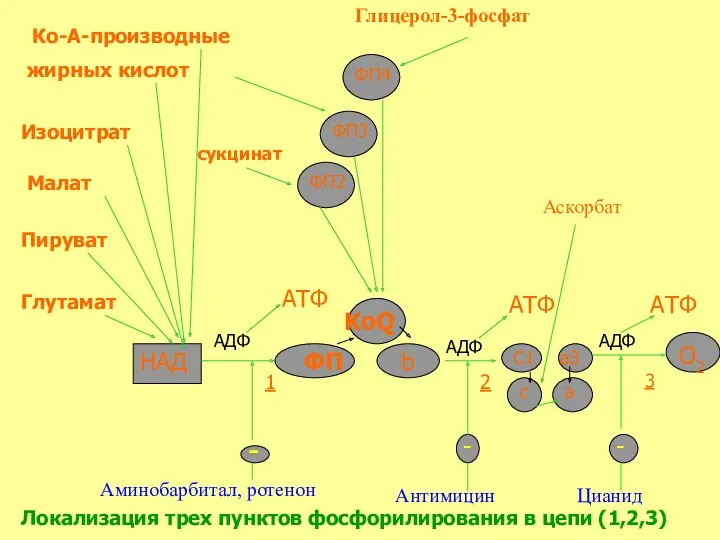
Ко-А-производные
жирных кислот
Изоцитрат
Малат
Пируват
Глутамат
НАД
ФП
b
КоQ
АТФ
АДФ
-
Аминобарбитал, ротенон
ФП2
ФП3
ФП4
Глицерол-3-фосфат
сукцинат
АДФ
АТФ
C1
c
a3
a
Антимицин
-
Аскорбат
О2
-
Цианид
АДФ
АТФ
1
2
3
Локализация трех пунктов фосфорилирования в цепи (1,2,3)
Ко-А-производные
жирных кислот
Изоцитрат
Малат
Пируват
Глутамат
НАД
ФП
b
КоQ
АТФ
АДФ
-
Аминобарбитал, ротенон
ФП2
ФП3
ФП4
Глицерол-3-фосфат
сукцинат
АДФ
АТФ
C1
c
a3
a
Антимицин
-
Аскорбат
О2
-
Цианид
АДФ
АТФ
1
2
3
Локализация трех пунктов фосфорилирования в цепи (1,2,3)

Окислительное фосфорилирование
При окислении одной молекулы субстрата на один атом поглощенного митохондриями
Окислительное фосфорилирование
При окислении одной молекулы субстрата на один атом поглощенного митохондриями

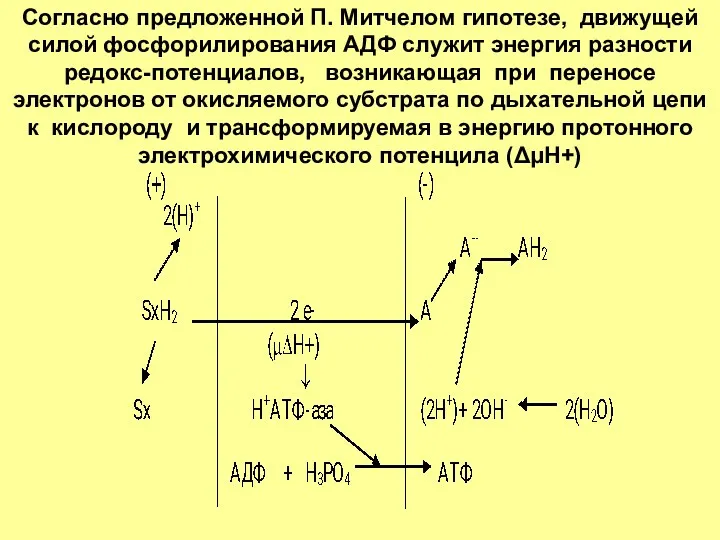
Согласно предложенной П. Митчелом гипотезе, движущей силой фосфорилирования АДФ служит энергия
Согласно предложенной П. Митчелом гипотезе, движущей силой фосфорилирования АДФ служит энергия
 Биологическая эффективность гербицида Miuris 125 в борьбе с однодольными сорными растениями в посевах сой
Биологическая эффективность гербицида Miuris 125 в борьбе с однодольными сорными растениями в посевах сой Биологическая память. Лекция № 31
Биологическая память. Лекция № 31 Способы размножения животных. Оплодотворение
Способы размножения животных. Оплодотворение Аптека под ногами
Аптека под ногами Высшая нервная деятельность. Лекция № 28
Высшая нервная деятельность. Лекция № 28 Травы и цветы полей и лугов
Травы и цветы полей и лугов Развитие эмбриологии в XVI—XVIII и начале XIX века. (Лекция 10)
Развитие эмбриологии в XVI—XVIII и начале XIX века. (Лекция 10) Мейоз. Деление мейоза
Мейоз. Деление мейоза Как размножаются живые организмы
Как размножаются живые организмы Неклеточные формы жизни. Вирусы и бактериофаги.
Неклеточные формы жизни. Вирусы и бактериофаги. Методы биологии
Методы биологии Бактерии
Бактерии Постэмбриональный период развития человека
Постэмбриональный период развития человека Кровь, кроветворение
Кровь, кроветворение Хим.состав клетки
Хим.состав клетки Пищевые цепи. Взаимосвязи организмов в природных сообществах
Пищевые цепи. Взаимосвязи организмов в природных сообществах Ферменты. АЛТ и АСТ
Ферменты. АЛТ и АСТ витамины
витамины Способность к размножению у животных
Способность к размножению у животных Дополнительный_материал_11.40_05.02.2024_eb5cb3bf (1)
Дополнительный_материал_11.40_05.02.2024_eb5cb3bf (1) Органеллы эукариотической клетки
Органеллы эукариотической клетки Экология и фитодизайн школьного кабинета биологии
Экология и фитодизайн школьного кабинета биологии Барханная кошка
Барханная кошка Клас ссавці
Клас ссавці Строение пластид. Фотосинтез
Строение пластид. Фотосинтез Адромискус. Семейство – толстянковые
Адромискус. Семейство – толстянковые Мутация. Геномдық мутация. Хромосомалық мутация. Гендік мутация. Мутацияның типтері
Мутация. Геномдық мутация. Хромосомалық мутация. Гендік мутация. Мутацияның типтері Что же такое трава?
Что же такое трава?