- Эмбриональные стволовые клетки

Содержание
- 2. Эмбриональные стволовые клетки (самая ранняя стадия развития эмбриона) еще не вырабатывают антигены тканевой несовместимости и потому
- 3. Опыты Джона Гердона (1962) впервые показали, что возможен не только прямой, но и обратный переход: Стволовые
- 4. Дж. Гёрдон,1962 – первый опыт по клонированию ядро из клетки кишечника шпорцевой лягушки альбиноса животных http://www.nature.com/nm/journal/v15/n10/
- 5. Соавтор Гердона по нобелевской премии 2012 г. Яманаки пошел еще дальше – он открыл 4 гена,
- 6. Эксперименты Яманаки (2006). Гены четырех белков –регуляторов (1) вставляют в уже дифференцированный фибробласт (клетка кожи), полученный
- 7. 2012: Группа японских исследователей во главе с профессором Митинори Сайто из Университета Киото впервые в истории
- 8. Профессор Ян Вильмут с овечкой Долли (прожила 6 лет оставив потомство)
- 9. Лекция Асланяна М.М. Ян Вильмут из 250 зигот получил одну овечку Долли (1996 - 2003)
- 10. KIlOHbl 1952 1996 1997 1998 2000 2001 2002 2003 2007 Лекция Д.Ребрикова ?
- 11. 2008 - первое коммерческое клонирование собаки (лабрадор) 2009 – первая неудачная попытка клонирования вымершего животного (пиренейского
- 12. 2013 – проект по воскрешению вымерших гигантских птиц (дронт, моа, эпиорнис) пока не достиг успеха Эпиорнис
- 13. Моа из Новой Зеландии, как и прочих птиц-гигантов истребил человек теперь он пытается вернуть утраченное…
- 14. Удастся ли воскресить мамонта?
- 15. Проблема воссоздания вымерших видов животных не только в отсутствии всей ядерной и митохондриальной ДНК данного вида,
- 16. Еще один важный объект генетики развития: Гомеозисные гены - (регуляторные эмбриональные гены) определяют процессы роста и
- 17. Гомеозисные гены животных, грибов и растений могут содержать участок (гомеобокс) почти одинаковый у всех видов (180
- 18. Нох-гены найдены у всех исследованных организмов (в геномах губок, гидры, пиявок, нематод, амфибий , рыб, млекопитающих).
- 19. В эмбриональных клетках существует сложное каскадное включение транскрипционных факторов: разные конц-и генов материнского эффекта включают разные
- 20. Несмотря на разнообразие структуры у цветка, его развитие разных видов контролируется консервативными гомеозисными генами Anthirrhinum Matthiola
- 21. Фолиарная (классическая) теория морфогенеза цветка Иоганна Вольфганга Гете: 1749 - 1832 Цветок – видоизмененный побег с
- 22. ABC-модель развития органов цветка (Coen, 2 Meyerowitz, 1991) 1 3 4 Ч Л Т П Ч
- 23. АВС –модель – современная парадигма генетики развития в области ботаники
- 24. Гомеозисные мутанты арабидопсис Мутация В-класса гена Мутация гена А-класса Мутация гена С-класса
- 25. ap1 ap3 ag Дикий тип Фенотип тройного мутанта – убедительное свидетельство в пользу фолиарной теории морфогенеза
- 26. Гомеозисные мутации найдены и у дрозофилы Муха дикого http://www.tmd.ac.jp/artsci/biol/ типа textlife/fruitfly.jpg Мутант bithorax http://www.mun.ca/biology/scarr/Bithorax_ Drosophila.html Кэлвин
- 27. мутация antennapedia) антенна → нога (доминантная трансформация передних разделов в задние из-за того, что у этого
- 28. Одинаков порядок расположения генов на хромосоме 3 и порядок расположения контролируемых ими органов на теле будущей
- 29. Ген Hox c-6 контролирует образование грудного отдела позвоночника У курицы ген Hoxc-6 отвечает за образование 7
- 30. Сравнение экспрессии Hox-генов в курином и мышином эмбрионе выявляют различия в протяженности (но не порядке!) доменов
- 31. Гомеобоксные НОХ-гены считаются «краеугольным камнем» процессов развития у животных. Эдвард Льюис, Кристиа Нюсляйн- Фольхард и касающиеся
- 32. НОХ- гены конечностей. фенотип кластеров А и D размечают разные участки О их роли в этом
- 33. Мутации полидактилии выявлены у разных организмов. Фенотипическое сходство связано с мутациями в ортологичных НОХ-генах.
- 34. М НОХ - белки НОХ-гены включают сотни генов – мишеней Как регулируется работа самих НОХ-генов? Органогенез/
- 35. У хордовых и позвоночных в регуляции экспрессии генов важную роль играет ретиноевая кислота (производное от витамина
- 36. Синтез ретиноевой кислоты (РК) АЛКОГОЛЬДЕГИДРОГЕНАЗА ЭТАНОЛ
- 37. Эти данные объясняют, почему употребление алкоголя во время беременности ведет к выраженным аномалиям у новорожденных, которые
- 38. Изменения экспрессии регуляторных генов, контролирующих развитие, приводят к нарушению развития (уродства). Однако эти нарушения, как правило,
- 39. Однако есть и особый тип модификаций, которые могут передаваться через деления клеток и даже сохраняться потомками.
- 40. Эпигенетические изменения - немутационные изменения в геноме (иногда наследственные). Пример: метилирование цитозина с участием интерферирующей (микроРНК)
- 41. В среднем, 1% всей ДНК человека метилирован!
- 42. ое ы и Вид метилирования деметилирования 5’-GGCGACTGCGATGCCATGCGTT- -CCGCTGACGCTACGGTACGCAA- me me me me me -GGCGACTGCGATGCCATGCGTT- -CCGCTGACGCTACGGTACGCAA- -GGCGACTGCGATGCCATGCGTT-
- 43. Основные функции метилирования ДНК 1.) Регуляция экспрессии генов на уровне транскрипции ДНК 2.) Распознавание и блокировка
- 44. – активация AP-2 CCGG -80 -70 Пример: AP-2 активирует транскрипцию гена проэнкефалина человека AP-2 me CCGG
- 45. Эпигенетические изменения связаны главным образом с процессами метилирования ДНК и гистоновых белков, которое осуществляется специальными ферментами
- 46. Главное отличие эпигенетических механизмов регуляции работы генов от других механизмов, которые используются в регуляции развития, заключается
- 47. Кастовая структура общественных насекомых также связана с эпигенетическими механизмами У пчел матка (царица), нянька и сборщица
- 48. Внешняя среда вступает в диалог с геномом уже во время внутриутробного развития Монозиготные близнецы имеют одинаковый
- 50. Скачать презентацию

Эмбриональные стволовые клетки (самая ранняя стадия развития эмбриона) еще не вырабатывают
Эмбриональные стволовые клетки (самая ранняя стадия развития эмбриона) еще не вырабатывают
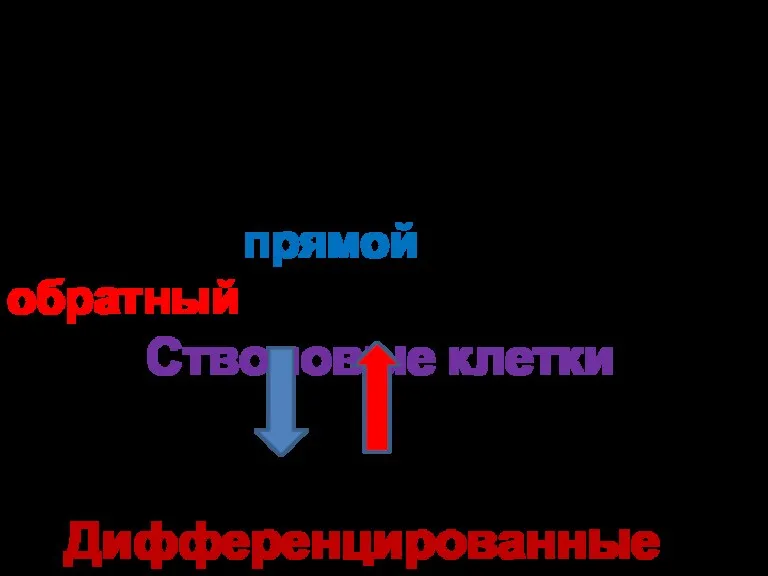
Опыты Джона Гердона (1962)
впервые показали, что возможен
не только прямой, но
Опыты Джона Гердона (1962)
впервые показали, что возможен
не только прямой, но

Дж.
Гёрдон,1962
–
первый
опыт
по клонированию
ядро из клетки
кишечника шпорцевой лягушки
альбиноса
животных
http://www.nature.com/nm/journal/v15/n10/
fig_tab/nm1009-1141_F1.html
2012
Дж.
Гёрдон,1962
–
первый
опыт
по клонированию
ядро из клетки
кишечника шпорцевой лягушки
альбиноса
животных
http://www.nature.com/nm/journal/v15/n10/
fig_tab/nm1009-1141_F1.html
2012

Соавтор Гердона по нобелевской премии 2012 г. Яманаки пошел еще дальше
Соавтор Гердона по нобелевской премии 2012 г. Яманаки пошел еще дальше

Эксперименты Яманаки (2006). Гены четырех белков –регуляторов (1) вставляют в уже
Эксперименты Яманаки (2006). Гены четырех белков –регуляторов (1) вставляют в уже

2012: Группа японских исследователей во главе с профессором Митинори Сайто
2012: Группа японских исследователей во главе с профессором Митинори Сайто

Профессор Ян Вильмут с овечкой Долли
(прожила 6 лет оставив потомство)
Профессор Ян Вильмут с овечкой Долли
(прожила 6 лет оставив потомство)
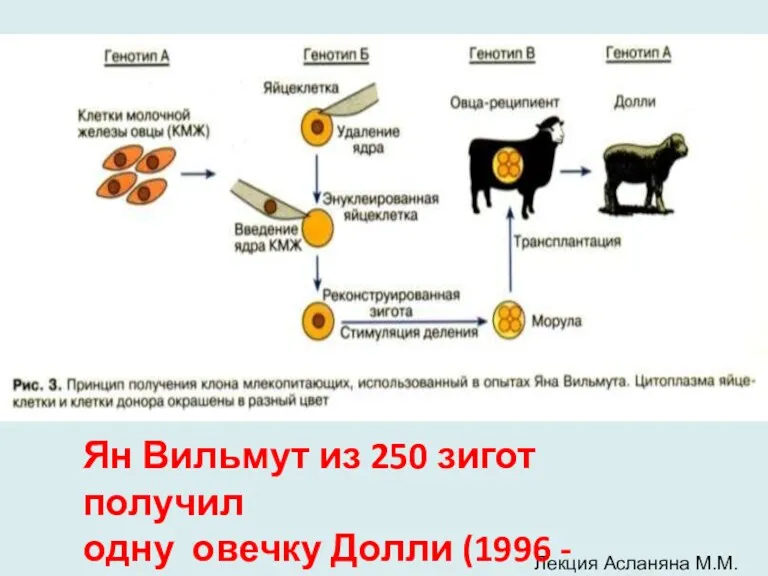
Лекция Асланяна М.М.
Ян Вильмут из 250 зигот получил
одну овечку Долли (1996
Лекция Асланяна М.М.
Ян Вильмут из 250 зигот получил
одну овечку Долли (1996

KIlOHbl
1952
1996
1997
1998
2000
2001
2002
2003
2007
Лекция Д.Ребрикова
?
KIlOHbl
1952
1996
1997
1998
2000
2001
2002
2003
2007
Лекция Д.Ребрикова
?

2008 - первое коммерческое
клонирование собаки (лабрадор)
2009 – первая неудачная попытка
2008 - первое коммерческое
клонирование собаки (лабрадор)
2009 – первая неудачная попытка

2013 – проект по воскрешению вымерших гигантских птиц (дронт, моа, эпиорнис)
2013 – проект по воскрешению вымерших гигантских птиц (дронт, моа, эпиорнис)

Моа из Новой Зеландии,
как и прочих
птиц-гигантов истребил человек
теперь
Моа из Новой Зеландии,
как и прочих
птиц-гигантов истребил человек
теперь

Удастся ли воскресить
мамонта?
Удастся ли воскресить
мамонта?

Проблема воссоздания вымерших видов животных не только в отсутствии всей ядерной
Проблема воссоздания вымерших видов животных не только в отсутствии всей ядерной

Еще один важный объект
генетики развития: Гомеозисные гены -
(регуляторные эмбриональные
Еще один важный объект
генетики развития: Гомеозисные гены -
(регуляторные эмбриональные

Гомеозисные гены животных, грибов и растений могут содержать участок (гомеобокс) почти
Гомеозисные гены животных, грибов и растений могут содержать участок (гомеобокс) почти
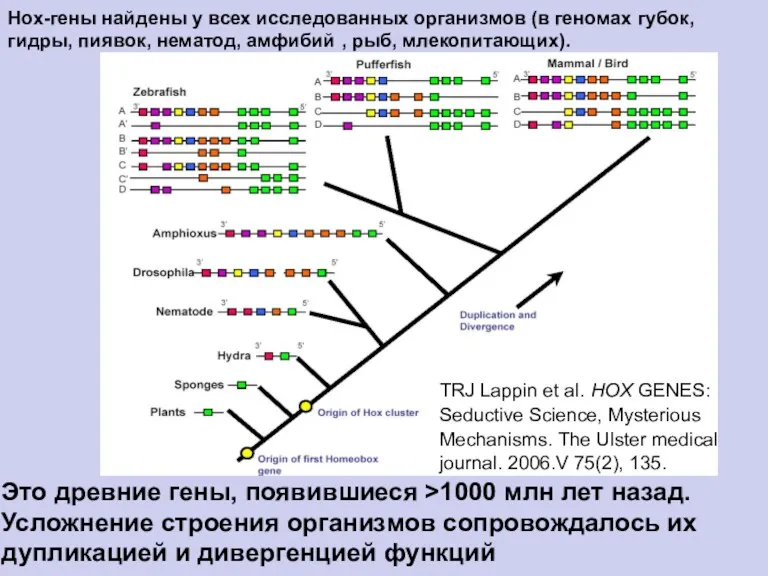
Нох-гены найдены у всех исследованных организмов (в геномах
губок,
гидры, пиявок,
нематод,
амфибий
,
рыб, млекопитающих).
TRJ Lappin
Нох-гены найдены у всех исследованных организмов (в геномах
губок,
гидры, пиявок,
нематод,
амфибий
,
рыб, млекопитающих).
TRJ Lappin

В эмбриональных клетках существует сложное каскадное включение транскрипционных факторов: разные конц-и
В эмбриональных клетках существует сложное каскадное включение транскрипционных факторов: разные конц-и

Несмотря
на
разнообразие
структуры
у
цветка,
его
развитие
разных
видов
контролируется
консервативными
гомеозисными
генами
Anthirrhinum
Matthiola
Arabis
Syringa
Trillium
Несмотря
на
разнообразие
структуры
у
цветка,
его
развитие
разных
видов
контролируется
консервативными
гомеозисными
генами
Anthirrhinum
Matthiola
Arabis
Syringa
Trillium
Фолиарная
(классическая) теория морфогенеза
цветка
Иоганна
Вольфганга
Гете:
1749 - 1832
Цветок – видоизмененный побег с укороченными междоузлиями.
Органы
Фолиарная
(классическая) теория морфогенеза
цветка
Иоганна
Вольфганга
Гете:
1749 - 1832
Цветок – видоизмененный побег с укороченными междоузлиями.
Органы
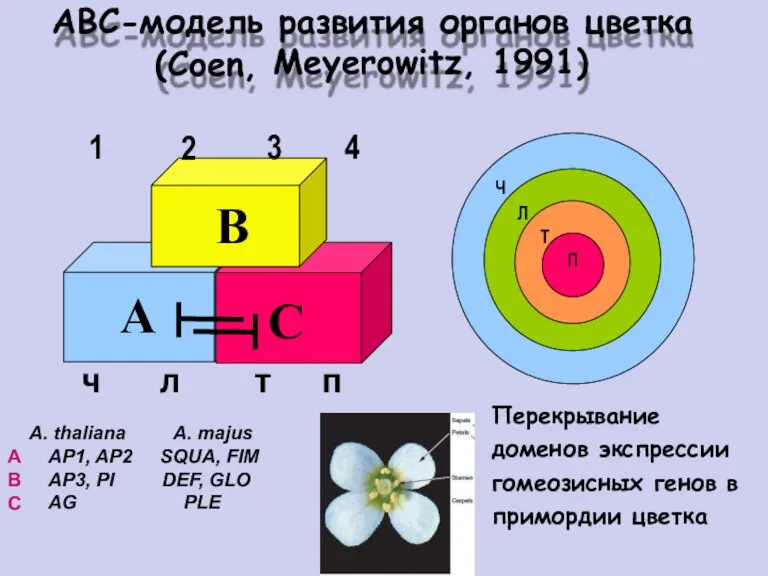
ABC-модель развития
органов
цветка
(Coen,
2
Meyerowitz,
1991)
1
3
4
Ч
Л
Т
П
Ч
Л
Т
П
Перекрывание
доменов экспрессии
гомеозисных генов в
примордии цветка
A. thaliana
AP1, AP2
AP3, PI
AG
A. majus
SQUA, FIM
DEF,
ABC-модель развития
органов
цветка
(Coen,
2
Meyerowitz,
1991)
1
3
4
Ч
Л
Т
П
Ч
Л
Т
П
Перекрывание
доменов экспрессии
гомеозисных генов в
примордии цветка
A. thaliana
AP1, AP2
AP3, PI
AG
A. majus
SQUA, FIM
DEF,
АВС –модель –
современная
парадигма
генетики развития
в области
ботаники
АВС –модель –
современная
парадигма
генетики развития
в области
ботаники

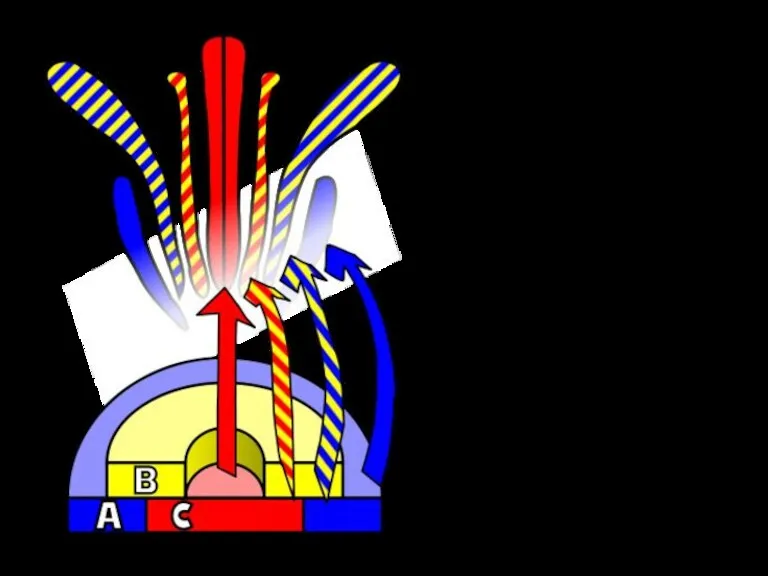
Гомеозисные
мутанты
арабидопсис
Мутация
В-класса
гена
Мутация гена
А-класса
Мутация гена
С-класса
Гомеозисные
мутанты
арабидопсис
Мутация
В-класса
гена
Мутация гена
А-класса
Мутация гена
С-класса

ap1
ap3
ag
Дикий
тип
Фенотип тройного
мутанта
– убедительное свидетельство
в пользу фолиарной теории морфогенеза цветка Гете
ap1
ap3
ag
Дикий
тип
Фенотип тройного
мутанта
– убедительное свидетельство
в пользу фолиарной теории морфогенеза цветка Гете

Гомеозисные
мутации
найдены
и
у
дрозофилы
Муха
дикого
http://www.tmd.ac.jp/artsci/biol/
типа
textlife/fruitfly.jpg
Мутант
bithorax
http://www.mun.ca/biology/scarr/Bithorax_
Drosophila.html
Кэлвин Бриджес 1915
Гомеозисные
мутации
найдены
и
у
дрозофилы
Муха
дикого
http://www.tmd.ac.jp/artsci/biol/
типа
textlife/fruitfly.jpg
Мутант
bithorax
http://www.mun.ca/biology/scarr/Bithorax_
Drosophila.html
Кэлвин Бриджес 1915

мутация antennapedia)
антенна →
нога
(доминантная
трансформация передних разделов в
задние
из-за того, что у этого мутанта
мутация antennapedia)
антенна →
нога
(доминантная
трансформация передних разделов в
задние
из-за того, что у этого мутанта

Одинаков порядок расположения генов на хромосоме
3 и порядок расположения контролируемых
ими
органов
на
теле
будущей
мухи
http://www.bio.miami.edu
/dana/pix/Hox_drosophila
.jpg
Одинаков порядок расположения генов на хромосоме
3 и порядок расположения контролируемых
ими
органов
на
теле
будущей
мухи
http://www.bio.miami.edu
/dana/pix/Hox_drosophila
.jpg
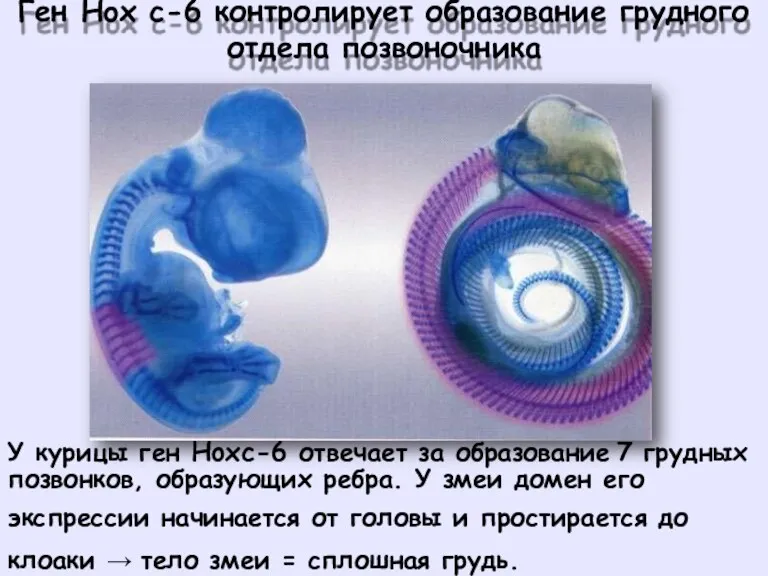
Ген
Hox
c-6
контролирует
образование
грудного
отдела
позвоночника
У курицы ген Hoxc-6 отвечает за образование
7 грудных
позвонков, образующих ребра. У
Ген
Hox
c-6
контролирует
образование
грудного
отдела
позвоночника
У курицы ген Hoxc-6 отвечает за образование
7 грудных
позвонков, образующих ребра. У
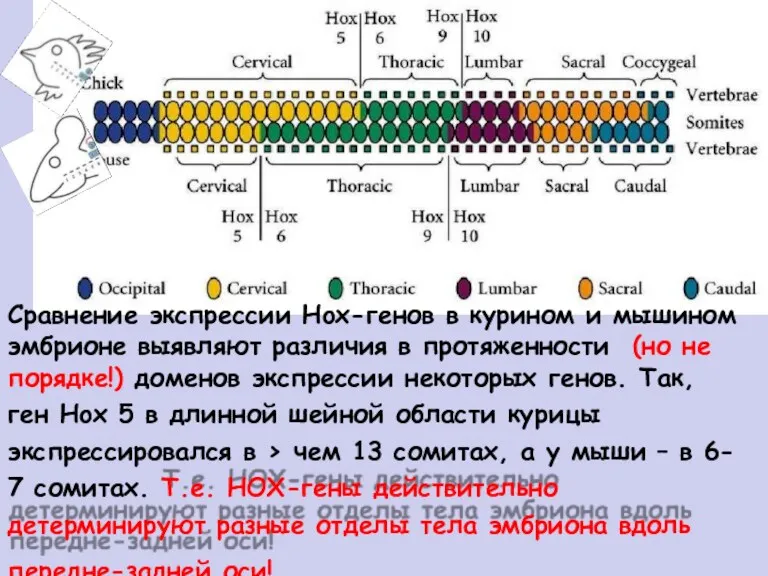
Сравнение экспрессии Hox-генов в курином и мышином
эмбрионе выявляют различия в протяженности
(но
Сравнение экспрессии Hox-генов в курином и мышином
эмбрионе выявляют различия в протяженности
(но

Гомеобоксные НОХ-гены считаются
«краеугольным камнем»
процессов
развития
у
животных.
Эдвард
Льюис,
Кристиа
Нюсляйн-
Фольхард и
касающиеся
Эрик
Вишаус
за
открытия,
генетического
контроля
на
ранней
стадии
эмбрионального
развития
НОХ-гены
включаются
в
онтогенезе
несколько раз.
Они не
только размечают
участки
тела
вдоль
передне-задней оси,
но играют
органов
важную
роль
в
развитии
многих
1995 г
Гомеобоксные НОХ-гены считаются
«краеугольным камнем»
процессов
развития
у
животных.
Эдвард
Льюис,
Кристиа
Нюсляйн-
Фольхард и
касающиеся
Эрик
Вишаус
за
открытия,
генетического
контроля
на
ранней
стадии
эмбрионального
развития
НОХ-гены
включаются
в
онтогенезе
несколько раз.
Они не
только размечают
участки
тела
вдоль
передне-задней оси,
но играют
органов
важную
роль
в
развитии
многих
1995 г
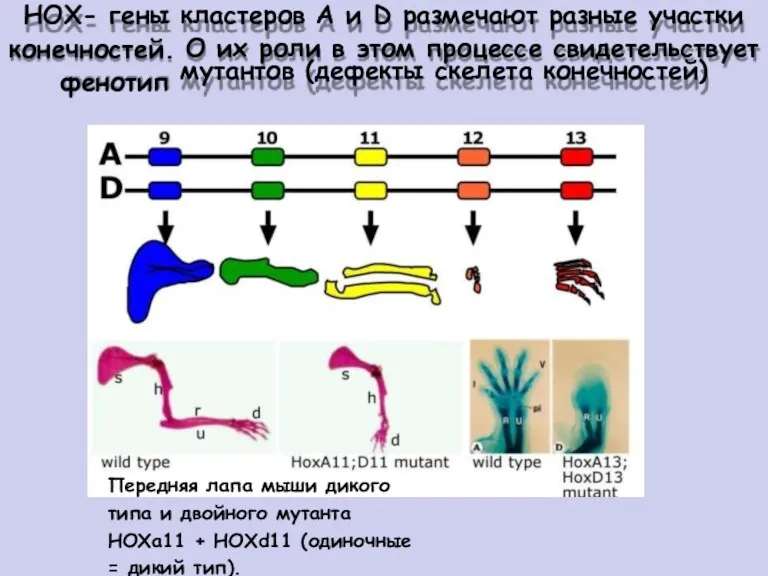
НОХ- гены
конечностей.
фенотип
кластеров А и D размечают разные участки
О их роли в
НОХ- гены
конечностей.
фенотип
кластеров А и D размечают разные участки
О их роли в
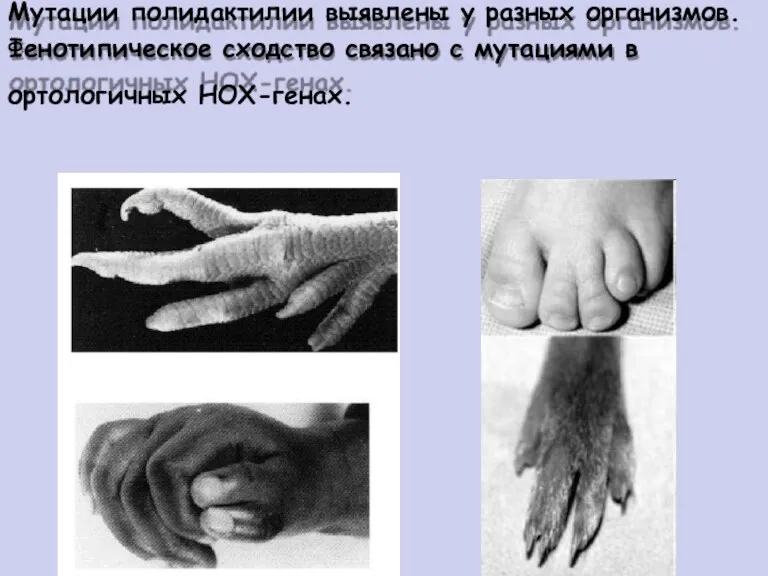
Мутации полидактилии выявлены у разных организмов.
Фенотипическое сходство связано с мутациями в
ортологичных
Мутации полидактилии выявлены у разных организмов.
Фенотипическое сходство связано с мутациями в
ортологичных
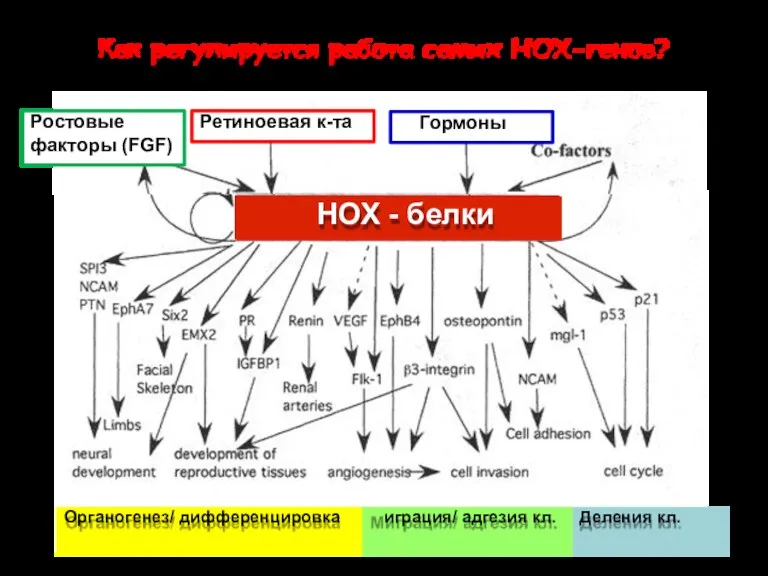
М
НОХ - белки
НОХ-гены
включают сотни генов – мишеней
Как регулируется работа самих НОХ-генов?
Органогенез/
НОХ - белки
НОХ-гены
включают сотни генов – мишеней
Как регулируется работа самих НОХ-генов?
Органогенез/
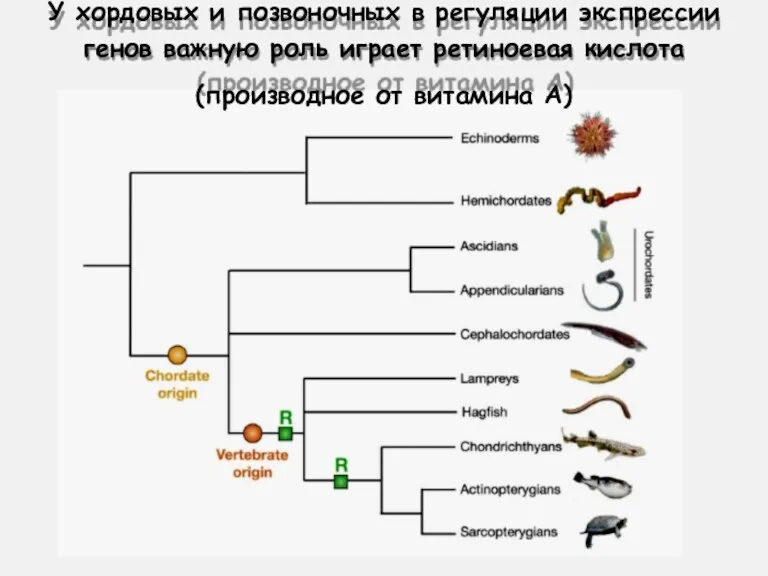
У хордовых и позвоночных в регуляции экспрессии
генов важную роль играет ретиноевая
У хордовых и позвоночных в регуляции экспрессии
генов важную роль играет ретиноевая
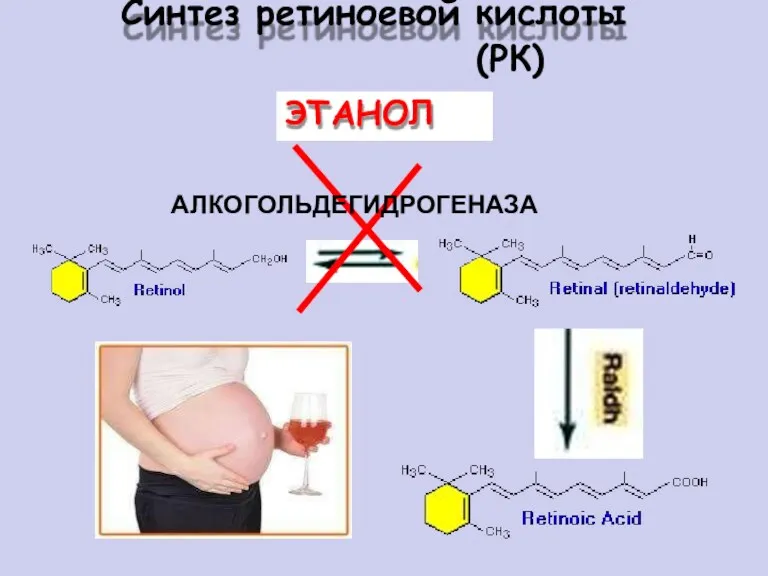
Синтез
ретиноевой
кислоты (РК)
АЛКОГОЛЬДЕГИДРОГЕНАЗА
ЭТАНОЛ
Синтез
ретиноевой
кислоты (РК)
АЛКОГОЛЬДЕГИДРОГЕНАЗА
ЭТАНОЛ
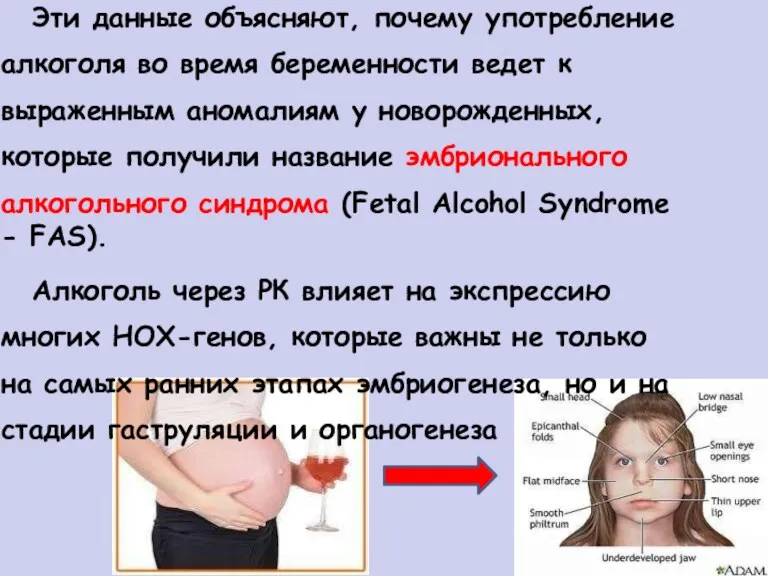
Эти данные объясняют, почему употребление
алкоголя во время беременности ведет к
Эти данные объясняют, почему употребление
алкоголя во время беременности ведет к

Изменения экспрессии
регуляторных генов,
контролирующих развитие, приводят к
нарушению развития (уродства). Однако эти
нарушения, как
Изменения экспрессии
регуляторных генов,
контролирующих развитие, приводят к
нарушению развития (уродства). Однако эти
нарушения, как

Однако есть и особый тип модификаций, которые
могут передаваться через деления клеток
Однако есть и особый тип модификаций, которые
могут передаваться через деления клеток

Эпигенетические изменения -
немутационные изменения в геноме (иногда наследственные). Пример: метилирование
Эпигенетические изменения -
немутационные изменения в геноме (иногда наследственные). Пример: метилирование
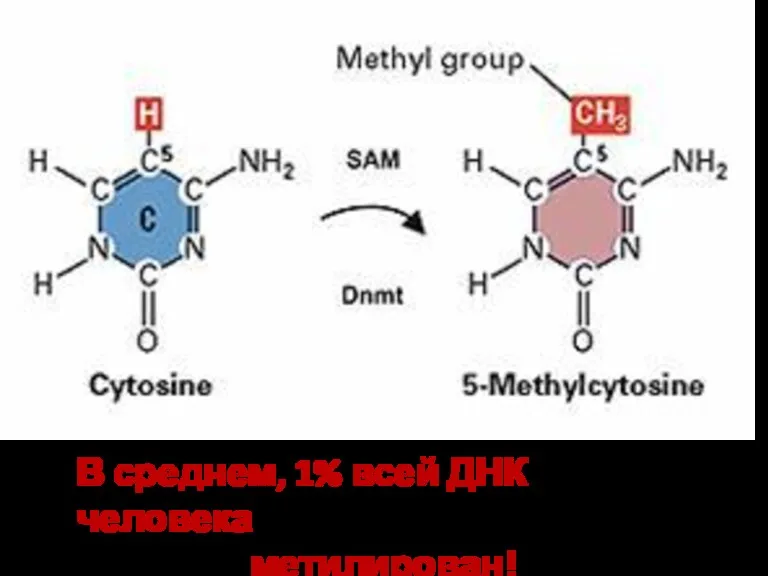
В среднем, 1% всей ДНК человека
метилирован!
В среднем, 1% всей ДНК человека
метилирован!
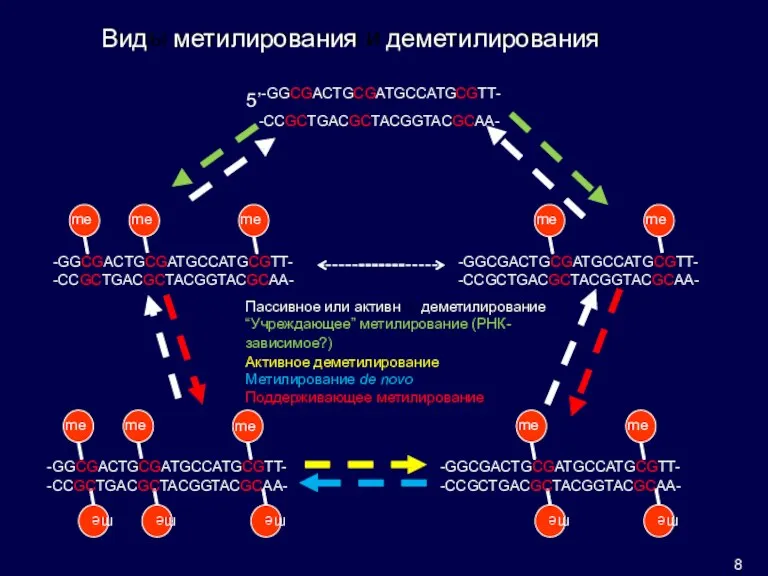
ое
ы
и
Вид
метилирования
деметилирования
5’-GGCGACTGCGATGCCATGCGTT-
-CCGCTGACGCTACGGTACGCAA-
me
me
me
me
me
-GGCGACTGCGATGCCATGCGTT-
-CCGCTGACGCTACGGTACGCAA-
-GGCGACTGCGATGCCATGCGTT-
-CCGCTGACGCTACGGTACGCAA-
Пассивное или активн
деметилирование
“Учреждающее” метилирование (РНК-
зависимое?)
Активное деметилирование Метилирование de novo Поддерживающее метилирование
me
me
me
me
me
-GGCGACTGCGATGCCATGCGTT-
-CCGCTGACGCTACGGTACGCAA-
-GGCGACTGCGATGCCATGCGTT-
-CCGCTGACGCTACGGTACGCAA-
8
me
me
me
me
me
ы
и
Вид
метилирования
деметилирования
5’-GGCGACTGCGATGCCATGCGTT-
-CCGCTGACGCTACGGTACGCAA-
me
me
me
me
me
-GGCGACTGCGATGCCATGCGTT-
-CCGCTGACGCTACGGTACGCAA-
-GGCGACTGCGATGCCATGCGTT-
-CCGCTGACGCTACGGTACGCAA-
Пассивное или активн
деметилирование
“Учреждающее” метилирование (РНК-
зависимое?)
Активное деметилирование Метилирование de novo Поддерживающее метилирование
me
me
me
me
me
-GGCGACTGCGATGCCATGCGTT-
-CCGCTGACGCTACGGTACGCAA-
-GGCGACTGCGATGCCATGCGTT-
-CCGCTGACGCTACGGTACGCAA-
8
me
me
me
me
me

Основные функции метилирования ДНК
1.) Регуляция экспрессии генов на уровне транскрипции ДНК
Основные функции метилирования ДНК
1.) Регуляция экспрессии генов на уровне транскрипции ДНК
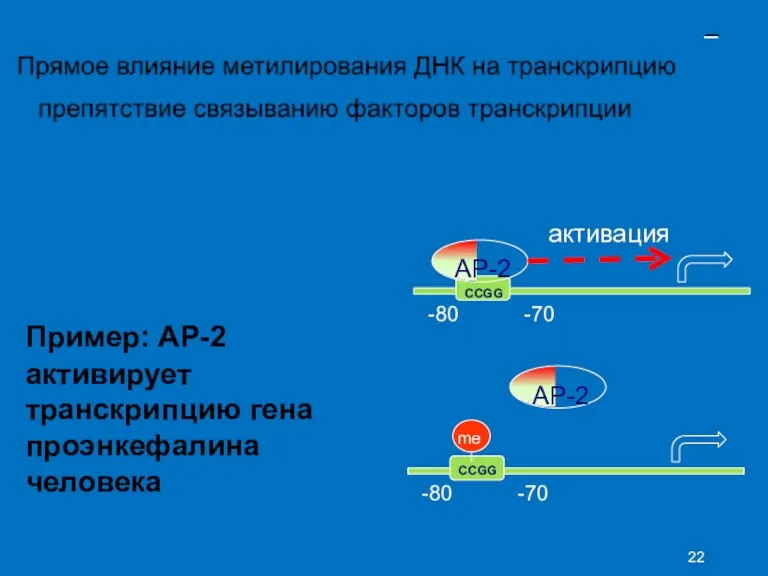
–
активация
AP-2
CCGG
-80
-70
Пример: AP-2
активирует транскрипцию гена проэнкефалина человека
AP-2
me
CCGG
-80
-70
22
–
активация
AP-2
CCGG
-80
-70
Пример: AP-2
активирует транскрипцию гена проэнкефалина человека
AP-2
me
CCGG
-80
-70
22
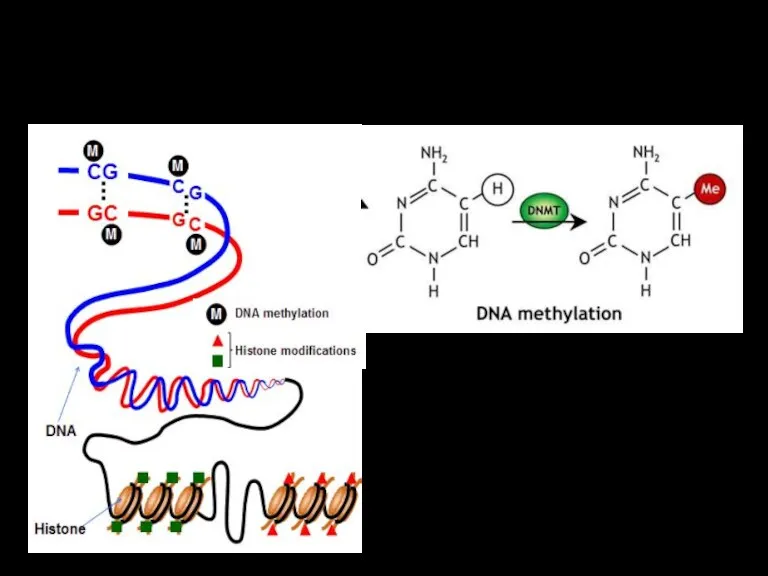
Эпигенетические изменения связаны главным образом с
процессами метилирования ДНК и гистоновых белков,
которое
осуществляется
специальными
ферментами
Это
Эпигенетические изменения связаны главным образом с
процессами метилирования ДНК и гистоновых белков,
которое
осуществляется
специальными
ферментами
Это

Главное отличие эпигенетических механизмов
регуляции работы генов от других механизмов,
которые используются в
Главное отличие эпигенетических механизмов
регуляции работы генов от других механизмов,
которые используются в

Кастовая структура общественных насекомых также
связана с эпигенетическими механизмами
У пчел матка (царица),
Кастовая структура общественных насекомых также
связана с эпигенетическими механизмами
У пчел матка (царица),
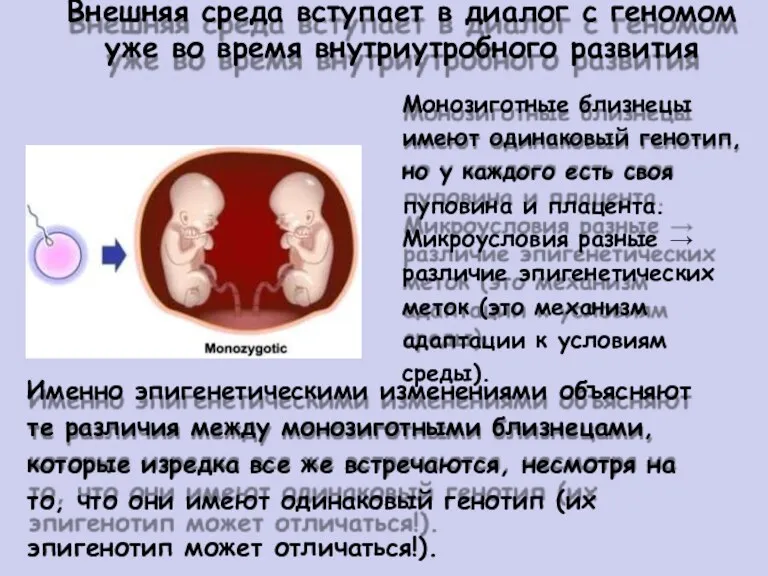
Внешняя
среда
вступает
в
диалог
с
геномом
уже
во
время
внутриутробного
развития
Монозиготные близнецы
имеют одинаковый генотип,
но у каждого есть своя
пуповина и плацента.
Микроусловия разные
Внешняя
среда
вступает
в
диалог
с
геномом
уже
во
время
внутриутробного
развития
Монозиготные близнецы
имеют одинаковый генотип,
но у каждого есть своя
пуповина и плацента.
Микроусловия разные
 Дневные хищные, совы, куриные
Дневные хищные, совы, куриные Понятие об организме. Доядерные и ядерные организмы. Клетка и её открытие
Понятие об организме. Доядерные и ядерные организмы. Клетка и её открытие Цианобактерии. Отличия от бактерий
Цианобактерии. Отличия от бактерий Амин қышқылдар. Дәрумендер
Амин қышқылдар. Дәрумендер Кити і дельфіни
Кити і дельфіни Методологія еволюційної систематики та історія макросистем
Методологія еволюційної систематики та історія макросистем Экологические сообщества. Экосистема, биогеоценоз
Экологические сообщества. Экосистема, биогеоценоз Происхождение растений. Основные этапы развития растительного мира
Происхождение растений. Основные этапы развития растительного мира Морской котик
Морской котик Птицы – лесные жители. 2 класс
Птицы – лесные жители. 2 класс Нервная система
Нервная система Луи Пастер (1822-1895)
Луи Пастер (1822-1895) Одомашнивание животных
Одомашнивание животных Генетический материал клетки. Генный уровень
Генетический материал клетки. Генный уровень Std ppwt fertilizers and oil
Std ppwt fertilizers and oil Общая характеристика насекомых
Общая характеристика насекомых Биосфера эволюциясы
Биосфера эволюциясы Психофизиологические аспекты поведения человека
Психофизиологические аспекты поведения человека Клеточный цикл. Деление клетки
Клеточный цикл. Деление клетки Солнце, растения и мы с вами
Солнце, растения и мы с вами Маңызды мәдени өсімдіктер іріктемелері мен үй жануарлары қолтұқымдары
Маңызды мәдени өсімдіктер іріктемелері мен үй жануарлары қолтұқымдары презентация Углеводы. Липиды
презентация Углеводы. Липиды Endangered animals
Endangered animals Эндокринная система. Железы внутренней секреции
Эндокринная система. Железы внутренней секреции Можно ли жить без воды
Можно ли жить без воды Поширення плодів та насіння у природі
Поширення плодів та насіння у природі Как называются деревья?
Как называются деревья? Цели и задачи биотехнологии. История развития биотехнологии. Слагаемые биотехнологического процесса
Цели и задачи биотехнологии. История развития биотехнологии. Слагаемые биотехнологического процесса