- Ямчатость стебля

Содержание
- 2. Общие положения Особенности строения древесины и луба карельской березы, как это видно из предыдущей лекции, дают
- 6. Причина возникновения ямчатости на поверхности древесинного цилиндра не во всех случаях выяснена. Когда возбудитель этого изменения
- 7. При этом наряду с ямчатостью в местах соединения компонентов прививки возникают наплывы древесины так называемая “зобоватость”
- 8. В результате изменений в лучевых инициалях происходит расширение лучей, их слияние и образование скопления недифференцированных изодиаметрических
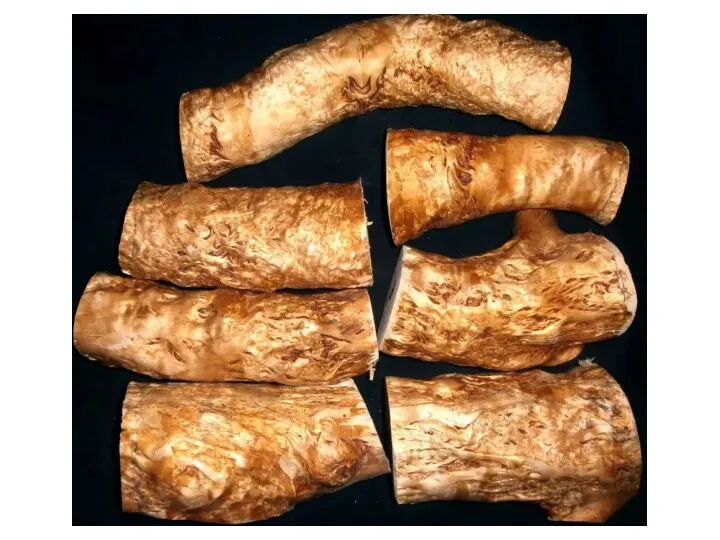
- 9. Ямчатость стебля у сосны обыкновенной Синдром ямчатости стебля у сосны обыкновенной связан с развитием наплывов на
- 12. По нашим наблюдениям, частота возникновения наплывов в ленточных борах Алтая связана с положением древостоев в рельефе
- 14. Вероятнее всего, что перечисленные неблагоприятные факторы внешней среды не являются непосредственной причиной возникновения наплывов, однако, в
- 15. Вероятно, денормализующий фактор как–то влияет на верхушечную меристему побега, в случае стволовых наплывов – лидирующего. Это
- 16. Анализ нескольких серий тангентальных и поперечных срезов, приготовленных из образцов древесины, включающих вместе с сердцевиной зону
- 18. Сама по себе ямчатость древесины предполагает наличие локальных зон замедленного прироста ("ямок"), однако "ямки" располагаются на
- 19. Наплывы могут возникать в любой части стебля и в любом возрасте растения, первые анатомически различимые изменения
- 20. Ямчатость стебля у березы повислой В Московской области мы обнаружили несколько экземпляров берёзы повислой с хорошо
- 21. После образования цепочки сомкнутых лучей паренхимная зона быстро, в течение одного вегетационного периода, расширяется. Ее увеличение
- 23. Ямчатость стебля у ольхи серой Ямчатость стебля у ольхи серой (Alnus incana Moench) ранее не была
- 24. Сначала эти трахеиды ориентированы радиально, затем, в процессе дальнейшего прироста древесины дифференцируются более длинные трахеальные элементы,
- 26. На второй год после образования “желобка” его дно оказывается составленным из коротких волокнистых трахеид, тяжевой паренхимы
- 27. Камбиальные производные, откладывающиеся в зоне развития аномальных лучей в строну луба отличаются от нормальных участков этой
- 28. Лущеный шпон этой древесины весьма декоративен. Анатомические исследования показали, что такой тип аномального строения также представляет
- 30. Одновременно с расширением лучей и образованием паренхиматизированной зоны на поверхности древесинного цилиндра появляется постепенно увеличивающееся продолговатое
- 31. ВИДЫ КЛЕНА (ACER L.) С ТЕКСТУРОЙ ДРЕВЕСИНЫ ТИПА "ПТИЧИЙ ГЛАЗ" Общее описание Коммерческое название древесины "птичий
- 32. Из этого был сделан вывод, что причиной формирования аномальной текстуры является угнетение роста. Ф. Рихтер (Richter,
- 33. Заметный вклад в изучение явора, формирующего аномальную древесину "птичий глаз" внесен исследователями из МЛТИ (Зуихина, Ирзун,
- 34. Проведенные исследования пока не позволяют с достаточной уверенностью объяснять причины формирования древесины типа "птичий глаз". Можно
- 36. Изменения в строении древесины при формировании текстуры "птичий глаз" начинаются, по нашим наблюдениям, с изменений в
- 37. В результате структурных изменений ограниченного участка меристематической ткани в древесине формируется аномальный луч, состоящий из живых
- 38. После образования паренхимной прослойки аномальный луч в процессе дальнейшего радиального прироста уменьшается или расчленяется на несколько
- 40. Приведенное описание внутреннего строения древесины "птичий глаз" сделано на основании изучения образцов из карпатских популяций явора.
- 42. Дальневосточный “"птичий глаз"” своеобразен и анатомическим строением. Аномальные зоны в древесине явора, как было отмечено, формируются
- 43. Аномальная древесина клена остролистного по макроструктурным признакам отличается от ранее описанных. В древесине остролистного клена зоны
- 45. Скачать презентацию

Общие положения
Особенности строения древесины и луба карельской березы, как это видно
Общие положения
Особенности строения древесины и луба карельской березы, как это видно



Причина возникновения ямчатости на поверхности древесинного цилиндра не во всех случаях
Причина возникновения ямчатости на поверхности древесинного цилиндра не во всех случаях

При этом наряду с ямчатостью в местах соединения компонентов прививки возникают
При этом наряду с ямчатостью в местах соединения компонентов прививки возникают

В результате изменений в лучевых инициалях происходит расширение лучей, их слияние
В результате изменений в лучевых инициалях происходит расширение лучей, их слияние

Ямчатость стебля у сосны обыкновенной
Синдром ямчатости стебля у сосны обыкновенной связан
Ямчатость стебля у сосны обыкновенной
Синдром ямчатости стебля у сосны обыкновенной связан


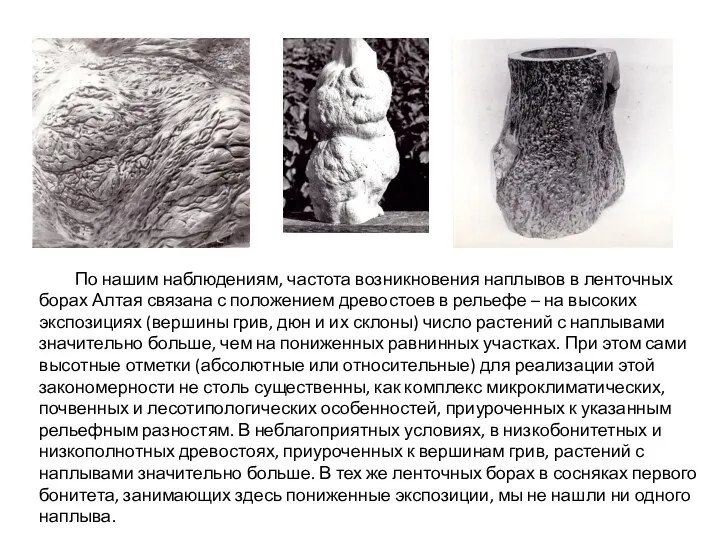
По нашим наблюдениям, частота возникновения наплывов в ленточных борах Алтая связана
По нашим наблюдениям, частота возникновения наплывов в ленточных борах Алтая связана


Вероятнее всего, что перечисленные неблагоприятные факторы внешней среды не являются непосредственной
Вероятнее всего, что перечисленные неблагоприятные факторы внешней среды не являются непосредственной

Вероятно, денормализующий фактор как–то влияет на верхушечную меристему побега, в случае
Вероятно, денормализующий фактор как–то влияет на верхушечную меристему побега, в случае

Анализ нескольких серий тангентальных и поперечных срезов, приготовленных из образцов древесины,
Анализ нескольких серий тангентальных и поперечных срезов, приготовленных из образцов древесины,
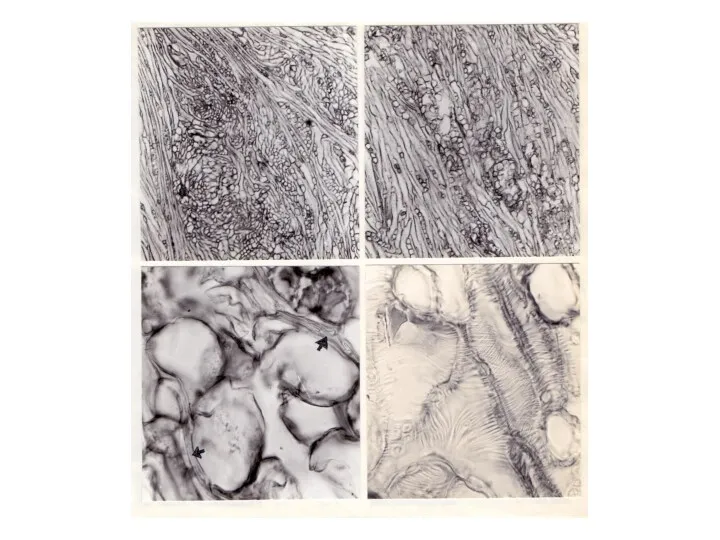

Сама по себе ямчатость древесины предполагает наличие локальных зон замедленного прироста
Сама по себе ямчатость древесины предполагает наличие локальных зон замедленного прироста

Наплывы могут возникать в любой части стебля и в любом возрасте
Наплывы могут возникать в любой части стебля и в любом возрасте

Ямчатость стебля у березы повислой
В Московской области мы обнаружили несколько экземпляров
Ямчатость стебля у березы повислой
В Московской области мы обнаружили несколько экземпляров

После образования цепочки сомкнутых лучей паренхимная зона быстро, в течение одного
После образования цепочки сомкнутых лучей паренхимная зона быстро, в течение одного
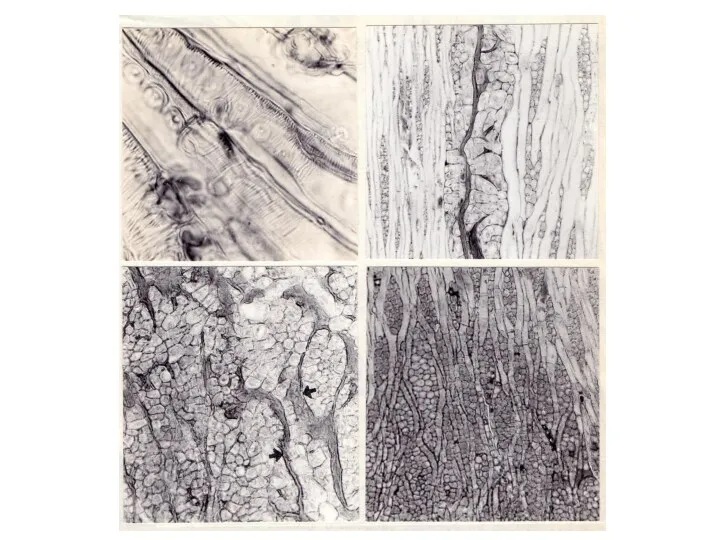

Ямчатость стебля у ольхи серой
Ямчатость стебля у ольхи серой (Alnus incana
Ямчатость стебля у ольхи серой
Ямчатость стебля у ольхи серой (Alnus incana

Сначала эти трахеиды ориентированы радиально, затем, в процессе дальнейшего прироста древесины
Сначала эти трахеиды ориентированы радиально, затем, в процессе дальнейшего прироста древесины


На второй год после образования “желобка” его дно оказывается составленным из
На второй год после образования “желобка” его дно оказывается составленным из

Камбиальные производные, откладывающиеся в зоне развития аномальных лучей в строну луба
Камбиальные производные, откладывающиеся в зоне развития аномальных лучей в строну луба

Лущеный шпон этой древесины весьма декоративен. Анатомические исследования показали, что такой
Лущеный шпон этой древесины весьма декоративен. Анатомические исследования показали, что такой
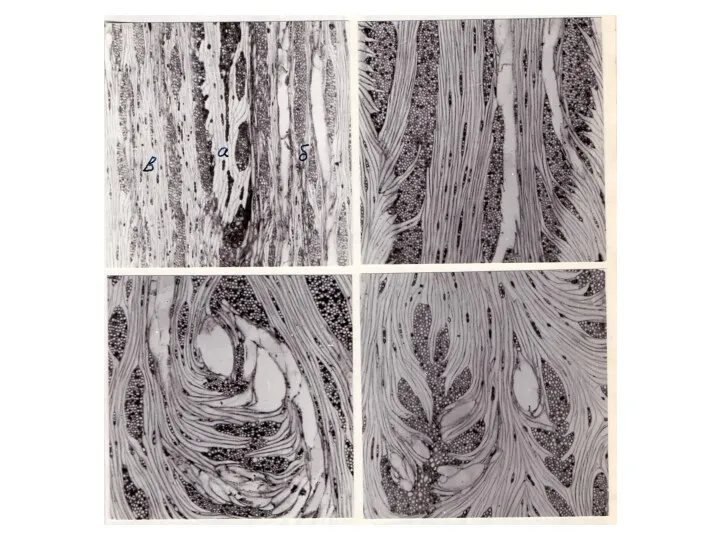

Одновременно с расширением лучей и образованием паренхиматизированной зоны на поверхности древесинного
Одновременно с расширением лучей и образованием паренхиматизированной зоны на поверхности древесинного

ВИДЫ КЛЕНА (ACER L.) С ТЕКСТУРОЙ ДРЕВЕСИНЫ ТИПА
"ПТИЧИЙ ГЛАЗ"
Общее описание
Коммерческое
ВИДЫ КЛЕНА (ACER L.) С ТЕКСТУРОЙ ДРЕВЕСИНЫ ТИПА
"ПТИЧИЙ ГЛАЗ"
Общее описание
Коммерческое

Из этого был сделан вывод, что причиной формирования аномальной текстуры является
Из этого был сделан вывод, что причиной формирования аномальной текстуры является

Заметный вклад в изучение явора, формирующего аномальную древесину "птичий глаз" внесен
Заметный вклад в изучение явора, формирующего аномальную древесину "птичий глаз" внесен

Проведенные исследования пока не позволяют с достаточной уверенностью объяснять причины формирования
Проведенные исследования пока не позволяют с достаточной уверенностью объяснять причины формирования
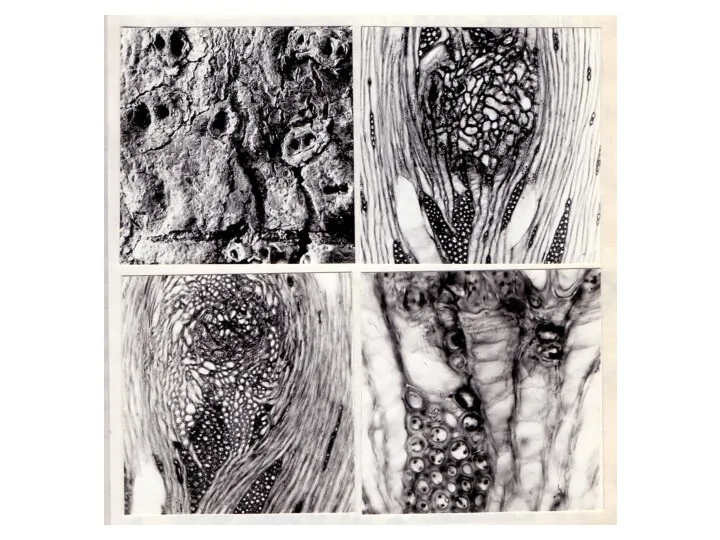

Изменения в строении древесины при формировании текстуры "птичий глаз" начинаются, по
Изменения в строении древесины при формировании текстуры "птичий глаз" начинаются, по

В результате структурных изменений ограниченного участка меристематической ткани в древесине формируется
В результате структурных изменений ограниченного участка меристематической ткани в древесине формируется

После образования паренхимной прослойки аномальный луч в процессе дальнейшего радиального прироста
После образования паренхимной прослойки аномальный луч в процессе дальнейшего радиального прироста


Приведенное описание внутреннего строения древесины "птичий глаз" сделано на основании изучения
Приведенное описание внутреннего строения древесины "птичий глаз" сделано на основании изучения
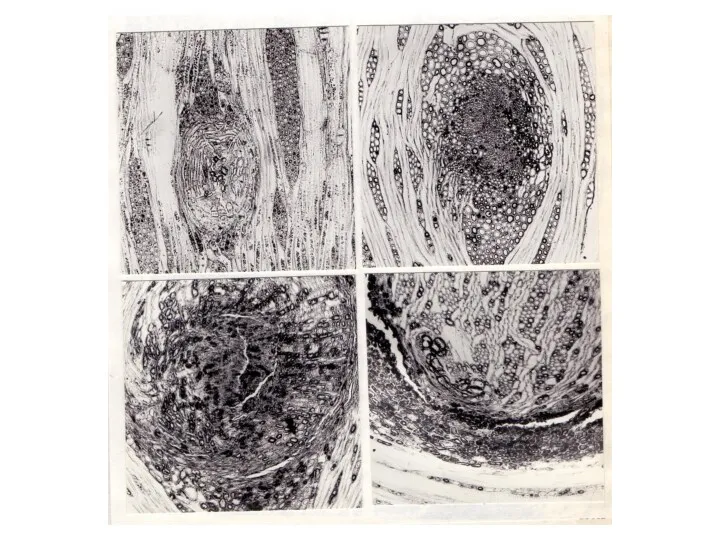

Дальневосточный “"птичий глаз"” своеобразен и анатомическим строением. Аномальные зоны в древесине
Дальневосточный “"птичий глаз"” своеобразен и анатомическим строением. Аномальные зоны в древесине

Аномальная древесина клена остролистного по макроструктурным признакам отличается от ранее описанных.
Аномальная древесина клена остролистного по макроструктурным признакам отличается от ранее описанных.
 Сравнение животной и растительной клетки. Ткани растений
Сравнение животной и растительной клетки. Ткани растений Растениеводство. Урок 48. Окружающий мир
Растениеводство. Урок 48. Окружающий мир Биологические модели развития популяций
Биологические модели развития популяций Микробиота тела человека. Роль микроорганизмов в возникновении инфекций. Способы передачи инфекций
Микробиота тела человека. Роль микроорганизмов в возникновении инфекций. Способы передачи инфекций Когнитивные функции мозга. Память и внимание
Когнитивные функции мозга. Память и внимание Семечки: вред или польза
Семечки: вред или польза Видоизменения побегов
Видоизменения побегов Современные проблемы медицинской краниологии
Современные проблемы медицинской краниологии игра-викторина по биологии для учащихся 5-6 классов
игра-викторина по биологии для учащихся 5-6 классов Різноманітність кісткових риб
Різноманітність кісткових риб Таргетинг генов
Таргетинг генов синтез белка и нук. кислот
синтез белка и нук. кислот 10 самых необычных грибов
10 самых необычных грибов Ткани растений
Ткани растений Интегрированная система защиты чечевицы от болезней, вредителей и сорняков
Интегрированная система защиты чечевицы от болезней, вредителей и сорняков Питание – это
Питание – это Тварини минулого
Тварини минулого Организменный уровень организации жизни
Организменный уровень организации жизни Эволюция опорно – двигательной системы у животных
Эволюция опорно – двигательной системы у животных Спинной мозг
Спинной мозг Молекулярная биология. Задачи
Молекулярная биология. Задачи Искусственные экосистемы. Аквариум
Искусственные экосистемы. Аквариум Мезозойская эра
Мезозойская эра Педагогика 21 века на пороге школы.
Педагогика 21 века на пороге школы. Powdery mildew of grape
Powdery mildew of grape В гости к весне
В гости к весне Отличия одноклеточных и многоклеточных организмов
Отличия одноклеточных и многоклеточных организмов Фауна Самарской Луки
Фауна Самарской Луки