Электрическая и межклеточная передача возбуждения по нервному волокну и через нервномышечный синапс презентация
- Электрическая и межклеточная передача возбуждения по нервному волокну и через нервномышечный синапс

Содержание
- 2. СИНАПС Переход (передача) возбуждения с нервного волокна на иннервируемую им клетку (нервную, мышечную, секреторную) осуществляется через
- 3. 3. Синапсы могут быть между двумя нейронами (межнейронные), между нейроном и мышечным волокном (нервно-мышечные), между рецепторными
- 4. ЭЛЕКТРИЧЕСКИЙ СИНАПС Межклеточное образование, которое обеспечивает передачу импульса возбуждения посредством возникновения электрического тока между пресинаптическим и
- 6. СВОЙСТВА ЭЛЕКТРИЧЕСКИХ СИНАПСОВ Быстродействие (значительно превосходит в химических синапсах) Слабость следовых эффектов (практически отсутствует суммация последовательных
- 7. Электрические синапсы обладают рядом специфических функциональных свойств: синаптическая задержка практически отсутствует, т.е. интервал между приходом импульса
- 8. Ионные токи, перемещающиеся из пресинаптического нейрона в постсинаптический, вызывают на его мембране колебания разности потенциалов и
- 9. ХИМИЧЕСКИЙ СИНАПС Межклеточное образование, которое обеспечивает передачу сигнала с помощью химического посредника-медиатора Для химических синапсов общими
- 11. НЕЙРОМЕДИАТОРЫ Нейромедиаторы - низкомолекулярные вещества - поступают из синаптических пузырьков в синаптическую щель и связываются со
- 12. ПРЕСИНАПТИЧЕСКАЯ ЧАСТЬ Пресинаптическая часть содержит синаптические пузырьки с нейромедиатором, элементы цитоскелета и митохондрии. В пресинаптическую мембрану
- 13. Роль Са2+ Слияние синаптических пузырьков с пресинаптической мембраной происходит, когда увеличивается концентрации Са2+ в цитозоле нервной
- 14. АКТИВНЫЕ ЗОНЫ Секреция нейромедиатора осуществляется в специализированных участках пресинаптического нервного окончания - в активных зонах -
- 16. СИНАПТИЧЕСКАЯ ЩЕЛЬ Промежуток между пре- и постсинаптическими мембранами шириной 20-35 нм. В синаптическую щель из синаптических
- 17. ПОСТСИНАПТИЧЕСКАЯ ЧАСТЬ В постсинаптической мембране находятся рецепторы, чувствительные к нейромедиатору. Взаимодействие нейромедиатора с рецептором приводит к
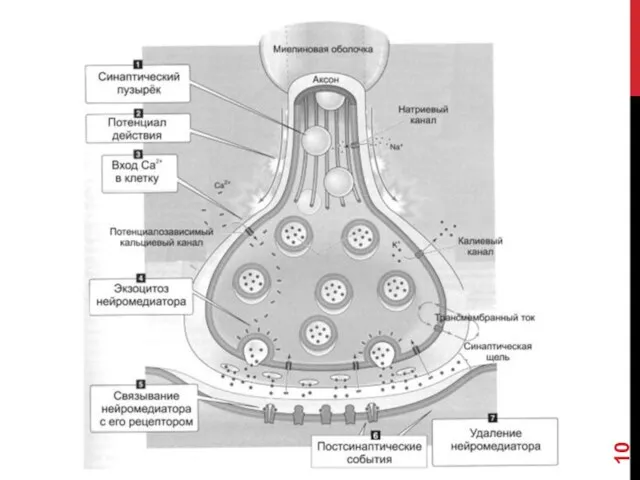
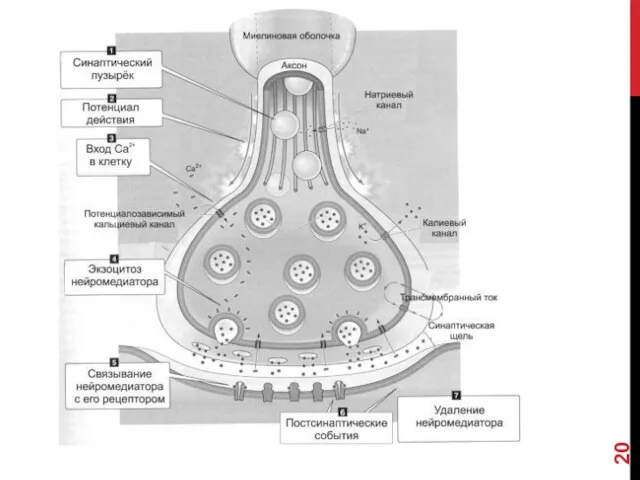
- 18. ЭТАПЫ СИНАПТИЧЕСКОЙ ПЕРЕДАЧИ 1. Молекулы нейромедиатора поступают в мембранные синаптические пузырьки, располагающиеся в пресинаптической терминали и
- 19. 5. Кванты нейромедиатора, поступившие в синаптическую щель, диффундируют в ней. Часть молекул нейромедиатора связывается со специфичными
- 21. НЕРВНО-МЫШЕЧНЫЙ СИНАПС Мионевральный (нервно-мышечный) синапс – образован аксоном мотонейрона и мышечной клеткой Структуры: пресинаптическая мембрана аксонной
- 22. НЕРВНО-МЫШЕЧНЫЙ СИНАПС Образован окончанием аксона моторного нейрона и мышечным волокном поперечно-полосатой мускулатуры Структуры: пресинаптическая мембрана аксонной
- 23. СТАДИИ ПЕРЕДАЧИ ВОЗБУЖДЕНИЯ ЧЕРЕЗ НЕРВНО-МЫШЕЧНЫЙ СИНАПС I. Трансформация электрического сигнала в химический: Потенциал действия (ПД) передается
- 24. 1 — миэлиновая оболочка аксона; 2 — концевые веточки аксона; 3 — пузырьки, содержащие ацетилхолин; 4
- 25. СТАДИИ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ ЧЕРЕЗ НЕРВНО-МЫШЕЧНЫЙ СИНАПС
- 26. СТРУКТУРА СИНАПСА
- 27. СРАВНИТЕЛЬНАЯ ХАРАКТЕРИСТИКА ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ ПО НЕРВНОМУ ВОЛОКНУ И ЧЕРЕЗ НЕРВНО-МЫШЕЧНЫЙ СИНАПС
- 28. В ЗАВИСИМОСТИ ОТ ХАРАКТЕРА ВОЗНИКАЮЩЕГО ПОСТСИНАПТИЧЕСКОГО ПОТЕНЦИАЛА (ДЕПОЛЯРИЗАЦИЯ ИЛИ ГИПЕРПОЛЯРИЗАЦИЯ) РАЗЛИЧАЮТ СИНАПСЫ ВОЗБУЖДАЮЩИЕ И ТОРМОЗНЫЕ Возбуждающий
- 29. Трансмембранный перенос ионов указан стрелками. При связывании ацетилхолина с никотиновым холинорецептором (н-холинорецептор) в составе последнего открывается
- 30. Характер электрического ответа постсинаптической стороны и дальнейший физиологический эффект определяются свойствами рецепторов. С точки зрения механизма
- 31. Ионотропные рецепторы сами по себе являются ионными каналами. Классический пример - н-холинорецепторы Метаботропные рецепторы связаны с
- 33. ВЫЗВАННЫЕ И СПОНТАННЫЕ ПОСТСИНАПТИЧЕСКИЕ ПОТЕНЦИАЛЫ (ПСП) Нейромедиаторы при связывании с ионотропными рецепторами вызывают ПСП. Количество квантов
- 34. СИНАПТИЧЕСКАЯ ЗАДЕРЖКА Между моментом поступления ПД к пресинаптическому нервному окончанию и временем возникновения ПСП существует временной
- 35. ВОЗБУЖДАЮЩИЕ И ТОРМОЗНЫЕ ПОСТСИНАПТИЧЕСКИЕ ПОТЕНЦИАЛЫ Возбуждающие ПСП вызваны возрастанием проводимости мембраны для Na+. Они деполяризуют постсинаптическую
- 36. СИНАПСЫ В НЕЙРОННЫХ СЕТЯХ функции нервной системы выполняются только при условии взаимодействия множества нервных клеток: нейронных
- 37. СИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ Торможение является одним из фундаментальных свойств ЦНС и было открыто в 1863 г. И.М.
- 38. Постсинаптическое торможение (рис. 6-7, 1В) наблюдается при выделении медиатора (например, ГАМК), повышающего проводимость постсинаптической мембраны для
- 39. Пресинаптическое торможение осуществляют нейроны, аксоны которых оканчиваются (Б) на возбуждающих синаптических окончаниях другого нейрона (А), образуя
- 40. Пресинаптическое облегчение Противоположный пресинаптическому торможению эффект оказывает пресинаптическое облегчение, обеспечивающее более продолжительное открытие Ca2+-каналов. Поскольку серотонин,
- 41. Возвратное торможение (рис. 6-7, 2). Нейроны ЦНС могут тормозить сами себя путём отрицательной обратной связи. Так,
- 42. Синаптическое торможение. 1 (слева) - пресинаптическое и постсинаптическое торможение: А - возбуждающее окончание, Б - нейрон,
- 43. СУММАЦИЯ, ОККЛЮЗИЯ И УТОМЛЕНИЕ Суммация. В мозге дендритная зона одного нейрона формирует с другими нервными клетками
- 44. Окклюзия. В некоторых случаях раздельная активация нейронов более эффективна, чем одновременная. Этот феномен, называемый окклюзией, рассмотрен
- 45. Суммация и окклюзия в нейронных сетях
- 46. Утомление. Повторная стимуляция возбуждающих синапсов с высокой частотой вначале вызывает появление большого количества разрядов в постсинаптических
- 47. ПЛАСТИЧНОСТЬ СИНАПСОВ В ходе функционирования синапсы подвергаются функциональным и морфологическим перестройкам. Этот процесс назван синаптической пластичностью.
- 48. Формы синаптической пластичности
- 49. КРАТКОВРЕМЕННЫЕ ФОРМЫ СИНАПТИЧЕСКОЙ ПЛАСТИЧНОСТИ (ОБЛЕГЧЕНИЕ, ПОТЕНЦИАЦИЯ, ДЕПРЕССИЯ И ПРИВЫКАНИЕ) Облегчение. В процессе активности в синапсах с
- 50. Потенциация, посттетаническая потенциация (сенситизация). Увеличение ПСП при высокочастотной активности может иметь и постсинаптическую природу. Такой вид
- 51. ДОЛГОВРЕМЕННЫЕ ФОРМЫ СИНАПТИЧЕСКОЙ ПЛАСТИЧНОСТИ Долговременная потенциация - быстро развивающееся устойчивое усиление синаптической передачи в ответ на
- 53. Скачать презентацию

СИНАПС
Переход (передача) возбуждения с нервного волокна на иннервируемую им клетку (нервную,
СИНАПС
Переход (передача) возбуждения с нервного волокна на иннервируемую им клетку (нервную,

3. Синапсы могут быть между двумя нейронами (межнейронные), между нейроном и
3. Синапсы могут быть между двумя нейронами (межнейронные), между нейроном и

ЭЛЕКТРИЧЕСКИЙ СИНАПС
Межклеточное образование, которое обеспечивает передачу импульса возбуждения посредством возникновения электрического
ЭЛЕКТРИЧЕСКИЙ СИНАПС
Межклеточное образование, которое обеспечивает передачу импульса возбуждения посредством возникновения электрического
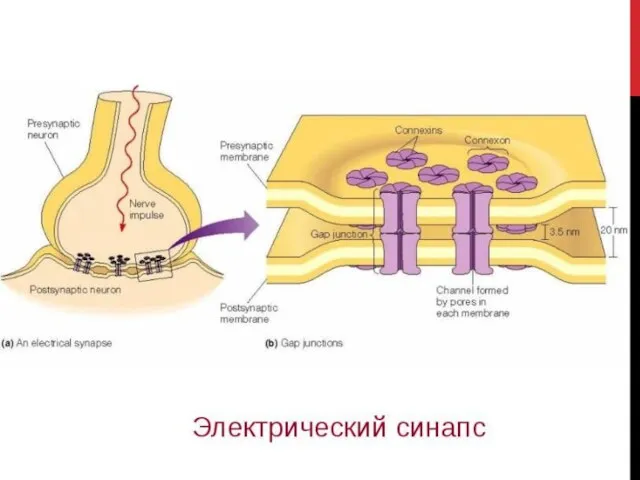

СВОЙСТВА ЭЛЕКТРИЧЕСКИХ СИНАПСОВ
Быстродействие (значительно превосходит в химических синапсах)
Слабость следовых эффектов (практически
СВОЙСТВА ЭЛЕКТРИЧЕСКИХ СИНАПСОВ
Быстродействие (значительно превосходит в химических синапсах)
Слабость следовых эффектов (практически

Электрические синапсы обладают рядом специфических функциональных свойств:
синаптическая задержка практически отсутствует, т.е.
Электрические синапсы обладают рядом специфических функциональных свойств:
синаптическая задержка практически отсутствует, т.е.

Ионные токи, перемещающиеся из пресинаптического нейрона в постсинаптический, вызывают на его
Ионные токи, перемещающиеся из пресинаптического нейрона в постсинаптический, вызывают на его

ХИМИЧЕСКИЙ СИНАПС
Межклеточное образование, которое обеспечивает передачу сигнала с помощью химического посредника-медиатора
Для
ХИМИЧЕСКИЙ СИНАПС
Межклеточное образование, которое обеспечивает передачу сигнала с помощью химического посредника-медиатора
Для

НЕЙРОМЕДИАТОРЫ
Нейромедиаторы - низкомолекулярные вещества - поступают из синаптических пузырьков в синаптическую
НЕЙРОМЕДИАТОРЫ
Нейромедиаторы - низкомолекулярные вещества - поступают из синаптических пузырьков в синаптическую

ПРЕСИНАПТИЧЕСКАЯ ЧАСТЬ
Пресинаптическая часть содержит синаптические пузырьки с нейромедиатором, элементы цитоскелета и
ПРЕСИНАПТИЧЕСКАЯ ЧАСТЬ
Пресинаптическая часть содержит синаптические пузырьки с нейромедиатором, элементы цитоскелета и

Роль Са2+
Слияние синаптических пузырьков с пресинаптической мембраной происходит, когда увеличивается концентрации
Роль Са2+
Слияние синаптических пузырьков с пресинаптической мембраной происходит, когда увеличивается концентрации

АКТИВНЫЕ ЗОНЫ
Секреция нейромедиатора осуществляется в специализированных участках пресинаптического нервного окончания -
АКТИВНЫЕ ЗОНЫ
Секреция нейромедиатора осуществляется в специализированных участках пресинаптического нервного окончания -
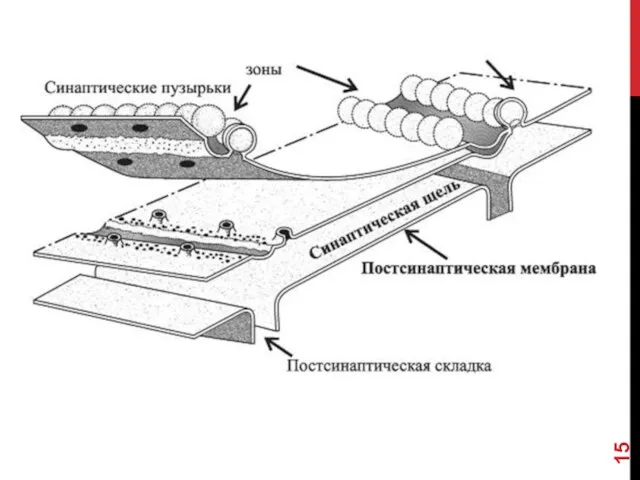

СИНАПТИЧЕСКАЯ ЩЕЛЬ
Промежуток между пре- и постсинаптическими мембранами шириной 20-35 нм. В
СИНАПТИЧЕСКАЯ ЩЕЛЬ
Промежуток между пре- и постсинаптическими мембранами шириной 20-35 нм. В

ПОСТСИНАПТИЧЕСКАЯ ЧАСТЬ
В постсинаптической мембране находятся рецепторы, чувствительные к нейромедиатору. Взаимодействие нейромедиатора
ПОСТСИНАПТИЧЕСКАЯ ЧАСТЬ
В постсинаптической мембране находятся рецепторы, чувствительные к нейромедиатору. Взаимодействие нейромедиатора

ЭТАПЫ СИНАПТИЧЕСКОЙ ПЕРЕДАЧИ
1. Молекулы нейромедиатора поступают в мембранные синаптические пузырьки, располагающиеся
ЭТАПЫ СИНАПТИЧЕСКОЙ ПЕРЕДАЧИ
1. Молекулы нейромедиатора поступают в мембранные синаптические пузырьки, располагающиеся

5. Кванты нейромедиатора, поступившие в синаптическую щель, диффундируют в ней. Часть
5. Кванты нейромедиатора, поступившие в синаптическую щель, диффундируют в ней. Часть

НЕРВНО-МЫШЕЧНЫЙ СИНАПС
Мионевральный (нервно-мышечный) синапс – образован аксоном мотонейрона и мышечной клеткой
Структуры:
НЕРВНО-МЫШЕЧНЫЙ СИНАПС
Мионевральный (нервно-мышечный) синапс – образован аксоном мотонейрона и мышечной клеткой
Структуры:

НЕРВНО-МЫШЕЧНЫЙ СИНАПС
Образован окончанием аксона моторного нейрона и мышечным волокном поперечно-полосатой мускулатуры
Структуры:
НЕРВНО-МЫШЕЧНЫЙ СИНАПС
Образован окончанием аксона моторного нейрона и мышечным волокном поперечно-полосатой мускулатуры
Структуры:

СТАДИИ ПЕРЕДАЧИ ВОЗБУЖДЕНИЯ ЧЕРЕЗ НЕРВНО-МЫШЕЧНЫЙ СИНАПС
I. Трансформация электрического сигнала в химический:
Потенциал
СТАДИИ ПЕРЕДАЧИ ВОЗБУЖДЕНИЯ ЧЕРЕЗ НЕРВНО-МЫШЕЧНЫЙ СИНАПС
I. Трансформация электрического сигнала в химический:
Потенциал

1 — миэлиновая оболочка аксона;
2 — концевые веточки аксона;
3 — пузырьки,
1 — миэлиновая оболочка аксона;
2 — концевые веточки аксона;
3 — пузырьки,
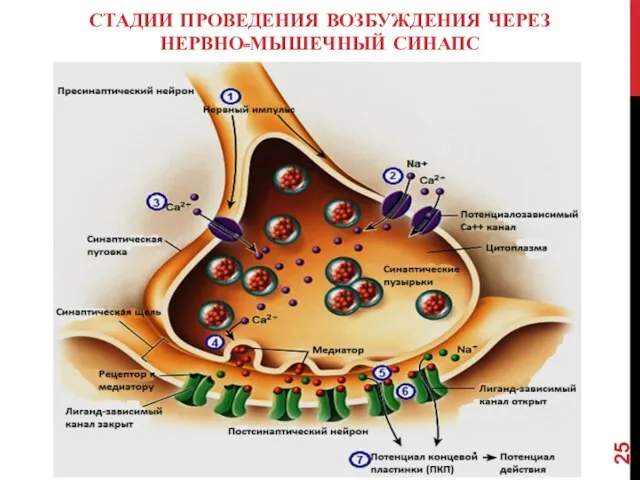
СТАДИИ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ ЧЕРЕЗ НЕРВНО-МЫШЕЧНЫЙ СИНАПС
СТАДИИ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ ЧЕРЕЗ НЕРВНО-МЫШЕЧНЫЙ СИНАПС
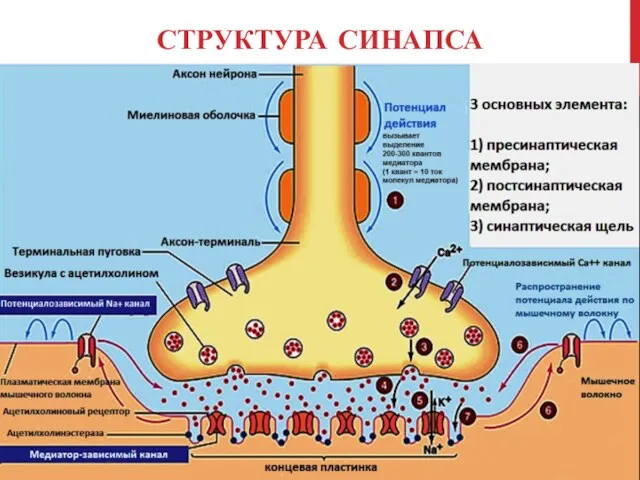
СТРУКТУРА СИНАПСА
СТРУКТУРА СИНАПСА
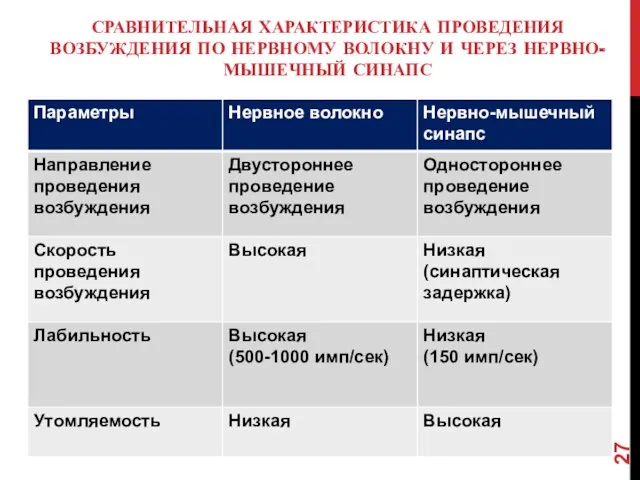
СРАВНИТЕЛЬНАЯ ХАРАКТЕРИСТИКА ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ ПО НЕРВНОМУ ВОЛОКНУ И ЧЕРЕЗ НЕРВНО-МЫШЕЧНЫЙ СИНАПС
СРАВНИТЕЛЬНАЯ ХАРАКТЕРИСТИКА ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ ПО НЕРВНОМУ ВОЛОКНУ И ЧЕРЕЗ НЕРВНО-МЫШЕЧНЫЙ СИНАПС

В ЗАВИСИМОСТИ ОТ ХАРАКТЕРА ВОЗНИКАЮЩЕГО ПОСТСИНАПТИЧЕСКОГО ПОТЕНЦИАЛА (ДЕПОЛЯРИЗАЦИЯ ИЛИ ГИПЕРПОЛЯРИЗАЦИЯ)
В ЗАВИСИМОСТИ ОТ ХАРАКТЕРА ВОЗНИКАЮЩЕГО ПОСТСИНАПТИЧЕСКОГО ПОТЕНЦИАЛА (ДЕПОЛЯРИЗАЦИЯ ИЛИ ГИПЕРПОЛЯРИЗАЦИЯ)
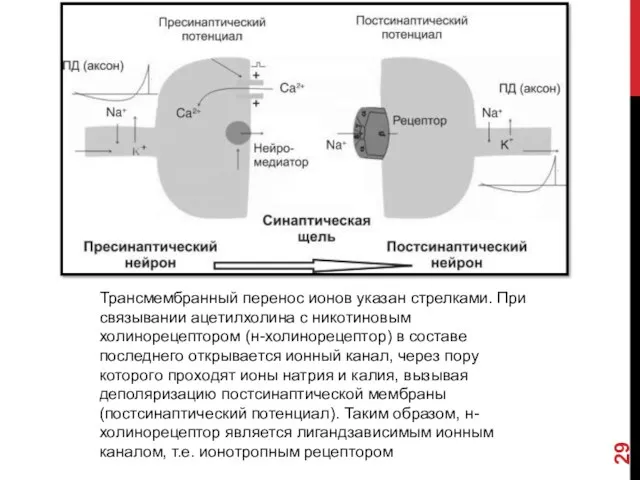
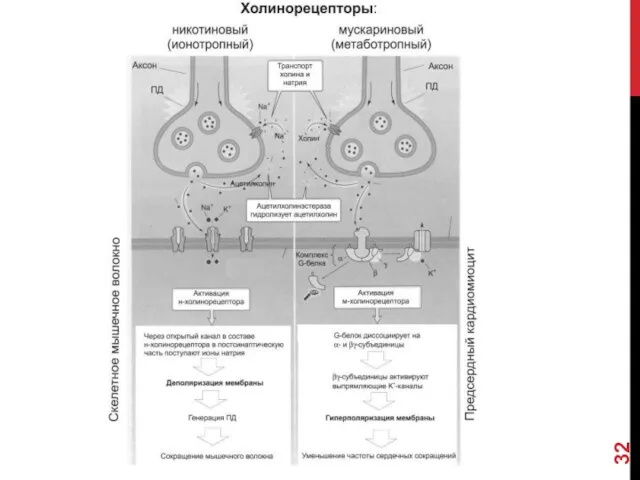
Трансмембранный перенос ионов указан стрелками. При связывании ацетилхолина с никотиновым холинорецептором
Трансмембранный перенос ионов указан стрелками. При связывании ацетилхолина с никотиновым холинорецептором

Характер электрического ответа постсинаптической стороны и дальнейший физиологический эффект определяются свойствами
Характер электрического ответа постсинаптической стороны и дальнейший физиологический эффект определяются свойствами

Ионотропные рецепторы сами по себе являются ионными каналами. Классический пример
Ионотропные рецепторы сами по себе являются ионными каналами. Классический пример

ВЫЗВАННЫЕ И СПОНТАННЫЕ ПОСТСИНАПТИЧЕСКИЕ ПОТЕНЦИАЛЫ (ПСП)
Нейромедиаторы при связывании с ионотропными рецепторами
ВЫЗВАННЫЕ И СПОНТАННЫЕ ПОСТСИНАПТИЧЕСКИЕ ПОТЕНЦИАЛЫ (ПСП)
Нейромедиаторы при связывании с ионотропными рецепторами

СИНАПТИЧЕСКАЯ ЗАДЕРЖКА
Между моментом поступления ПД к пресинаптическому нервному окончанию и временем
СИНАПТИЧЕСКАЯ ЗАДЕРЖКА
Между моментом поступления ПД к пресинаптическому нервному окончанию и временем

ВОЗБУЖДАЮЩИЕ И ТОРМОЗНЫЕ ПОСТСИНАПТИЧЕСКИЕ ПОТЕНЦИАЛЫ
Возбуждающие ПСП вызваны возрастанием проводимости мембраны
ВОЗБУЖДАЮЩИЕ И ТОРМОЗНЫЕ ПОСТСИНАПТИЧЕСКИЕ ПОТЕНЦИАЛЫ
Возбуждающие ПСП вызваны возрастанием проводимости мембраны

СИНАПСЫ В НЕЙРОННЫХ СЕТЯХ
функции нервной системы выполняются только при условии взаимодействия
СИНАПСЫ В НЕЙРОННЫХ СЕТЯХ
функции нервной системы выполняются только при условии взаимодействия

СИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ
Торможение является одним из фундаментальных свойств ЦНС и было открыто
СИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ
Торможение является одним из фундаментальных свойств ЦНС и было открыто

Постсинаптическое торможение (рис. 6-7, 1В) наблюдается при выделении медиатора (например,
Постсинаптическое торможение (рис. 6-7, 1В) наблюдается при выделении медиатора (например,

Пресинаптическое торможение осуществляют нейроны, аксоны которых оканчиваются (Б) на возбуждающих синаптических
Пресинаптическое торможение осуществляют нейроны, аксоны которых оканчиваются (Б) на возбуждающих синаптических

Пресинаптическое облегчение
Противоположный пресинаптическому торможению эффект оказывает пресинаптическое облегчение, обеспечивающее более
Пресинаптическое облегчение
Противоположный пресинаптическому торможению эффект оказывает пресинаптическое облегчение, обеспечивающее более

Возвратное торможение (рис. 6-7, 2). Нейроны ЦНС могут тормозить сами себя
Возвратное торможение (рис. 6-7, 2). Нейроны ЦНС могут тормозить сами себя
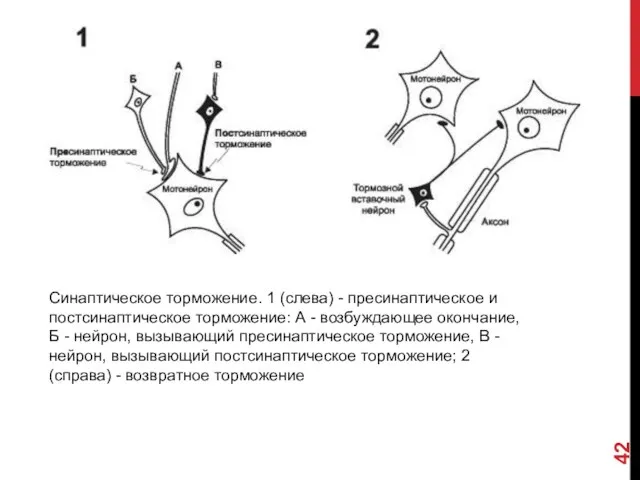
Синаптическое торможение. 1 (слева) - пресинаптическое и постсинаптическое торможение: А -
Синаптическое торможение. 1 (слева) - пресинаптическое и постсинаптическое торможение: А -

СУММАЦИЯ, ОККЛЮЗИЯ И УТОМЛЕНИЕ
Суммация. В мозге дендритная зона одного нейрона формирует
СУММАЦИЯ, ОККЛЮЗИЯ И УТОМЛЕНИЕ
Суммация. В мозге дендритная зона одного нейрона формирует

Окклюзия. В некоторых случаях раздельная активация нейронов более эффективна, чем одновременная.
Окклюзия. В некоторых случаях раздельная активация нейронов более эффективна, чем одновременная.
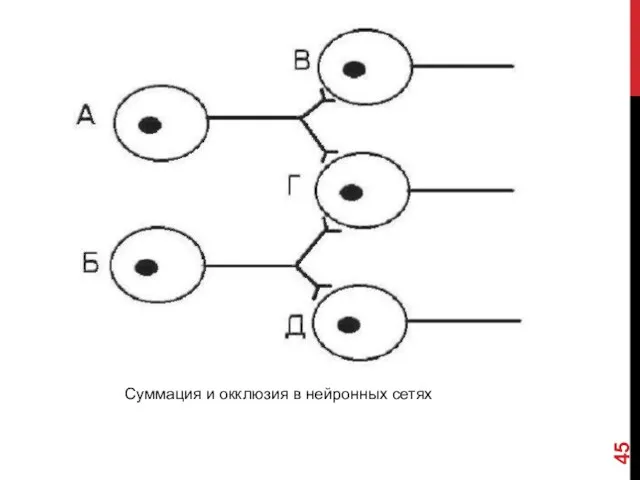
Суммация и окклюзия в нейронных сетях
Суммация и окклюзия в нейронных сетях

Утомление. Повторная стимуляция возбуждающих синапсов с высокой частотой вначале вызывает
Утомление. Повторная стимуляция возбуждающих синапсов с высокой частотой вначале вызывает

ПЛАСТИЧНОСТЬ СИНАПСОВ
В ходе функционирования синапсы подвергаются функциональным и морфологическим перестройкам. Этот
ПЛАСТИЧНОСТЬ СИНАПСОВ
В ходе функционирования синапсы подвергаются функциональным и морфологическим перестройкам. Этот
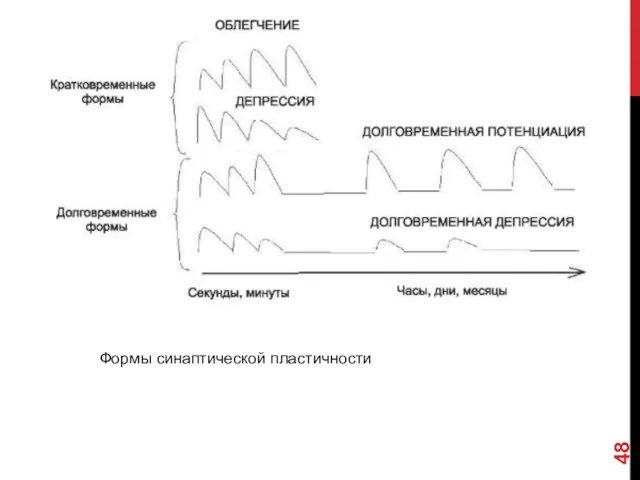
Формы синаптической пластичности
Формы синаптической пластичности

КРАТКОВРЕМЕННЫЕ ФОРМЫ СИНАПТИЧЕСКОЙ ПЛАСТИЧНОСТИ (ОБЛЕГЧЕНИЕ, ПОТЕНЦИАЦИЯ, ДЕПРЕССИЯ И ПРИВЫКАНИЕ)
Облегчение. В
КРАТКОВРЕМЕННЫЕ ФОРМЫ СИНАПТИЧЕСКОЙ ПЛАСТИЧНОСТИ (ОБЛЕГЧЕНИЕ, ПОТЕНЦИАЦИЯ, ДЕПРЕССИЯ И ПРИВЫКАНИЕ)
Облегчение. В

Потенциация, посттетаническая потенциация (сенситизация). Увеличение ПСП при высокочастотной активности может иметь
Потенциация, посттетаническая потенциация (сенситизация). Увеличение ПСП при высокочастотной активности может иметь

ДОЛГОВРЕМЕННЫЕ ФОРМЫ СИНАПТИЧЕСКОЙ ПЛАСТИЧНОСТИ
Долговременная потенциация - быстро развивающееся устойчивое усиление синаптической
ДОЛГОВРЕМЕННЫЕ ФОРМЫ СИНАПТИЧЕСКОЙ ПЛАСТИЧНОСТИ
Долговременная потенциация - быстро развивающееся устойчивое усиление синаптической
 Строение микроскопа
Строение микроскопа Строение клетки. Основные органоиды
Строение клетки. Основные органоиды Строение клетки
Строение клетки Полиморфизм человеческих популяций. Генетический груз. Популяционная структура человечества
Полиморфизм человеческих популяций. Генетический груз. Популяционная структура человечества Паразитология. Разделы паразитологии
Паразитология. Разделы паразитологии Воздушные и тепловые свойства почв
Воздушные и тепловые свойства почв Дополнительная презентация Что такое фитодизайн?к элективному курсу Основы фитодизайна, 9 класс
Дополнительная презентация Что такое фитодизайн?к элективному курсу Основы фитодизайна, 9 класс Значение водорослей в природе
Значение водорослей в природе Основы аграрных технологий. 7 класс
Основы аграрных технологий. 7 класс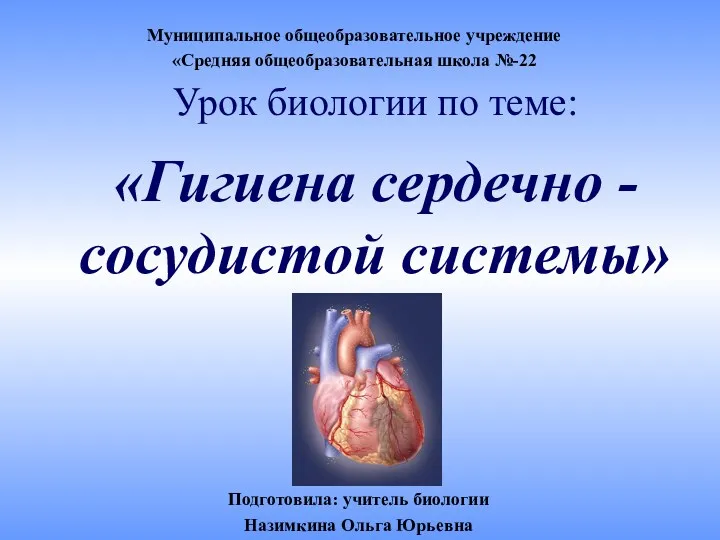 Урок биологии по теме: Гигиена сердечно - сосудистой системы
Урок биологии по теме: Гигиена сердечно - сосудистой системы Вода. Роль воды в жизни человека
Вода. Роль воды в жизни человека Насекомые. Особенности строения и жизнедеятельности
Насекомые. Особенности строения и жизнедеятельности Acid-base equilibrium in biological systems
Acid-base equilibrium in biological systems Проводящие пути. 1
Проводящие пути. 1 урок красная книга Мордовии
урок красная книга Мордовии Дисахариды. Полисахариды
Дисахариды. Полисахариды Животные и растения Красной книги Самарской области
Животные и растения Красной книги Самарской области Сучасні молекулярно-генетичні методи досліджень спадковості людини
Сучасні молекулярно-генетичні методи досліджень спадковості людини Fish are gill-bearing aquatic craniate animals that lack limbs with digits
Fish are gill-bearing aquatic craniate animals that lack limbs with digits Торғайтәрізділер (Passerіformes) – құстар отряды
Торғайтәрізділер (Passerіformes) – құстар отряды Технология выращивания комнатных растений
Технология выращивания комнатных растений Қамба бізтұмсығы және қамба күйесі
Қамба бізтұмсығы және қамба күйесі Пищевые отравления
Пищевые отравления Жалпақ құрттар
Жалпақ құрттар Приспособленность организмов как результат естественного отбора
Приспособленность организмов как результат естественного отбора Качество питания и здоровье -презентация к родительскому лекторию
Качество питания и здоровье -презентация к родительскому лекторию Презентация по биологии 5 класс Как питается растение?
Презентация по биологии 5 класс Как питается растение? Человек как результат биологической и социокультурной эволюции
Человек как результат биологической и социокультурной эволюции