- Элементарные факторы эволюции

Содержание
- 2. ЭВОЛЮЦИОННЫЕ ФАКТОРЫ - это любые явления или процессы, оказывающие какое-либо воздействие на эволюцию организмов (изменчивость организмов
- 3. МУТАЦИОННЫЙ ПРОЦЕСС МУТАЦИОННЫЙ ПРОЦЕСС – это процесс возникновения в популяциях самых разнообразных мутаций: генных, хромосомных и
- 4. Частота мутаций конкретных генов очень низка: она колеблется от 10–3 (мутирует один ген из тысячи) до
- 5. Судьба мутантного аллеля в популяциях диплобионтов Диплобионты – это диплоидные организмы, к которым относятся все животные,
- 6. 1. Рецессивные аутосомные мутации Практически не имеют шансов проявиться в фенотипе. Вероятность встречи мутантных аллелей равна
- 7. 2. Рецессивные мутации, сцепленные с полом. Такие мутации сразу проявляются у гетерогаметного пола. У человека такие
- 8. Судьба мутантного аллеля в популяциях гаплобионтов и полиплоидов К гаплобионтам относятся все прокариоты; водоросли и грибы
- 9. ДАВЛЕНИЕ МУТАЦИЙ Одна и та же мутация с одной и той же частотой р возникает в
- 10. МЕЙОТИЧЕСКИЙ ДРАЙВ механизмы, с помощью которых мутантные гены могут избегать элиминации естественным отбором. Если такой ген
- 11. РЕКОМБИНАЦИИ Как правило, один фенотипический признак формируется под влиянием множества генетических факторов, то есть является полигенным.
- 12. ДРЕЙФ ГЕНОВ Дрейф генов - случайные ненаправленные изменения частот аллелей в популяциях. В малых популяциях случайная
- 13. 1. ПОПУЛЯЦИОННЫЕ ВОЛНЫ Популяционные волны (волны жизни, волны численности) - колебания численности природных популяций. Типы популяционных
- 14. 2. ИЗОЛЯЦИЯ. В большой популяции частота мутации порядка 10–6 означает, что примерно одна из миллиона особей
- 15. ИЗОЛЯЦИЯ ФИЗИКО-ГЕОГРАФИЧЕСКАЯ Механическая, пространственная БИОЛОГИЧЕСКАЯ ЭКОЛОГИЧЕСКАЯ ЭТОЛОГИЧЕСКАЯ МОРФО-ФУНКЦИОНАЛЬНАЯ ГЕНЕТИЧЕСКАЯ СЕЗОННАЯ БИОТОПИЧЕСКАЯ - Особенности поведения у животных
- 16. ГЕОГРАФИЧЕСКАЯ ИЗОЛЯЦИЯ - отделение данной популяции от других популяций родительского вида каким-либо географическим барьером, непроходимым для
- 17. Изолированные популяции могут возникать двумя путями: при появлении каких-либо географических барьеров при активном расселении вида на
- 18. 3. ЭФФЕКТ ОСНОВАТЕЛЯ Эффект основателя представляет собой частный случай совместного действия изоляции и популяционных волн. Популяция
- 20. Скачать презентацию

ЭВОЛЮЦИОННЫЕ ФАКТОРЫ - это любые явления или процессы, оказывающие какое-либо воздействие
ЭВОЛЮЦИОННЫЕ ФАКТОРЫ - это любые явления или процессы, оказывающие какое-либо воздействие

МУТАЦИОННЫЙ ПРОЦЕСС
МУТАЦИОННЫЙ ПРОЦЕСС – это процесс возникновения в популяциях самых разнообразных
МУТАЦИОННЫЙ ПРОЦЕСС
МУТАЦИОННЫЙ ПРОЦЕСС – это процесс возникновения в популяциях самых разнообразных

Частота мутаций конкретных генов очень низка: она колеблется от 10–3 (мутирует

Судьба мутантного аллеля в популяциях диплобионтов
Диплобионты – это диплоидные организмы, к
Судьба мутантного аллеля в популяциях диплобионтов
Диплобионты – это диплоидные организмы, к

1. Рецессивные аутосомные мутации
Практически не имеют шансов проявиться в фенотипе.
1. Рецессивные аутосомные мутации
Практически не имеют шансов проявиться в фенотипе.

2. Рецессивные мутации, сцепленные с полом.
Такие мутации сразу проявляются у
2. Рецессивные мутации, сцепленные с полом.
Такие мутации сразу проявляются у

Судьба мутантного аллеля в популяциях гаплобионтов и полиплоидов
К гаплобионтам относятся все
Судьба мутантного аллеля в популяциях гаплобионтов и полиплоидов
К гаплобионтам относятся все
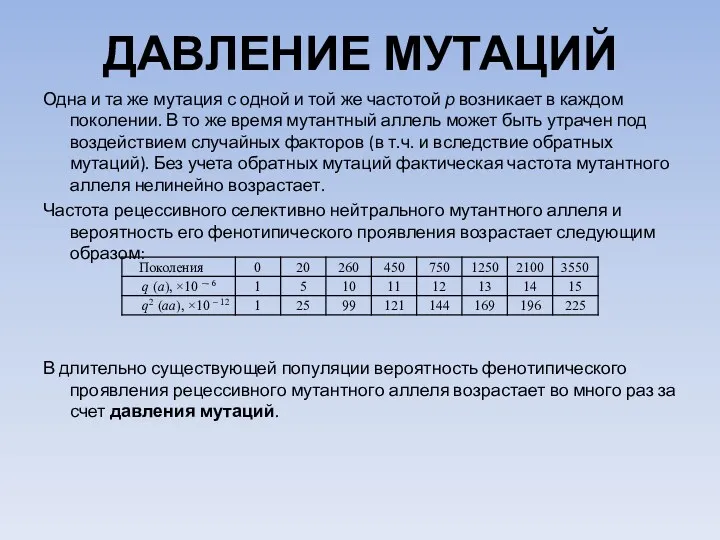
ДАВЛЕНИЕ МУТАЦИЙ
Одна и та же мутация с одной и той же
ДАВЛЕНИЕ МУТАЦИЙ
Одна и та же мутация с одной и той же

МЕЙОТИЧЕСКИЙ ДРАЙВ
механизмы, с помощью которых мутантные гены могут избегать элиминации естественным
МЕЙОТИЧЕСКИЙ ДРАЙВ
механизмы, с помощью которых мутантные гены могут избегать элиминации естественным

РЕКОМБИНАЦИИ
Как правило, один фенотипический признак формируется под влиянием множества генетических факторов,
РЕКОМБИНАЦИИ
Как правило, один фенотипический признак формируется под влиянием множества генетических факторов,

ДРЕЙФ ГЕНОВ
Дрейф генов - случайные ненаправленные изменения частот аллелей в популяциях.
ДРЕЙФ ГЕНОВ
Дрейф генов - случайные ненаправленные изменения частот аллелей в популяциях.

1. ПОПУЛЯЦИОННЫЕ ВОЛНЫ
Популяционные волны (волны жизни, волны численности) - колебания численности
1. ПОПУЛЯЦИОННЫЕ ВОЛНЫ
Популяционные волны (волны жизни, волны численности) - колебания численности

2. ИЗОЛЯЦИЯ.
В большой популяции частота мутации порядка 10–6 означает, что
2. ИЗОЛЯЦИЯ.
В большой популяции частота мутации порядка 10–6 означает, что
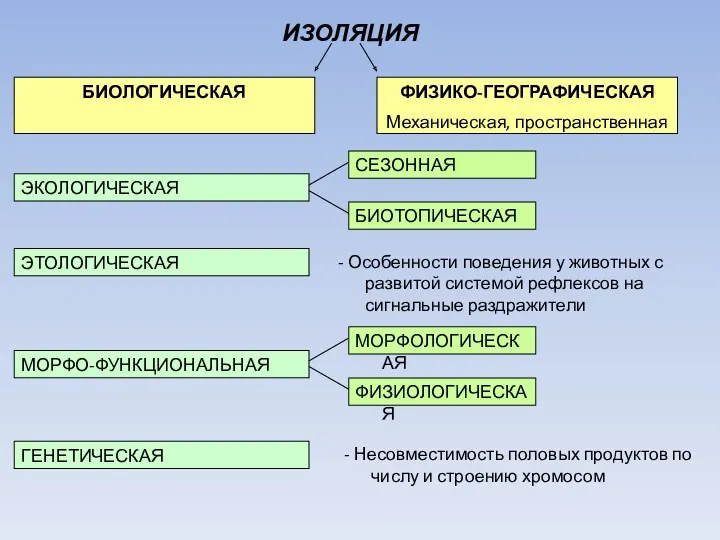
ИЗОЛЯЦИЯ
ФИЗИКО-ГЕОГРАФИЧЕСКАЯ
Механическая, пространственная
БИОЛОГИЧЕСКАЯ
ЭКОЛОГИЧЕСКАЯ
ЭТОЛОГИЧЕСКАЯ
МОРФО-ФУНКЦИОНАЛЬНАЯ
ГЕНЕТИЧЕСКАЯ
СЕЗОННАЯ
БИОТОПИЧЕСКАЯ
- Особенности поведения у животных с развитой системой рефлексов
ИЗОЛЯЦИЯ
ФИЗИКО-ГЕОГРАФИЧЕСКАЯ
Механическая, пространственная
БИОЛОГИЧЕСКАЯ
ЭКОЛОГИЧЕСКАЯ
ЭТОЛОГИЧЕСКАЯ
МОРФО-ФУНКЦИОНАЛЬНАЯ
ГЕНЕТИЧЕСКАЯ
СЕЗОННАЯ
БИОТОПИЧЕСКАЯ
- Особенности поведения у животных с развитой системой рефлексов

ГЕОГРАФИЧЕСКАЯ ИЗОЛЯЦИЯ - отделение данной популяции от других популяций родительского вида
ГЕОГРАФИЧЕСКАЯ ИЗОЛЯЦИЯ - отделение данной популяции от других популяций родительского вида

Изолированные популяции могут возникать двумя путями:
при появлении каких-либо географических барьеров
при
Изолированные популяции могут возникать двумя путями:
при появлении каких-либо географических барьеров
при

3. ЭФФЕКТ ОСНОВАТЕЛЯ
Эффект основателя представляет собой частный случай совместного действия изоляции
Эффект основателя представляет собой частный случай совместного действия изоляции
 Царство животные
Царство животные викторина по биологии
викторина по биологии Додарвиновский период. Развитие эволюционных идей в биологии
Додарвиновский период. Развитие эволюционных идей в биологии Факторы среды и их влияние на биоценозы. (7 класс)
Факторы среды и их влияние на биоценозы. (7 класс) Сучасна клітинна теорія. Клітина – елементарна цілісна жива система
Сучасна клітинна теорія. Клітина – елементарна цілісна жива система Нуклеопротеины и нуклеиновые кислоты
Нуклеопротеины и нуклеиновые кислоты Функциональная анатомия спинного и головного мозга
Функциональная анатомия спинного и головного мозга Эндокринная система. Физиология желез внутренней секреции
Эндокринная система. Физиология желез внутренней секреции Корень. Строение и функции
Корень. Строение и функции Уразовский Заказник
Уразовский Заказник Общие представления о биомолекулах. Химическая эволюция. Прокариоты и эукариоты. Структура клеток. Вирусы
Общие представления о биомолекулах. Химическая эволюция. Прокариоты и эукариоты. Структура клеток. Вирусы Транскрипция, трансляция
Транскрипция, трансляция Цитоплазма. Химический состав
Цитоплазма. Химический состав Зоопарк (зоологічний парк)
Зоопарк (зоологічний парк) Вода как фактор роста и развития растений
Вода как фактор роста и развития растений В гости к весне. Изменения в живой и неживой природе весной
В гости к весне. Изменения в живой и неживой природе весной Витамины - кофакторы ферментативных реакций. Классификация витаминов. Гиповитаминозы. Гипервитаминозы. Антивитамины
Витамины - кофакторы ферментативных реакций. Классификация витаминов. Гиповитаминозы. Гипервитаминозы. Антивитамины Сравнение процессов развития половых клеток у растений и животных
Сравнение процессов развития половых клеток у растений и животных Цикл развития кузнечика
Цикл развития кузнечика Ямчатость стебля
Ямчатость стебля Характеристика слухового анализатора
Характеристика слухового анализатора Понятие о сортах сельскохозяйственных культур
Понятие о сортах сельскохозяйственных культур Эволюционная теория
Эволюционная теория Методы познания живой природы
Методы познания живой природы Анатомия листа С3 и С4 растений. Особенности фиксация диоксида углерода в клетках мезофилла. Акцепторы диоксида углерода
Анатомия листа С3 и С4 растений. Особенности фиксация диоксида углерода в клетках мезофилла. Акцепторы диоксида углерода Водоросли. Строение водорослей
Водоросли. Строение водорослей The Nervous System
The Nervous System Многообразие видов в природе
Многообразие видов в природе