- Энергетический обмен. Взаимосвязь обмена веществ и энергии. Лекция №3

Содержание
- 2. План лекции: Общие закономерности метаболизма. Этапы катаболизма. Цепь переноса электронов (ЦПЭ), состав, строение компонентов дыхательной цепи.
- 3. ОБМЕН ВЕЩЕСТВ «ОБМЕН ВЕЩЕСТВ или метаболизм - совокупность всех химических изменений и всех видов превращений веществ
- 4. Промежуточный обмен (внутриклеточный метаболизм) включает два типа реакций: Катаболизм – биосинтетические процессы расщепления органических молекул до
- 5. Промежуточный обмен (внутриклеточный метаболизм) включает два типа реакций: Анаболизм - биосинтетические процессы, в которых простые строительные
- 7. Процессы окисления протекают в организме и вне организма. Эти процессы имеют сходства и различия. Биологическое окисление
- 8. Сходство между окислением в организме и вне организма. В результате окисления образуются одинаковые конечные продукты СО2
- 9. Различия между окислением в организме и вне организма. Вне организма энергия выделяется за счет окисления атомов
- 10. Различия между окислением в организме и вне организма. Вне организма энергия выделяется одномоментно и не аккумулируется,
- 11. Различия между окислением в организме и вне организма. Основной реакцией окисления в организме является реакция дегидрирования,
- 12. Утилизация энергии в виде макроэргических связей При разрыве макроэргической связи образуется более 30 кДж/моль свободной энергии
- 13. Центральную роль в энергетическом обмене играет АТФ Фосфорилирование АДФ в клетках происходит путем присоединения неорганического фосфата
- 15. Аденозинтрифосфорная кислота АТФ АТФ функционирует как общий промежуточный продукт, переносящий энергию от реакций, сопровождающихся выделением свободной
- 16. Типы фосфорилирования АДФ В зависимости от источника энергии, обеспечивающего присоединение фосфатного участка, выделяют два типа фосфорилирования
- 17. Субстратное фосфорилирование - это процесс синтез АТФ из АДФ и неорганического фосфата за счет энергии, выделившийся
- 19. Окислительное фосфорилирование АДФ - синтез АТФ из АДФ и Н3РО4 за счет переноса электронов по ЦПЭ
- 20. Типы биологического окисления анаэробное окисление (акцептором Н+ и ē служит не О2, а другой субстрат «субстратное
- 21. Тканевое дыхание Тканевым дыханием являются реакции окисления органических веществ, в которых используется кислород и образуется вода
- 22. Цепь переноса электронов (ЦПЭ) ЦПЭ – это комплекс оксидоредуктаз, локализованных на внутренней мембране митохондрий, участвующих в
- 23. Первичные акцепторы водорода Никотинамидзависимые дегидрогеназы (NAD, NADР) (находятся в матриксе митохондрий или в цитозоле) NAD (NADР)
- 24. Первичные акцепторы водорода Флавинзависимые дегидрогеназы (FAD, FMN) В отличие от NAD FAD соединен прочно с белковой
- 26. Состав ЦПЭ NADH – дегидрогеназа (комплекс I) Сукцинатдегидрогеназа – (FAD - кофермент) (комплекс II) QH2 –
- 27. NADH – дегидрогеназа (комплекс I) Комплекс I ЦПЭ содержит ФМН и 5 железосерных белков. NADН-дегидрогеназа (комплекс
- 28. Сукцинатдегидрогеназа – (FAD - кофермент) (комплекс II) Сукцинатдегидрогеназа - СДГ (FAD – зависимая дегидрогеназа) (комплекс II)
- 29. Бензохиноновое соединение локализуется во внутренней мембране митохондрий и собирает электроны с комплекса I и со всех
- 30. QH2 (коэнзим Q –дегидрогеназа) (комплекс III) Состоит из 2-типов цитохромов (b и с1) и железосерного белка
- 31. Низкомолекулярные переносчики электронов Цитохром с водорастворимый мембранный белок; ММ 12,5 кД, одна полипептидная цепь из 100
- 32. Цитохром-c-оксидаза (цит а/а3) (комплекс IV) Комплекс IV ЦПЭ содержит цитохромы а, а3, два иона меди. Это
- 33. АТФ-синтаза (комплекс V) Возвращение Н+ из межмембранного пространства в матрикс происходит с помощью фермента АТФ-синтаза (комплекс
- 34. (II) Пути поступления электронов и протонов в ЦПЭ от первичных доноров
- 35. Распределение энергии, выделяемой ЦПЭ ~ 20% ~ 40-50 % ~ 30%
- 36. + 0,8 При движении от окисляемого субстрата к кислороду электроны теряют часть своей потенциальной энергии. Эту
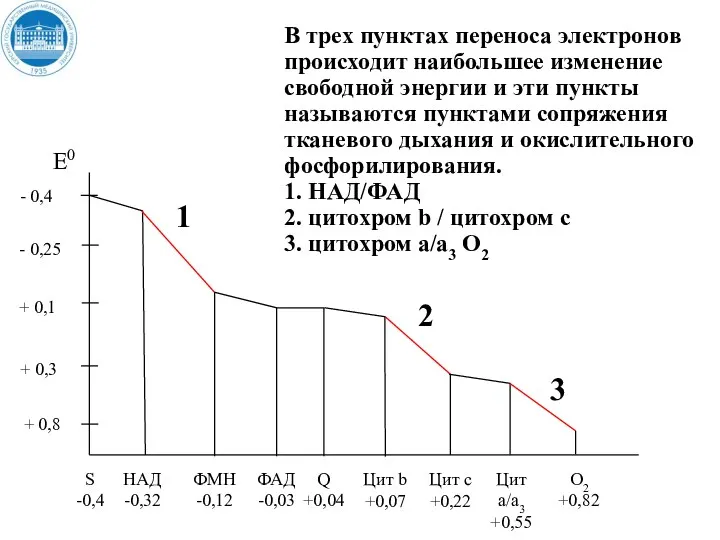
- 37. + 0,8 В трех пунктах переноса электронов происходит наибольшее изменение свободной энергии и эти пункты называются
- 38. Пункты сопряжения постоянны, но их количество зависит от природы окисляемого субстрата. При окисления НАД – зависимых
- 39. Коэффициент окислительного фосфорилирование Р/О показывает, какое количество неорганического фосфата затрачивается для фосфорилирования АДФ при использовании 1
- 40. Коэффициент окислительного фосфорилирования НАД-Н+ + ½О2 →НАД+ + Н2О 3 АДФ + 3 Н3РО4→ 3 АТФ
- 41. Причины нарушения биологического окисления Недостаток субстрата (голодание, пищева-рение, межуточное нарушение обменов белков, углеводов, липидов); Недостаток О2
- 42. Вещества, понижающие интенсивность окислительного фосфолирирования АДФ. Ингибиторы ферментов ЦПЭ подавляют активность ферментных комплексов I, II и
- 43. Примеры веществ, понижающие интенсивность окислительного фосфолирирования АДФ. ингибиторы NADH-дегидрогиназы - лекарственные препараты со снотворным действием –
- 44. Вещество, способное полностью блокировать тканевое дыхание, синильная кислота Синильная кислота является веществом, вызывающим кислородное голодание тканевого
- 45. Вещество, способное полностью блокировать тканевое дыхание, синильная кислота Соединившись с цианидом, цитохромоксидаза теряет способность переносить электроны
- 46. Вещества, понижающие интенсивность окислительного фосфолирирования АДФ. Ингибиторы АТФ-синтазы снижают активность фермента, скорость фосфорилирования АДФ и коэффициент
- 47. Вещества, понижающие интенсивность окислительного фосфолирирования АДФ. Разобщители окисления и фосфорилирования – липофильные протонофоры способны легко проникать
- 48. Лекция № 4 Обмен углеводов: значение, переваривание. Гликолиз. Пентозофосфатный путь окисления глюкозы.
- 49. ПЛАН ЛЕКЦИИ: Обмен углеводов. Гликолиз. Пентозофосфатный путь окисления глюкозы.
- 50. Обмен углеводов в организме человека складываются из следующих процессов: Расщепление в желудочно-кишечном тракте до моносахаридов поступающих
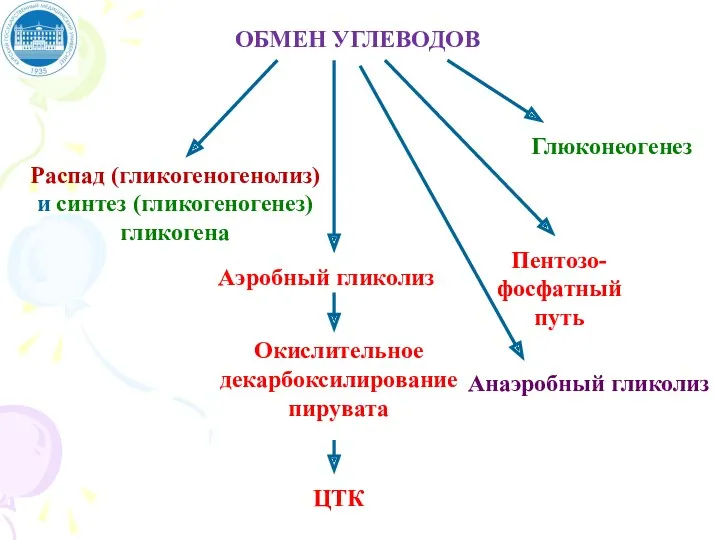
- 51. ОБМЕН УГЛЕВОДОВ Распад (гликогеногенолиз) и синтез (гликогеногенез) гликогена Аэробный гликолиз Окислительное декарбоксилирование пирувата ЦТК Пентозо- фосфатный
- 52. Две основные функции: Углеводы – источник углеродов, который необходим для синтеза ряда соединений (белков, нуклеиновых кислот,
- 53. Другие функции: Резервная (крахмал, гликоген). Структурная (полисахариды образуют прочный остов в комплексе с белками и липидами,
- 54. Переваривание углеводов в организме Источником углеводов для организма служат углеводы пищи - крахмал, сахароза и лактоза.
- 55. Переваривание углеводов Амилаза слюны расщепляет α-1,4-гликозидные связи в крахмале. В ротовой полости происходит лишь частичное переваривание
- 56. Переваривание углеводов Последующее переваривание нерасщепленного или частично расщепленного крахмала происходит в кишечнике. В двенадцатиперстной кишке pH
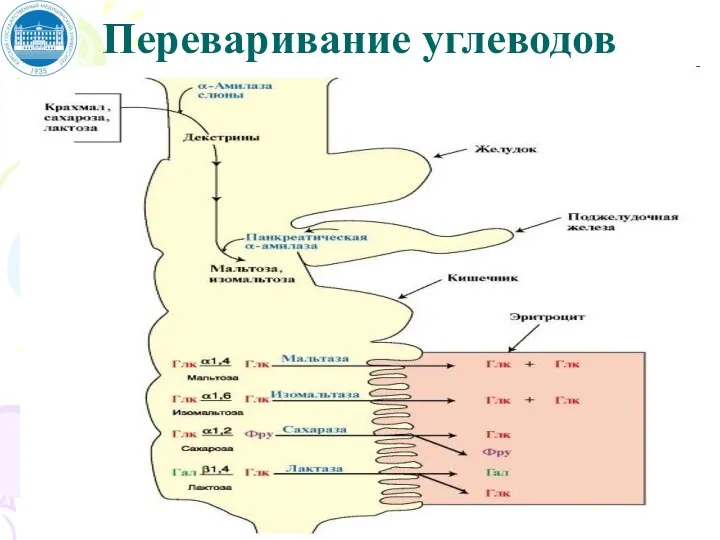
- 57. Переваривание углеводов Мальтоза и изомальтоза вместе с другими пищевыми дисахаридами - сахарозой и лактозой - гидролизуются
- 58. Переваривание углеводов
- 59. Продукты полного переваривания углеводов – глюкоза, галактоза и фруктоза – через клетки кишечника поступают в кровь.
- 60. Гликолиз Гликолиз (от греч. glykys – сладкий, lysys – распад ) – один центральных путей катаболизма
- 61. Гликолиз включает превращения трех разных типов: Распад углеродного скелета глюкозы с образованием пирувата ( путь атомов
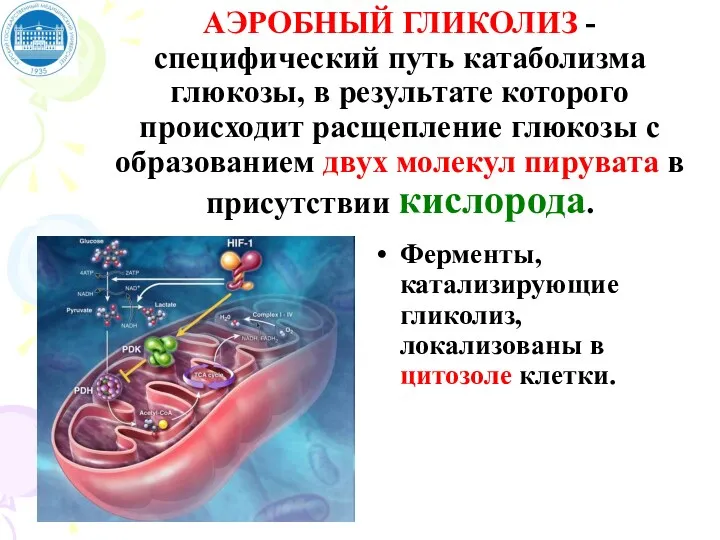
- 62. АЭРОБНЫЙ ГЛИКОЛИЗ - специфический путь катаболизма глюкозы, в результате которого происходит расщепление глюкозы с образованием двух
- 63. Стадии гликолиза I. 1 реакция. Фосфорилирование глюкозы: реакция протекает необратимо, катализируется глюкокиназой или гексокиназой и требует
- 64. Ферменты участвующие в фосфорилировании глюкозы. Связывание гексокиназы с гексозой происходит по типу индуцированного соответствия: молекула фермента
- 65. Второй реакцией глюколиза является превращение глюкозо-6-фосфата под действием фермента глюкозо-6-фосфатизомеразы во фруктозо-6-фосфат:
- 66. Третья реакция катализируется ферментом фосфофруктокиназой; образовавшийся фруктозо-6-фосфат вновь фосфорилируется за счет второй молекулы АТФ: Данная реакция
- 67. Четвертую реакцию гликолиза катализирует фермент альдолаза. Под влиянием этого фермента фруктозо-1,6-бисфосфат расщепляется на две фосфотриозы: 5%
- 68. Пятая реакция – это реакция изомеризации триозофосфатов. Катализируется ферментом триозофосфатизомеразой:
- 69. II. На второй стадии гликолиза происходит запасание энергии. Из одной молекулы ГЛЮ образуется две молекулы глицеральдегид-3-фосфата,
- 70. Седьмая реакция катализируется фосфоглицераткиназой, при этом происходит передача богатого энергией фосфатного остатка (фосфатной группы в положении
- 71. Восьмая реакция сопровождается внутримолекулярным переносом оставшейся фосфатной группы, и 3-фосфоглицерат превращается 2-фосфоглицерат. Фермент – фосфоглицерамутаза. Реакция
- 72. Девятая реакция катализируется ферментом енолазой, при этом 2-фосфоглицерат в результате отщепления молекулы воды переходит в фосфоенолпируват,
- 73. Десятая реакция характеризуется разрывом высокоэргической связи и переносом фосфатного остатка от фосфоенолпирувата на АДФ (субстратное фосфорилирование).
- 74. Конечным продуктом аэробного гликолиза является пируват, а энергетический баланс складывается из 2 молекул АТФ образовавшихся в
- 75. Глицерофосфатный челночный механизм Цитоплазма Митохондрии дигидрокси- ацетонфосфат Глицерол-3-фосфат 2 2 2НАД·Н + Н+ 2НАД + ФАД+
- 76. Малат-аспартатный челночный механизм Цитоплазма Митохондрии ЩУК малат НАД·Н + Н+ НАД + глутамат аспартат глутамат аспартат
- 77. Баланс аэробного гликолиза 1 реакция - 1 АТФ 3 реакция - 1 АТФ 6 реакция +
- 78. АНАЭРОБНЫЙ ГЛИКОЛИЗ - специфический путь катаболизма глюкозы, в результате которого происходит расщепление глюкозы с образованием двух
- 79. Дальше процесс идет в зависимости от наличия или отсутствия кислорода в клетке: При анаэробных условиях, например
- 80. В этих условиях образовавшийся при гликолизе НАДН регенерируется за счет пирувата, который восстанавливается до лактата. Электроны,
- 81. Баланс анаэробного гликолиза 1 реакция - 1 АТФ 3 реакция - 1 АТФ 6 реакция -
- 82. Пентозофосфатный путь окисления глюкозы Локализация: молочная железа, эмбриональная и жировая ткань, эритроциты, легкие, щитовидная железа, мозг,
- 83. В пентозофосфатном пути превращения глюкозы можно выделить: Окислительный этап Поставляет клеткам кофермент NADPH (использующийся как донор
- 84. Глюкоза Глюкозо -6-фосфат Фруктозо -6-фосфат Глицеральдегид-3-фосфат Гликолиз 2НАДФ+ 2НАДФН·Н+ Окислительная фаза Синтез жирных кислот Синтез стероидов
- 85. Условия протекания пентозофосфатного пути окисления глюкозы Судьба глюкозо-6-фосфата — вступит ли он в гликолиз или пентозофосфатный
- 86. НАДФ+ НАДФН+Н+ Глюкозо-6-фосфатдегидрогеназа Глюконатлактон-гидратаза Н2О НАДФ+ НАДФН+Н+ 6-Фосфоглюконатдегидрогеназа СО2 Реакции окислительного этапа пентозофосфатного пути
- 87. Патология пентозофосфатного пути окисления глюкозы Генетический дефект фермента глюкозо-6-фосфатдегидрогеназы является причиной: усиления процессов перекисного окисления липидов
- 88. Лекция № 5 Специфические пути катаболизма. Общий путь катаболизма: окислительное декарбоксилирование пирувата и цикл трикарбоновых кислот.
- 89. Специфические и общий пути катаболизма
- 90. Специфические и общий пути катаболизма Начальные этапы катаболизма (специфические пути катаболизма) основных пищевых веществ (белков, жиров
- 91. Суммарное уравнение реакции окислительного декарбоксилирования пирувата: Эту реакцию катализирует сложно организованный пируватдегидрогеназный комплекс (ПДК). Реакция протекает
- 92. Состав пируватдегидрогеназного комплекса Ферменты: Е1 - пируватдекарбоксилаза Е2 - дигидролипоилтрансацетилаза Е3 - дигидролипоилдегидрогеназа Коферменты: ТДФ –
- 93. Цикл трикарбоновых кислот Реакции цитратного цикла происходят в матриксе митохондрий.
- 94. Цикл трикарбоновых кислот Цикл Кребса — это ключевой этап дыхания всех клеток, использующих кислород, центр пересечения
- 95. Цитратный цикл (цикл трикарбоновых кислот) представляет собой совокупность 8 последовательных химических реакций, в ходе которых происходят:
- 96. В I-й реакции под действием цитратсинтазы происходят конденсация ацетильного остатка ацетил-КоА с оксалоацетатом (щавелевоуксусная кислота -
- 97. Далее цитрат в две стадии (дегидратация и последующая гидратация по двойной связи) превращается в изоцитрат. Промежуточным
- 98. В III- й реакции под действием НАД+- зависимой изоцитратдегидрогеназы происходят окисление и декарбоксилирование изоцитрата с образованием
- 99. В IV–й реакции происходит окислительное декарбоксилирование α – кетоглутарата с выделением еще одной молекулы СО2 и
- 100. Вторая половина цикла – V –я реакция превращения сукцинил – КоА в сукцинат (янтарная кислота) ,
- 101. VI реакция. Сукцинат под действием ФАД – зависимой сукцинатдегидрогеназы превращается в фумарат (фумаровая кислота). Реакция окислительного
- 102. VII реакция гидратации. К фумарату фермент фумараза (фумаратгидратаза) присоединяет молекулу воды и образуется малат (яблочная кислота).
- 103. В заключительной VIII реакции цикла происходит дегидрирование малата НАД+ - зависимым ферментом малатдегидрогеназой и образование оксалоацетата.
- 104. Функции цикла ЦТК Интегративная функция — цикл является связующим звеном между реакциями анаболизма и катаболизма. Катаболическая
- 105. Функции цикла ЦТК Водорододонорная функция — цикл Кребса поставляет на дыхательную цепь митохондрий протоны в виде
- 106. Таким образом, в ОПК происходит распад 3 – углеродного соединения пирувата с выделением 3 молекул СО2.
- 107. Баланс АТФ при окислении глюкозы до С2О и Н2О I. Гликолиз А) 1. глю→ глю-ф-ф –
- 108. Мнемоническое правило Для более легкого запоминания кислот, участвующих в цикле Кребса, существует мнемоническое правило: Целый Ананас
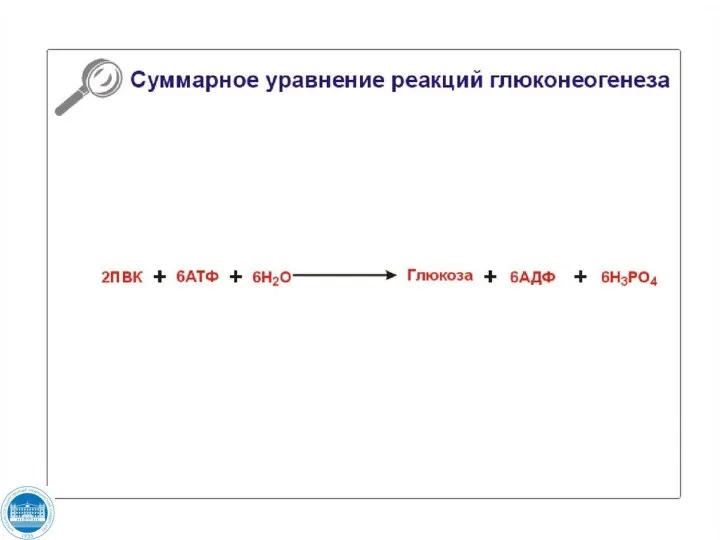
- 109. СИНТЕЗ ГЛЮКОЗЫ - ГЛЮКОНЕОГЕНЕЗ
- 110. Глюконеогенез - это процесс синтеза глюкозы из веществ неуглеводной природы. Субстратами глюконеогенеза являются: пируват, лактат, глицерол,
- 111. Важнейшей функцией глюконеогенеза является: поддержание уровня глюкозы в крови в период длительного голодания интенсивных физических нагрузок.
- 112. Глюконеогенез протекает главным образом в печени менее интенсивно - в корковом веществе почек, в слизистой оболочке
- 113. Включение различных субстратов в глюконеогенез зависит от физиологического состояния организма: лактат является продуктом анаэробного гликолиза в
- 114. Схема гликолиза и глюконеогенеза
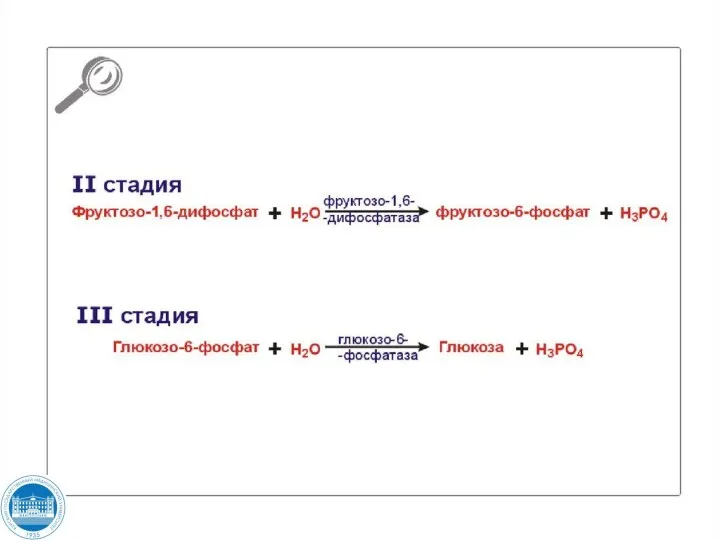
- 116. Глюконеогенез Большинство реакций гликолиза и глюконеогенеза являются обратимыми и катализируются одними и теми же ферментами. Четыре
- 117. пируват ОА ОА малат
- 118. ОА
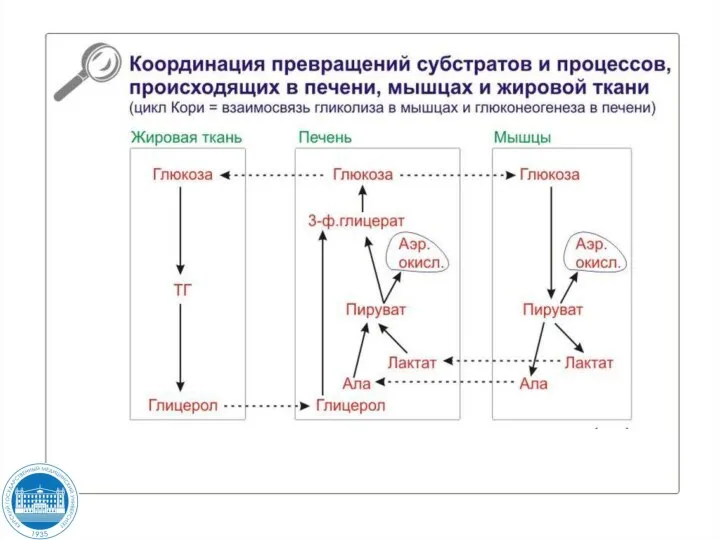
- 123. Цикл Кори Использование лактата в качестве субстрата в глюконеогенезе связано с транспортом его в печень и
- 124. Благодарю за внимание
- 125. Лекция № 6 Глюконеогенез (продолжение). Обмен гликогена. Особенности обмена углеводов в различных органах и тканях.
- 127. Включение глицерина в синтез глюкозы
- 128. Включение глицерина в синтез глюкозы
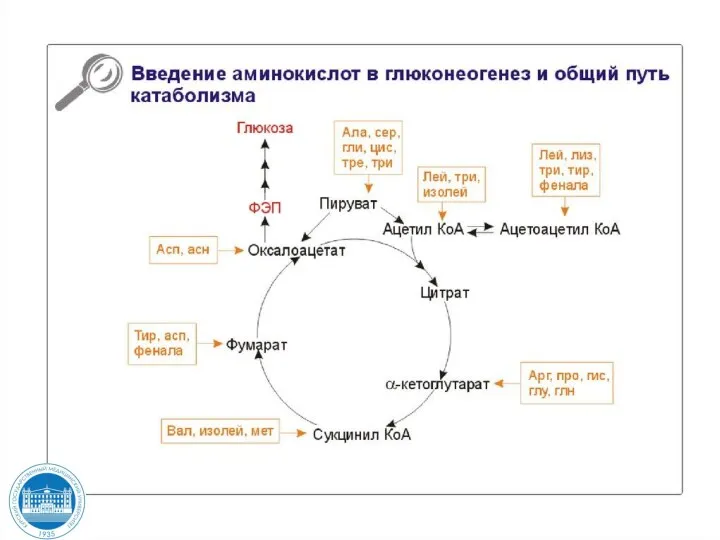
- 129. Включение аминокислот в синтез глюкозы
- 130. СИНТЕЗ ГЛИКОГЕНА (ГЛИКОГЕНОГЕНЕЗ), МОБИЛИЗАЦИЯ ГЛИКОГЕНА (ГЛИКОГЕНОЛИЗ).
- 131. Гликоген - основной резервный полисахарид в клетках животных Гликоген представляет собой разветвленный гомополисахарид, мономером которого является
- 132. Гликоген хранится в цитозоле клеток в форме гранул и депонируется главным образом в печени и скелетных
- 133. Гранулы гликогена плохо растворимы в воде и не влияют на осмотическое давление в клетке. Это обстоятельство
- 134. Синтез гликогена Гликоген синтезируется в период пищеварения (абсорбтивный период: 1-2 часа после приема углеводной пищи) в
- 135. Синтез гликогена Когда длина синтезируемой цепи увеличивается на 11-12 остатков глюкозы, фермент ветвления - глюкозил- 1,4-1,6-трансфераза
- 136. Синтез гликогена - гликогеногенез
- 137. Мобилизация (распад) гликогена Мобилизация (распад) гликогена происходит в интервалах между приемами пищи (постабсорбтивный период) и ускоряется
- 138. Мобилизация (распад) гликогена Мобилизация гликогена в печени отличается от таковой в мышцах одной реакцией (реакция 5),
- 139. Мобилизация (распад) гликогена - гликогеногенолиз
- 140. Гормоны, обеспечивающие переключение метаболических путей: Переключение процессов синтеза и мобилизации гликогена в печени и мышцах происходит
- 141. Регуляция метаболизма гликогена в печени Под влиянием инсулина происходит: ускорение транспорта глюкозы в клетки инсулинзависимых мышечной
- 142. Регуляция метаболизма гликогена в печени Под влиянием инсулина происходит: активация фосфопротеинфосфатазой гранул гликогена, которая дефосфорилирует гликогенсинтазу
- 143. Первичным сигналом для синтеза инсулина и глюкагона является изменение концентрации глюкозы в крови. Инсулин и глюкагон
- 144. Регуляция метаболизма гликогена в печени В период пищеварения концентрация глюкозы в крови повышается до 10-12 ммоль/л,
- 145. Особенности обмена глюкозы в различных тканях и органах
- 146. ОСОБЕННОСТИ УГЛЕВОДНОГО ОБМЕНА В ПЕЧЕНИ (ГЕПАТОЦИТЫ) 1. Превращение фруктозы и галактозы в глюкозу 2. Резервная роль
- 147. Обмен углеводов в печени Одной из важнейших функций печени в процессах обмена веществ является ее участие
- 148. Обмен углеводов в печени В печени катаболизм глюкозы представлен 2 процессами: 1) гликолитический путь превращения 1
- 149. Обмен углеводов в печени Гликолиз обеспечивает энергией клеточные реакции фосфорилирования, синтез белка; пентозофосфатный путь служит источником
- 150. Обмен углеводов в печени При аэробных условиях происходит сочетание гликолиза, протекающего в цитоплазме и цикла лимонной
- 151. Обмен углеводов в печени В печени протекают гликогенез и гликогенолиз. Эти процессы взаимосвязаны и регулируются как
- 152. Обмен углеводов в мышцах Цель мышечной клетки – наиболее эффективно использовать поступающую глюкозу для образования АТФ,
- 153. Обмен углеводов в мышцах Цитоплазма мышечных клеток содержит в высоких концентрациях ферменты гликолиза, а изобилие митохондрий
- 154. Обмен углеводов в мышцах В мышцах идет гликогенез, мышца осуществляет лишь немногие синтетические функции. Ключевые ферменты
- 155. Особенности обмена углеводов в мышцах ИНСУЛИН ГЛЮ ГЛЮ ГЛЮТ-4 ГЛЮ-6-Ф ПИРУВАТ АТФ ЦТК Ацетил - КоА
- 156. Обмен углеводов в мышцах Фосфорилирование глюкозы в мышцах происходит под дейстием гексокиназы, в печени этот процесс
- 157. Обмен углеводов в сердечной мышце Ни медленно сокращающаяся гладкая мышечная ткань, ни сердечная мышца не потребляют
- 158. Особенности обмена углеводов в сердце ЛАКТАТ Печень (глюконеогенез) Почки (выведение) ЛАКТАТ ЛАКТАТ ПИРУВАТ ГЛЮ-6-Ф ЛДГ4 (м3н)
- 159. Кровь Глю-6-ф Глю Кет. тела Ацетил-КоА ЖК ГФЛ Ацетилхолин Сфингофосфо- липиды Мембраны нейронов и миелиновый футляр
- 160. Обмен углеводов в мозге По сравнению со всеми органами тела функций мозга в наибольшей степени зависит
- 161. Обмен углеводов в мозге Высокая активность митохондриальных ферментов цикла лимонной кислоты предотвращает накопление лактата в тканях
- 162. Особенности обмена углеводов в жировой ткани ГЛЮТ-4 + ГЛЮ ГЛЮ ДФА α-ГФ ТАГ Ацил-КоА АДИПОЦИТ ПИРУВАТ
- 163. Обмен углеводов в эритроцитах Эритроциты не содержат ядра, митохондрий. В эритроците не идут реакции цикла лимонной
- 164. Особенности обмена углеводов в эритроците ГЛЮ ГЛЮ-6-Ф 1,3 ДФГ ПИРУВАТ ЛАКТАТ ЛАКТАТ НАД НАДНН+ Hb(Fe2+) Мет
- 165. Особенности обмена глюкозы в клетках опухoли В клетках опухоли отмечается повышенная активность гексокиназы, что приводит к
- 166. Особенности обмена глюкозы в клетках опухoли Мембранная гексокиназа – работает как насос. Гипогликемия. Анаэробный гликолиз. «Принудительный»
- 167. Благодарю за внимание
- 168. Лекция №7 Регуляция и патология углеводного обмена
- 169. Регуляция обмена углеводов состоит из регуляции: Соотношение между процессами катаболизма и анаболизма глюкозы в клетках печени
- 170. Регуляции гликолиза Основное значение гликолиза - синтез АТФ, поэтому его скорость должна коррелировать с затратами энергии
- 171. Регуляции гликолиза Вовлечение глюкозных остатков в процесс гликолиза обеспечивает важная реакция и эта реакция контролируется регуляторным
- 172. Регуляции гликолиза В печени преобладает фермент – глюкокиназа, которая не ингибируется глюкозо-6-фосфатом. Поэтому в печени, способной
- 173. Регуляции гликолиза
- 174. Гормональная регуляция гликолиза Глюкагон и инсулин влияют на синтез ключевых ферментов, используя системы трансмембранной передачи сигналов,
- 175. Регуляции пентозофосфатного пути превращения глюкозы
- 176. Регуляции глюконеогенеза
- 177. Гормональная регуляция глюконеогенеза В постабсорбтивный период глюкагон повышает транскрипцию генов и синтез ключевых ферментов глюконеогенеза -
- 178. Регуляция синтеза гликогена (гликогеногенез)
- 179. Регуляция мобилизации гликогена (гликогенолиз)
- 181. Регуляция общего пути катаболизма
- 183. Механизм действия инсулина – гипогликемического гормона Повышает проницаемость клеточных мембран для глюкозы, способствуя переходу ее из
- 184. Механизм действия инсулина Подавляет синтез ферментов глюконеогенеза, препятствует избыточному катаболизму жиров и белков и переходу их
- 185. Инсулин активирует: Ферменты гликолиза: гексокиназу, фосфофруктокиназу, пируваткиназу. Ферменты пентозофосфатного пути: глюкозо-6-фосфатдегидрогеназу, 6-фосфоглюконатдегидрогеназу. Ферменты гликогенеза (синтез гликогена):
- 186. Механизм действия адреналина и глюкагона Усиливают распад гликогена в мышцах и печени, активируя фосфорилазу гликогена и
- 187. Механизм действия глюкокортикоидов Усиливают глюконеогенез за счет индукции синтеза в клетках печени ключевых ферментов глюконеогенеза –
- 188. Гипергликемические гормоны: Адреналин, глюкогон – активация фосфорилазы. Кортикостероиды – активация (усиление синтеза) ферментов глюконеогенеза: пируваткарбоксилаза, ФЕП-карбоксилаза,
- 189. Регуляция скоростей синтеза и распада гликогена в печени поддерживает постоянство концентрации глюкозы в крови (3,33-5,55 ммоль/л).
- 190. Основным показателем состояния углеводного обмена является содержание глюкозы в крови. Нормальный уровень глюкозы в крови, взятой
- 191. Патология обмена углеводов гипергликемии физиологические патологические
- 192. Патология обмена углеводов К физиологическим гипергликемиям относятся: алиментарные, возникающие при одномоментном приеме больших количеств углеводов нейтрогенные,
- 193. Патология обмена углеводов Патологические гипергликемии обусловлены нарушением оптимального соотношения между секрецией гормонов гипо- и гипергликемического действия:
- 194. Клинические рекомендации Сахарный диабет 1 типа у взрослых, 2019 https://cr.minzdrav.gov.ru/recomend/286_2 Клинические рекомендации Сахарный диабет 2 типа
- 195. В основе сахарного диабета лежат следующие молекулярные дефекты: Нарушение превращения проинсулина в инсулин в результате мутаций,
- 196. В основе сахарного диабета лежат следующие молекулярные дефекты: Дефект рецепторов инсулина. У ряда больных секретируется нормальный
- 197. Важнейшими биохимическими признаками сахарного диабета являются: Гипергликемия. В результате недостатка инсулина нарушается проникновение глюкозы в ткани
- 198. ДИАГНОСТИЧЕСКИЕ КРИТЕРИИ САХАРНОГО ДИАБЕТА И ДРУГИХ НАРУШЕНИЙ ГЛИКЕМИИ (ВОЗ, 1999–2013) !!!!! Клинические рекомендации Сахарный диабет 2
- 199. ДИАГНОСТИЧЕСКИЕ КРИТЕРИИ САХАРНОГО ДИАБЕТА И ДРУГИХ НАРУШЕНИЙ ГЛИКЕМИИ (ВОЗ, 1999–2013) !!!!! Клинические рекомендации Сахарный диабет 2
- 200. Время определения концентрация глюкозы !!!!! Перевод глюкозы крови из ммоль/л в мг/дл: 1 ммоль/л × 18,02
- 201. ПГТТ – пероральный глюкозотолерантный тест. Проводится в случае сомнительных значений гликемии для уточнения диагноза. !!!!! Правила
- 202. ОСТРЫЕ ОСЛОЖНЕНИЯ САХАРНОГО ДИАБЕТА Диабетический кетоацидоз (дка, диабетическая кетоацидотическая кома) Гиперосмолярное гипергликемическое состояние (ггс) Молочнокислый ацидоз
- 203. 1. Диабетический кетоацидоз (дка, диабетическая кетоацидотическая кома) ! ДКА – требующая экстренной госпитализации острая декомпенсация СД,
- 204. Лабораторные изменения: диагностика и дифференциальная диагностика ДКА Общий анализ мочи: Глюкозурия, кетонурия, протеинурия (непостоянно). Биохимический анализ
- 205. 2. ГИПЕРОСМОЛЯРНОЕ ГИПЕРГЛИКЕМИЧЕСКОЕ СОСТОЯНИЕ (ГГС)! ГГС – острая декомпенсация СД, с резко выраженной гипергликемией (как правило,
- 206. Лабораторные изменения: диагностика и дифференциальная диагностика ГГС Общий анализ мочи: Общий анализ мочи: Массивная глюкозурия, протеинурия
- 207. 3. МОЛОЧНОКИСЛЫЙ АЦИДОЗ (ЛАКТАТАЦИДОЗ) ! Лактатацидоз – метаболический ацидоз с большой анионной разницей (≥10 ммоль/л) и
- 208. Лабораторные изменения: диагностика и дифференциальная диагностика лактатацидоза Биохимический анализ крови: Лактат > 4,0 ммоль/л, реже 2,2
- 209. 4. ГИПОГЛИКЕМИЯ И ГИПОГЛИКЕМИЧЕСКАЯ КОМА ! Гипогликемия – уровень глюкозы плазмы Основная причина – повышенное образование
- 210. Лабораторные изменения: диагностика и дифференциальная диагностика гипогликемии и гипогликемической комы Биохимический анализ крови: Глюкоза плазмы
- 211. Гликогеновые болезни - это группа наследственных заболеваний, причинами которых являются дефекты ферментов, участвующих в синтезе или
- 212. Гликогенозы (обмен гликогена) Глю Глю-6ф Глю-1ф УДФ-глю Гликоген(n+1) Гликогенсинтаза (1-4) «Ветвящий» фермент (1-6) Гликогенn УДФ Глю-6-фосфатаза
- 213. Гликогенозы Всего известно 12 типов гликогенозов. По патогенетическому признаку гликогенозы делят: печеночные – 0, I, III,
- 214. Печеночные гликогенозы
- 215. Печеночные гликогенозы
- 216. Мышечные гликогенозы
- 217. Смешанные гликогенозы
- 219. Скачать презентацию

План лекции:
Общие закономерности метаболизма.
Этапы катаболизма.
Цепь переноса электронов (ЦПЭ), состав, строение компонентов
План лекции:
Общие закономерности метаболизма.
Этапы катаболизма.
Цепь переноса электронов (ЦПЭ), состав, строение компонентов
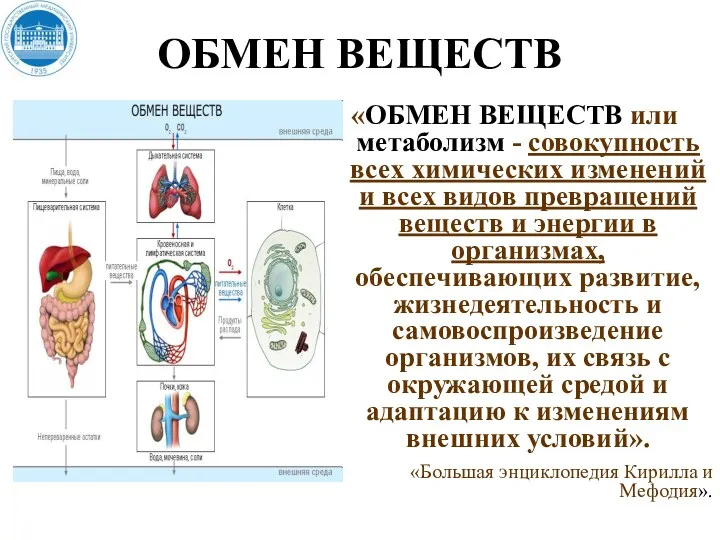
ОБМЕН ВЕЩЕСТВ
«ОБМЕН ВЕЩЕСТВ или метаболизм - совокупность всех химических изменений и
ОБМЕН ВЕЩЕСТВ
«ОБМЕН ВЕЩЕСТВ или метаболизм - совокупность всех химических изменений и

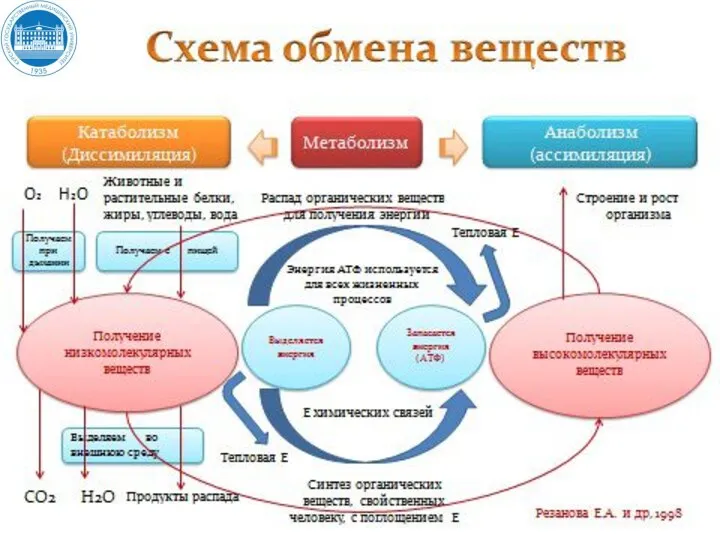
Промежуточный обмен
(внутриклеточный метаболизм)
включает два типа реакций:
Катаболизм – биосинтетические
Промежуточный обмен
(внутриклеточный метаболизм)
включает два типа реакций:
Катаболизм – биосинтетические

Промежуточный обмен
(внутриклеточный метаболизм)
включает два типа реакций:
Анаболизм - биосинтетические процессы,
Промежуточный обмен
(внутриклеточный метаболизм)
включает два типа реакций:
Анаболизм - биосинтетические процессы,

Процессы окисления протекают в организме и вне организма. Эти процессы имеют
Процессы окисления протекают в организме и вне организма. Эти процессы имеют

Сходство между окислением в организме и вне организма.
В результате окисления образуются
Сходство между окислением в организме и вне организма.
В результате окисления образуются

Различия между окислением в организме и вне организма.
Вне организма энергия выделяется
Различия между окислением в организме и вне организма.
Вне организма энергия выделяется

Различия между окислением в организме и вне организма.
Вне организма энергия выделяется
Различия между окислением в организме и вне организма.
Вне организма энергия выделяется

Различия между окислением в организме и вне организма.
Основной реакцией окисления в
Различия между окислением в организме и вне организма.
Основной реакцией окисления в

Утилизация энергии в виде макроэргических связей
При разрыве макроэргической связи образуется более
Утилизация энергии в виде макроэргических связей
При разрыве макроэргической связи образуется более
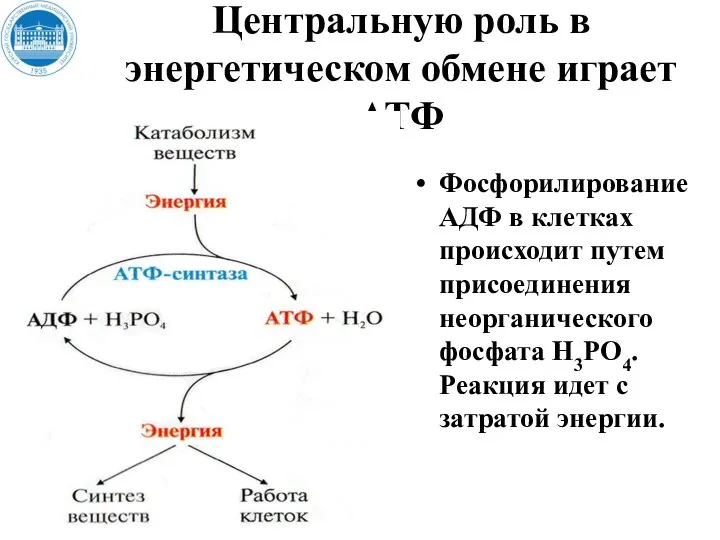
Центральную роль в энергетическом обмене играет АТФ
Фосфорилирование АДФ в клетках происходит
Центральную роль в энергетическом обмене играет АТФ
Фосфорилирование АДФ в клетках происходит
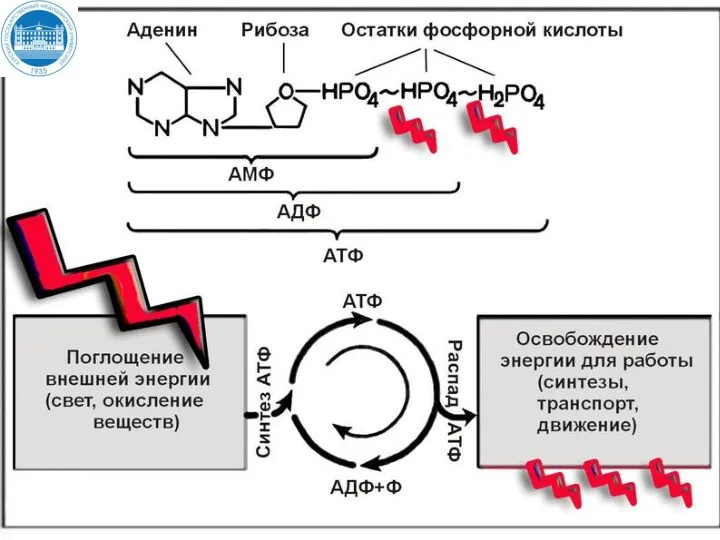
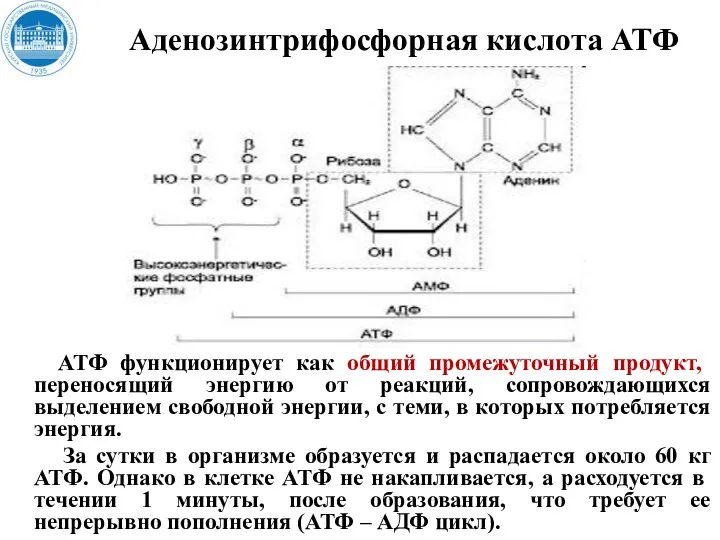
Аденозинтрифосфорная кислота АТФ
АТФ функционирует как общий промежуточный продукт, переносящий энергию
Аденозинтрифосфорная кислота АТФ
АТФ функционирует как общий промежуточный продукт, переносящий энергию

Типы фосфорилирования АДФ
В зависимости от источника энергии, обеспечивающего присоединение фосфатного
Типы фосфорилирования АДФ
В зависимости от источника энергии, обеспечивающего присоединение фосфатного

Субстратное фосфорилирование -
это процесс синтез АТФ из АДФ и неорганического фосфата
Субстратное фосфорилирование -
это процесс синтез АТФ из АДФ и неорганического фосфата

Окислительное фосфорилирование АДФ - синтез АТФ из АДФ и Н3РО4 за
Окислительное фосфорилирование АДФ - синтез АТФ из АДФ и Н3РО4 за
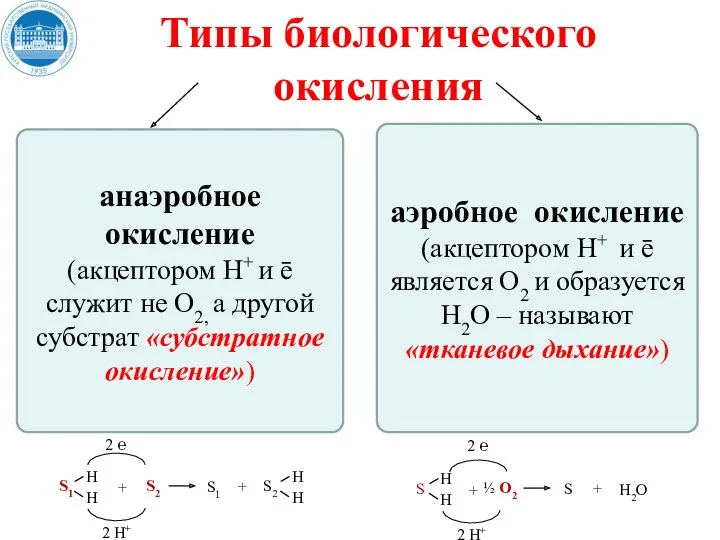
Типы биологического окисления
анаэробное окисление (акцептором Н+ и ē служит не О2,
Типы биологического окисления
анаэробное окисление (акцептором Н+ и ē служит не О2,

Тканевое дыхание
Тканевым дыханием являются реакции окисления органических веществ, в которых используется
Тканевое дыхание
Тканевым дыханием являются реакции окисления органических веществ, в которых используется
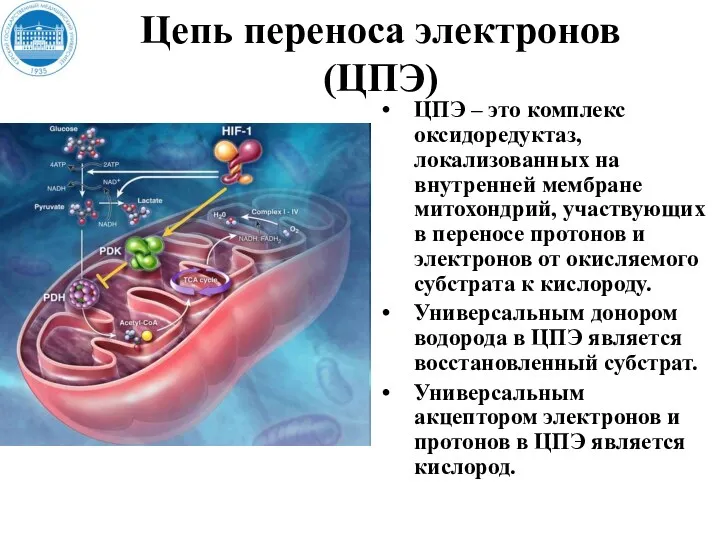
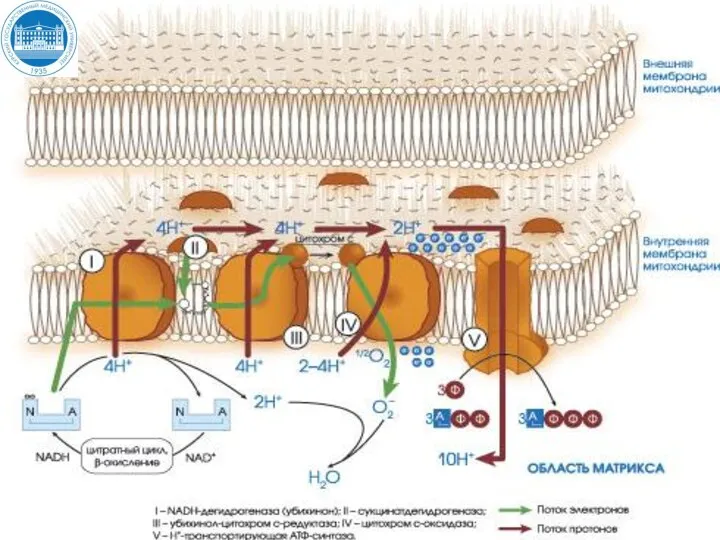
Цепь переноса электронов (ЦПЭ)
ЦПЭ – это комплекс оксидоредуктаз, локализованных на внутренней
Цепь переноса электронов (ЦПЭ)
ЦПЭ – это комплекс оксидоредуктаз, локализованных на внутренней

Первичные акцепторы водорода
Никотинамидзависимые дегидрогеназы (NAD, NADР)
(находятся в матриксе митохондрий
Первичные акцепторы водорода Никотинамидзависимые дегидрогеназы (NAD, NADР) (находятся в матриксе митохондрий

Первичные акцепторы водорода
Флавинзависимые
дегидрогеназы (FAD, FMN)
В отличие от NAD FAD соединен
Первичные акцепторы водорода
Флавинзависимые
дегидрогеназы (FAD, FMN)
В отличие от NAD FAD соединен
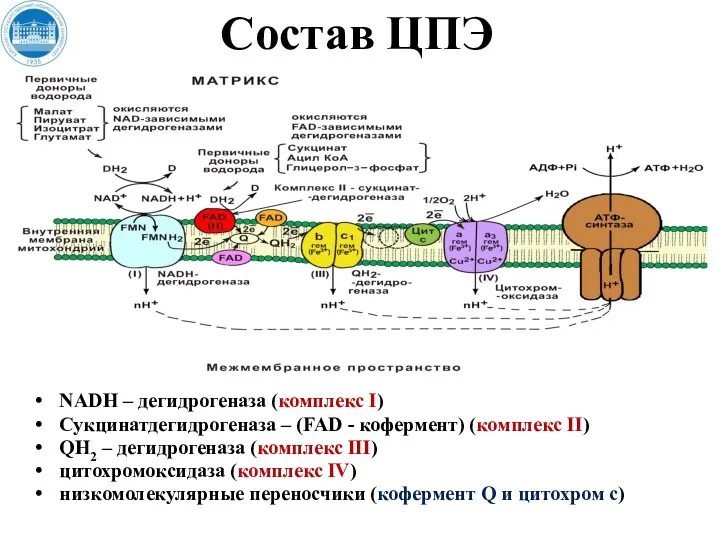
Состав ЦПЭ
NADH – дегидрогеназа (комплекс I)
Сукцинатдегидрогеназа – (FAD - кофермент) (комплекс
Состав ЦПЭ
NADH – дегидрогеназа (комплекс I)
Сукцинатдегидрогеназа – (FAD - кофермент) (комплекс

NADH – дегидрогеназа (комплекс I)
Комплекс I ЦПЭ содержит ФМН и 5
NADH – дегидрогеназа (комплекс I)
Комплекс I ЦПЭ содержит ФМН и 5

Сукцинатдегидрогеназа –
(FAD - кофермент) (комплекс II)
Сукцинатдегидрогеназа - СДГ (FAD –
Сукцинатдегидрогеназа –
(FAD - кофермент) (комплекс II)
Сукцинатдегидрогеназа - СДГ (FAD –
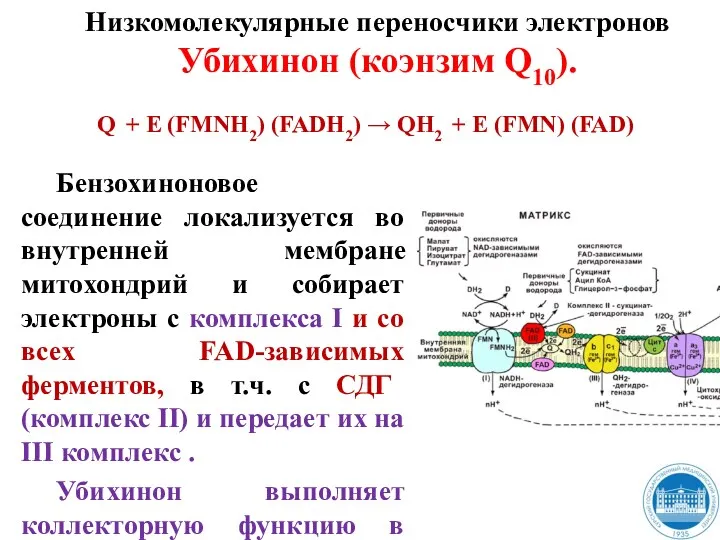
Бензохиноновое соединение локализуется во внутренней мембране митохондрий и собирает электроны с
Бензохиноновое соединение локализуется во внутренней мембране митохондрий и собирает электроны с

QH2 (коэнзим Q –дегидрогеназа)
(комплекс III)
Состоит из 2-типов цитохромов (b и
QH2 (коэнзим Q –дегидрогеназа)
(комплекс III)
Состоит из 2-типов цитохромов (b и
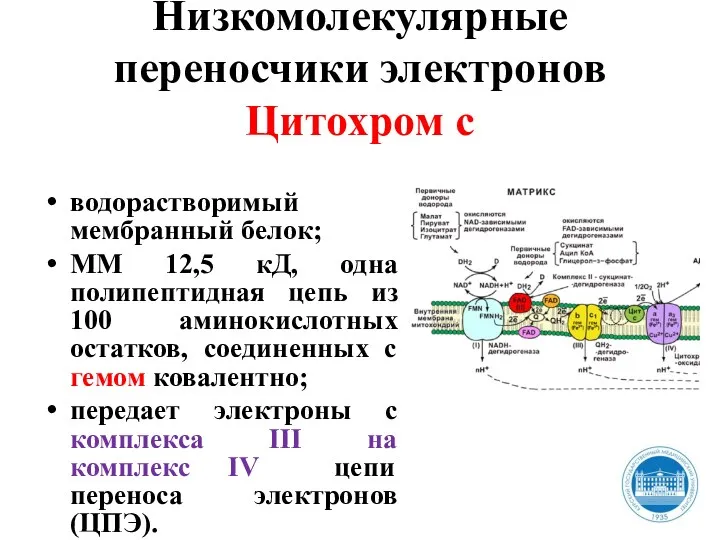
Низкомолекулярные переносчики электронов
Цитохром с
водорастворимый мембранный белок;
ММ 12,5 кД, одна
Низкомолекулярные переносчики электронов
Цитохром с
водорастворимый мембранный белок;
ММ 12,5 кД, одна

Цитохром-c-оксидаза (цит а/а3)
(комплекс IV)
Комплекс IV ЦПЭ содержит цитохромы а, а3,
Цитохром-c-оксидаза (цит а/а3)
(комплекс IV)
Комплекс IV ЦПЭ содержит цитохромы а, а3,
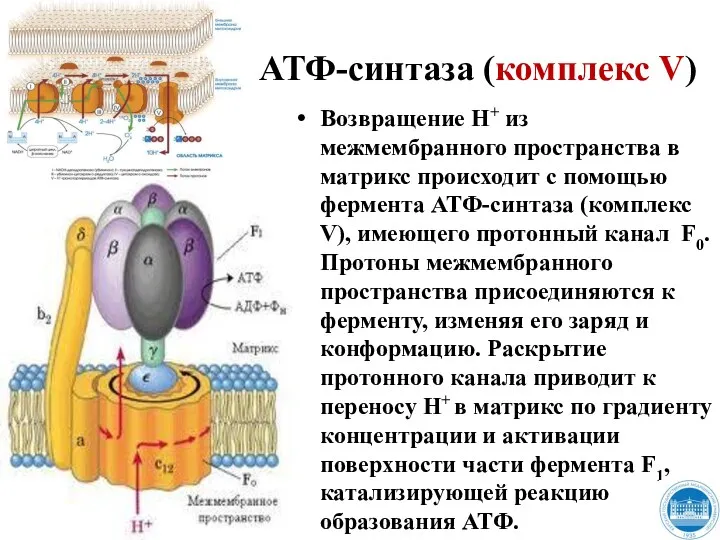
АТФ-синтаза (комплекс V)
Возвращение Н+ из межмембранного пространства в матрикс происходит с
АТФ-синтаза (комплекс V)
Возвращение Н+ из межмембранного пространства в матрикс происходит с
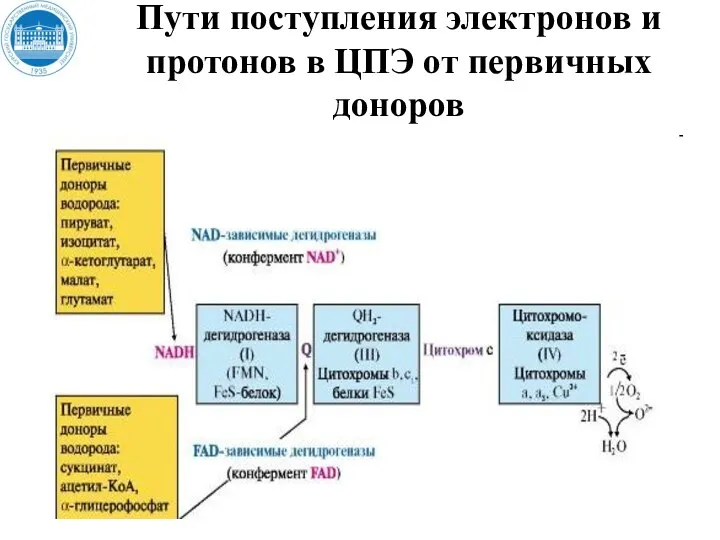
(II)
Пути поступления электронов и протонов в ЦПЭ от первичных доноров
(II)
Пути поступления электронов и протонов в ЦПЭ от первичных доноров
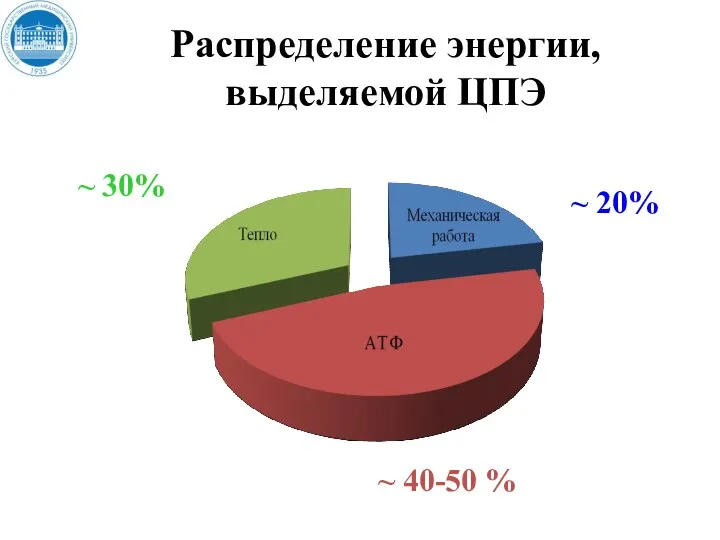
Распределение энергии, выделяемой ЦПЭ
~ 20%
~ 40-50 %
~ 30%
Распределение энергии, выделяемой ЦПЭ
~ 20%
~ 40-50 %
~ 30%
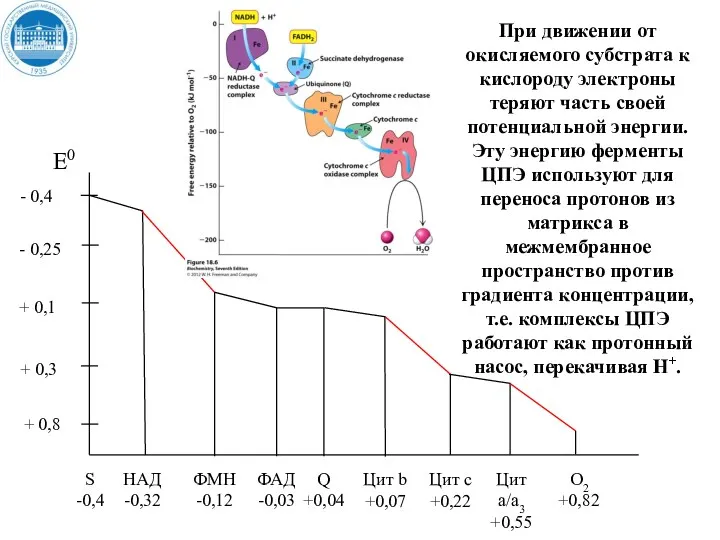
+ 0,8
При движении от окисляемого субстрата к кислороду электроны теряют часть
+ 0,8
При движении от окисляемого субстрата к кислороду электроны теряют часть
+ 0,8
В трех пунктах переноса электронов происходит наибольшее изменение свободной энергии
+ 0,8
В трех пунктах переноса электронов происходит наибольшее изменение свободной энергии

Пункты сопряжения постоянны, но их количество зависит от природы окисляемого субстрата.
При
Пункты сопряжения постоянны, но их количество зависит от природы окисляемого субстрата.
При

Коэффициент окислительного фосфорилирование Р/О показывает, какое количество неорганического фосфата затрачивается для
Коэффициент окислительного фосфорилирование Р/О показывает, какое количество неорганического фосфата затрачивается для

Коэффициент окислительного фосфорилирования
НАД-Н+ + ½О2 →НАД+ + Н2О
3 АДФ + 3
Коэффициент окислительного фосфорилирования
НАД-Н+ + ½О2 →НАД+ + Н2О
3 АДФ + 3

Причины нарушения биологического окисления
Недостаток субстрата (голодание, пищева-рение, межуточное нарушение обменов белков,
Причины нарушения биологического окисления
Недостаток субстрата (голодание, пищева-рение, межуточное нарушение обменов белков,

Вещества, понижающие интенсивность окислительного фосфолирирования АДФ.
Ингибиторы ферментов ЦПЭ подавляют активность ферментных
Вещества, понижающие интенсивность окислительного фосфолирирования АДФ.
Ингибиторы ферментов ЦПЭ подавляют активность ферментных

Примеры веществ, понижающие интенсивность окислительного фосфолирирования АДФ.
ингибиторы NADH-дегидрогиназы - лекарственные препараты
Примеры веществ, понижающие интенсивность окислительного фосфолирирования АДФ.
ингибиторы NADH-дегидрогиназы - лекарственные препараты

Вещество, способное полностью блокировать тканевое дыхание, синильная кислота
Синильная кислота является веществом,
Вещество, способное полностью блокировать тканевое дыхание, синильная кислота
Синильная кислота является веществом,

Вещество, способное полностью блокировать тканевое дыхание, синильная кислота
Соединившись с цианидом, цитохромоксидаза
Вещество, способное полностью блокировать тканевое дыхание, синильная кислота
Соединившись с цианидом, цитохромоксидаза

Вещества, понижающие интенсивность окислительного фосфолирирования АДФ.
Ингибиторы АТФ-синтазы снижают активность фермента, скорость
Вещества, понижающие интенсивность окислительного фосфолирирования АДФ.
Ингибиторы АТФ-синтазы снижают активность фермента, скорость

Вещества, понижающие интенсивность окислительного фосфолирирования АДФ.
Разобщители окисления и фосфорилирования – липофильные
Вещества, понижающие интенсивность окислительного фосфолирирования АДФ.
Разобщители окисления и фосфорилирования – липофильные

Лекция № 4
Обмен углеводов: значение, переваривание. Гликолиз. Пентозофосфатный путь окисления глюкозы.
Лекция № 4
Обмен углеводов: значение, переваривание. Гликолиз. Пентозофосфатный путь окисления глюкозы.

ПЛАН ЛЕКЦИИ:
Обмен углеводов.
Гликолиз.
Пентозофосфатный путь окисления глюкозы.
ПЛАН ЛЕКЦИИ:
Обмен углеводов.
Гликолиз.
Пентозофосфатный путь окисления глюкозы.

Обмен углеводов в организме человека складываются из следующих процессов:
Расщепление в желудочно-кишечном
Обмен углеводов в организме человека складываются из следующих процессов:
Расщепление в желудочно-кишечном
ОБМЕН УГЛЕВОДОВ
Распад (гликогеногенолиз)
и синтез (гликогеногенез)
гликогена
Аэробный гликолиз
Окислительное
декарбоксилирование
пирувата
ЦТК
Пентозо-
фосфатный
путь
Глюконеогенез
Анаэробный гликолиз
ОБМЕН УГЛЕВОДОВ
Распад (гликогеногенолиз)
и синтез (гликогеногенез)
гликогена
Аэробный гликолиз
Окислительное
декарбоксилирование
пирувата
ЦТК
Пентозо-
фосфатный
путь
Глюконеогенез
Анаэробный гликолиз

Две основные функции:
Углеводы – источник углеродов, который необходим для синтеза ряда
Две основные функции:
Углеводы – источник углеродов, который необходим для синтеза ряда

Другие функции:
Резервная (крахмал, гликоген).
Структурная (полисахариды образуют прочный остов в комплексе с
Другие функции:
Резервная (крахмал, гликоген).
Структурная (полисахариды образуют прочный остов в комплексе с

Переваривание
углеводов в организме
Источником углеводов для организма служат углеводы пищи -
Переваривание
углеводов в организме
Источником углеводов для организма служат углеводы пищи -

Переваривание углеводов
Амилаза слюны расщепляет α-1,4-гликозидные связи в крахмале. В ротовой полости
Переваривание углеводов
Амилаза слюны расщепляет α-1,4-гликозидные связи в крахмале. В ротовой полости

Переваривание углеводов
Последующее переваривание нерасщепленного или частично расщепленного крахмала происходит в кишечнике.
Переваривание углеводов
Последующее переваривание нерасщепленного или частично расщепленного крахмала происходит в кишечнике.

Переваривание углеводов
Мальтоза и изомальтоза вместе с другими пищевыми дисахаридами - сахарозой
Переваривание углеводов
Мальтоза и изомальтоза вместе с другими пищевыми дисахаридами - сахарозой
Переваривание углеводов
Переваривание углеводов

Продукты полного переваривания углеводов – глюкоза, галактоза и фруктоза –
Продукты полного переваривания углеводов – глюкоза, галактоза и фруктоза –

Гликолиз
Гликолиз (от греч. glykys – сладкий, lysys – распад )
Гликолиз
Гликолиз (от греч. glykys – сладкий, lysys – распад )

Гликолиз включает превращения трех разных типов:
Распад углеродного скелета глюкозы с
Гликолиз включает превращения трех разных типов:
Распад углеродного скелета глюкозы с
АЭРОБНЫЙ ГЛИКОЛИЗ - специфический путь катаболизма глюкозы, в результате которого происходит
АЭРОБНЫЙ ГЛИКОЛИЗ - специфический путь катаболизма глюкозы, в результате которого происходит
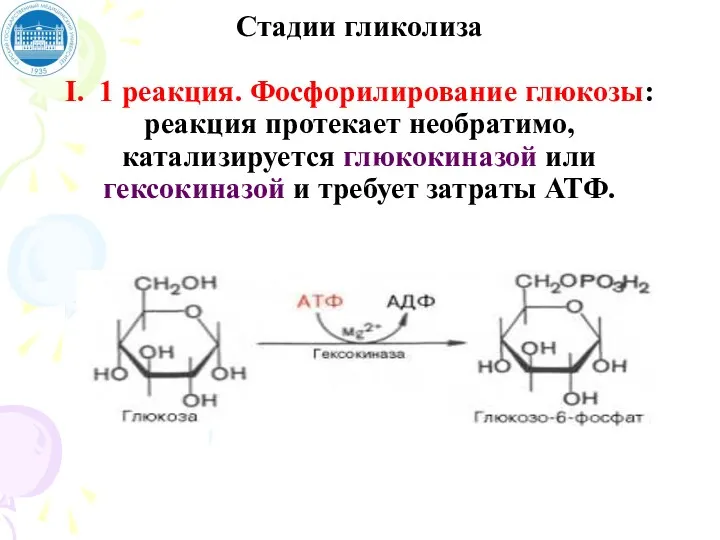
Стадии гликолиза
I. 1 реакция. Фосфорилирование глюкозы: реакция протекает необратимо, катализируется глюкокиназой
Стадии гликолиза I. 1 реакция. Фосфорилирование глюкозы: реакция протекает необратимо, катализируется глюкокиназой

Ферменты участвующие в фосфорилировании глюкозы.
Связывание гексокиназы с гексозой происходит по типу
Ферменты участвующие в фосфорилировании глюкозы.
Связывание гексокиназы с гексозой происходит по типу
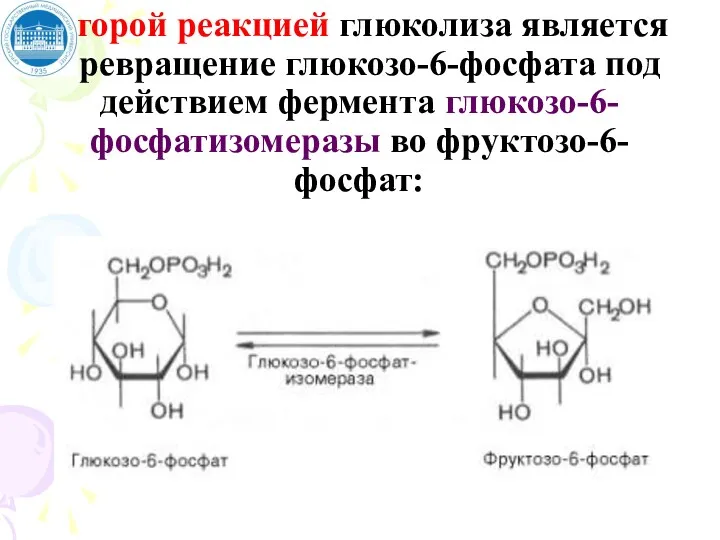
Второй реакцией глюколиза является превращение глюкозо-6-фосфата под действием фермента глюкозо-6-фосфатизомеразы во фруктозо-6-фосфат:
Второй реакцией глюколиза является превращение глюкозо-6-фосфата под действием фермента глюкозо-6-фосфатизомеразы во фруктозо-6-фосфат:
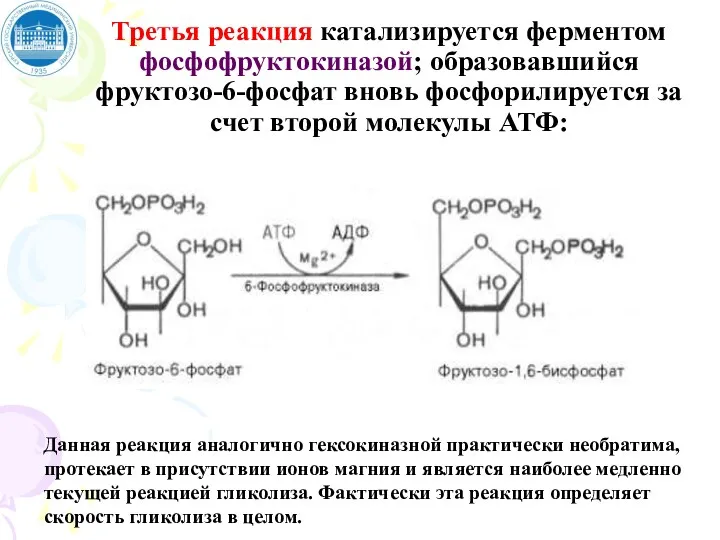
Третья реакция катализируется ферментом фосфофруктокиназой; образовавшийся фруктозо-6-фосфат вновь фосфорилируется за счет второй молекулы АТФ:
Данная реакция
Третья реакция катализируется ферментом фосфофруктокиназой; образовавшийся фруктозо-6-фосфат вновь фосфорилируется за счет второй молекулы АТФ:
Данная реакция
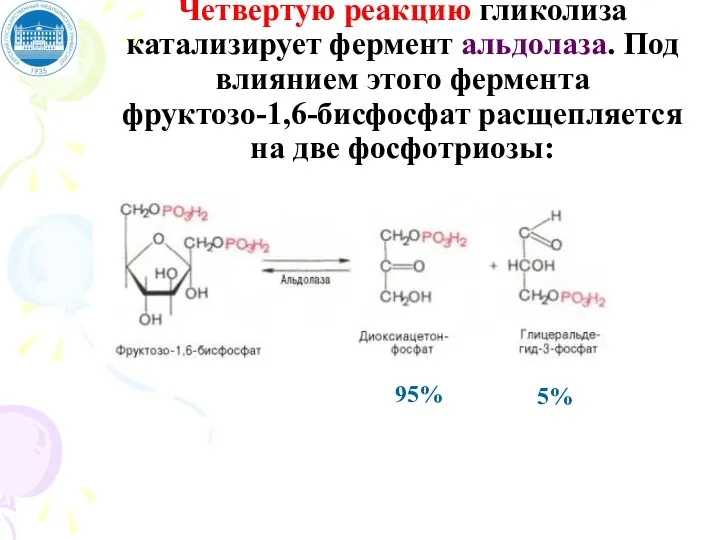
Четвертую реакцию гликолиза катализирует фермент альдолаза. Под влиянием этого фермента фруктозо-1,6-бисфосфат
Четвертую реакцию гликолиза катализирует фермент альдолаза. Под влиянием этого фермента фруктозо-1,6-бисфосфат
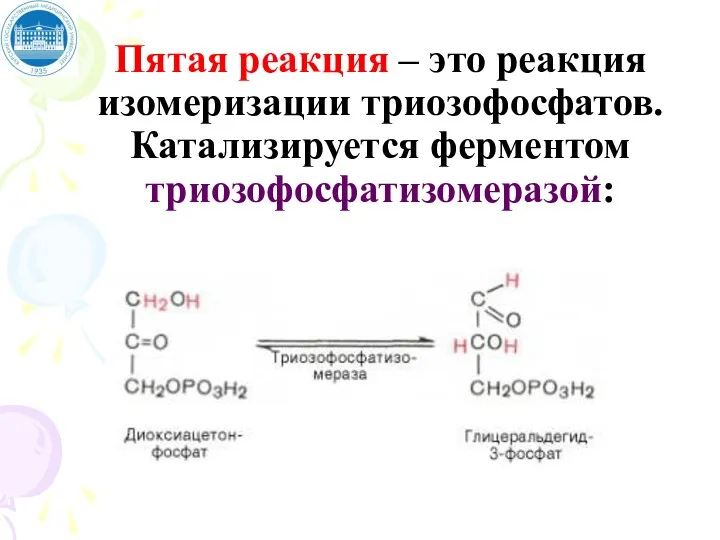
Пятая реакция – это реакция изомеризации триозофосфатов. Катализируется ферментом триозофосфатизомеразой:
Пятая реакция – это реакция изомеризации триозофосфатов. Катализируется ферментом триозофосфатизомеразой:
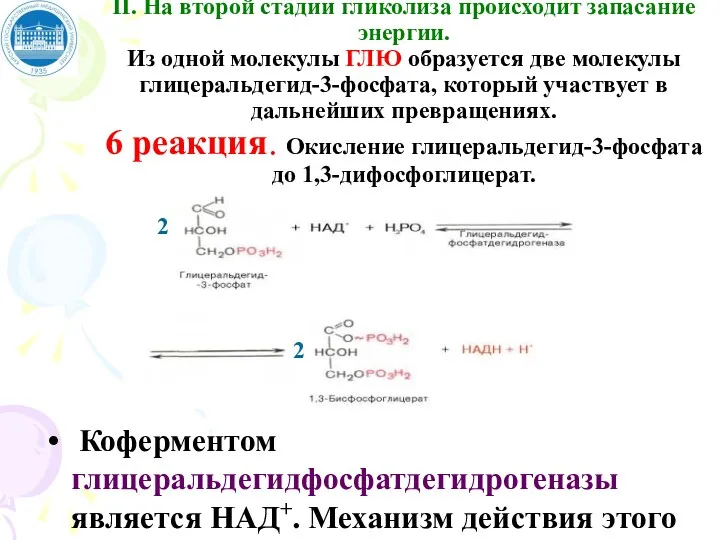
II. На второй стадии гликолиза происходит запасание энергии.
Из одной молекулы ГЛЮ
II. На второй стадии гликолиза происходит запасание энергии. Из одной молекулы ГЛЮ
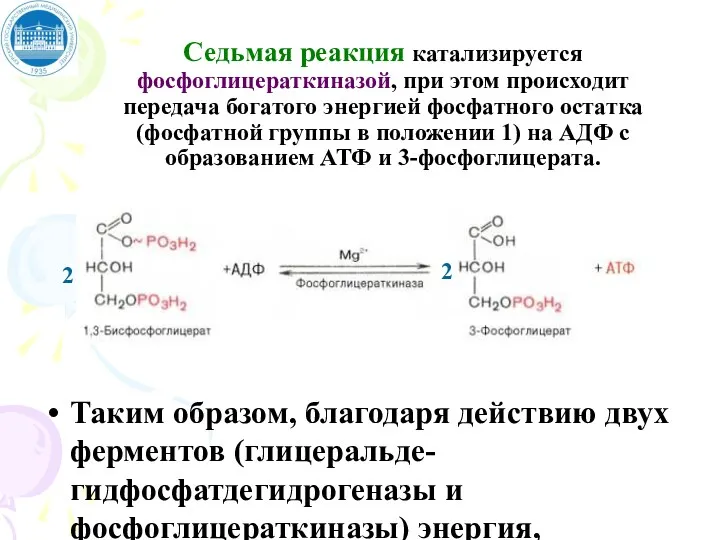
Седьмая реакция катализируется фосфоглицераткиназой, при этом происходит передача богатого энергией фосфатного
Седьмая реакция катализируется фосфоглицераткиназой, при этом происходит передача богатого энергией фосфатного

Восьмая реакция сопровождается внутримолекулярным переносом оставшейся фосфатной группы, и
3-фосфоглицерат превращается
Восьмая реакция сопровождается внутримолекулярным переносом оставшейся фосфатной группы, и 3-фосфоглицерат превращается
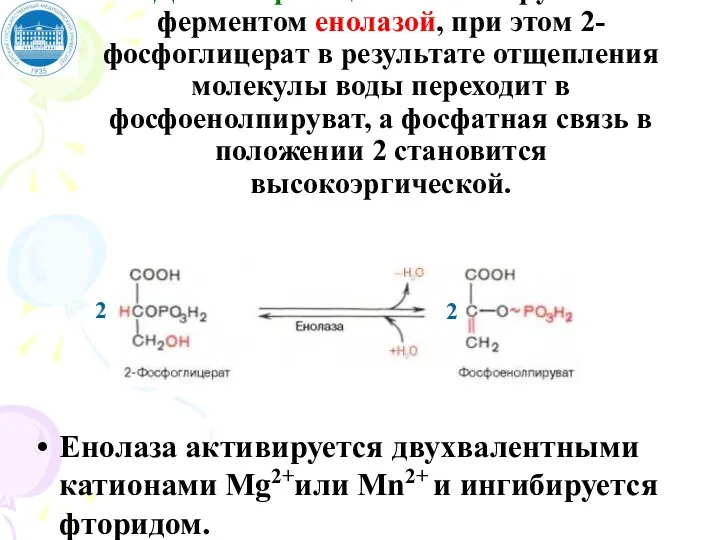
Девятая реакция катализируется ферментом енолазой, при этом 2-фосфоглицерат в результате отщепления
Девятая реакция катализируется ферментом енолазой, при этом 2-фосфоглицерат в результате отщепления

Десятая реакция характеризуется разрывом высокоэргической связи и переносом фосфатного остатка от
Десятая реакция характеризуется разрывом высокоэргической связи и переносом фосфатного остатка от

Конечным продуктом аэробного гликолиза является
пируват, а энергетический баланс складывается
Конечным продуктом аэробного гликолиза является
пируват, а энергетический баланс складывается
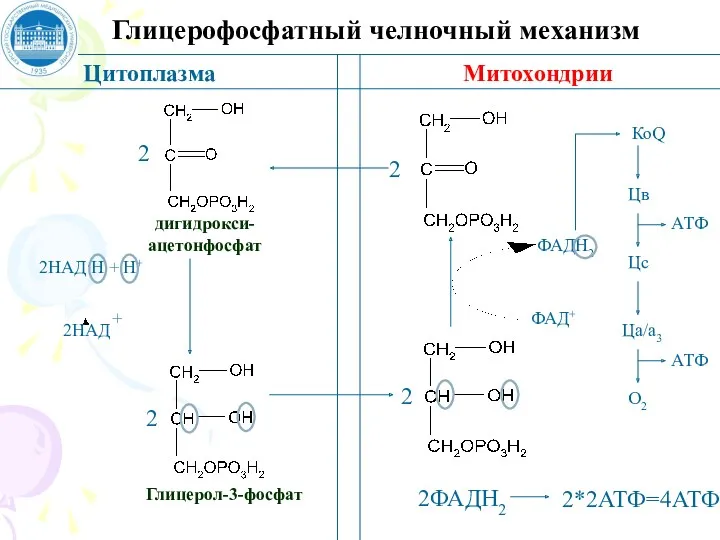
Глицерофосфатный челночный механизм
Цитоплазма
Митохондрии
дигидрокси-
ацетонфосфат
Глицерол-3-фосфат
2
2
2НАД·Н + Н+
2НАД
+
ФАД+
ФАДН2
2
2
КоQ
Цв
Цс
Ца/а3
O2
АТФ
АТФ
2ФАДН2
2*2АТФ=4АТФ
Глицерофосфатный челночный механизм
Цитоплазма
Митохондрии
дигидрокси-
ацетонфосфат
Глицерол-3-фосфат
2
2
2НАД·Н + Н+
2НАД
+
ФАД+
ФАДН2
2
2
КоQ
Цв
Цс
Ца/а3
O2
АТФ
АТФ
2ФАДН2
2*2АТФ=4АТФ
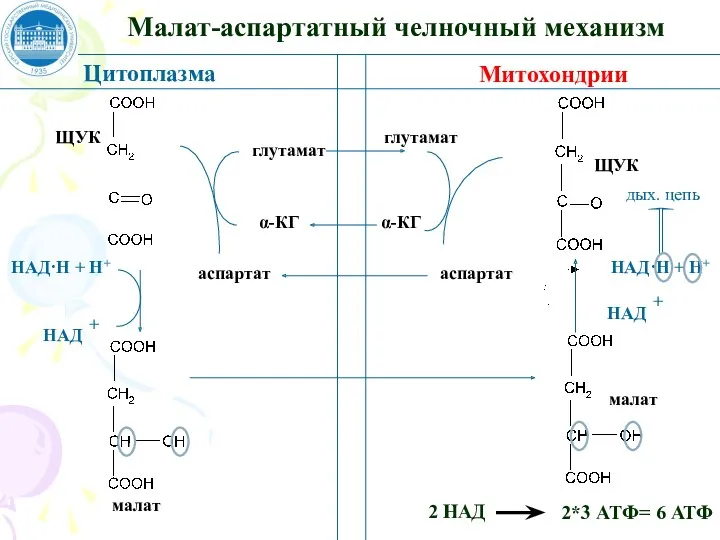
Малат-аспартатный челночный механизм
Цитоплазма
Митохондрии
ЩУК
малат
НАД·Н + Н+
НАД
+
глутамат
аспартат
глутамат
аспартат
ЩУК
НАД·Н + Н+
НАД
+
дых. цепь
малат
α-КГ
α-КГ
2 НАД
2*3
Малат-аспартатный челночный механизм
Цитоплазма
Митохондрии
ЩУК
малат
НАД·Н + Н+
НАД
+
глутамат
аспартат
глутамат
аспартат
ЩУК
НАД·Н + Н+
НАД
+
дых. цепь
малат
α-КГ
α-КГ
2 НАД
2*3

Баланс аэробного гликолиза
1 реакция - 1 АТФ
3 реакция - 1 АТФ
6
Баланс аэробного гликолиза
1 реакция - 1 АТФ
3 реакция - 1 АТФ
6
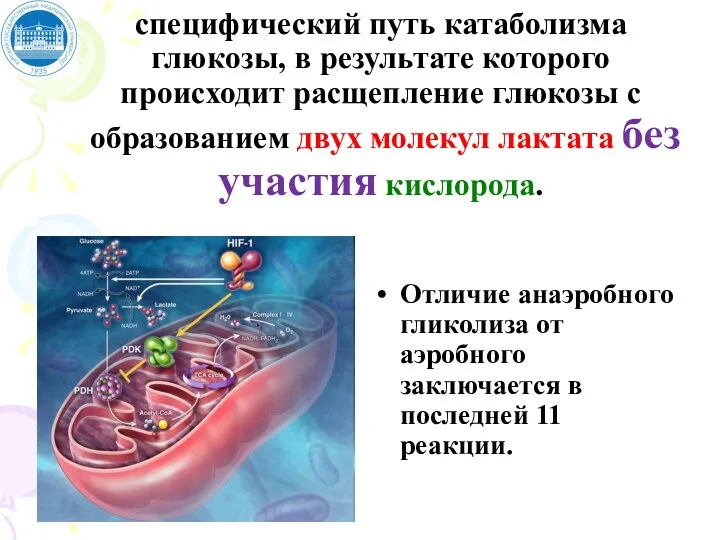
АНАЭРОБНЫЙ ГЛИКОЛИЗ - специфический путь катаболизма глюкозы, в результате которого происходит
АНАЭРОБНЫЙ ГЛИКОЛИЗ - специфический путь катаболизма глюкозы, в результате которого происходит

Дальше процесс идет в зависимости от наличия или отсутствия кислорода
Дальше процесс идет в зависимости от наличия или отсутствия кислорода

В этих условиях образовавшийся при гликолизе НАДН регенерируется за счет
В этих условиях образовавшийся при гликолизе НАДН регенерируется за счет

Баланс анаэробного гликолиза
1 реакция - 1 АТФ
3 реакция - 1 АТФ
6
Баланс анаэробного гликолиза
1 реакция - 1 АТФ
3 реакция - 1 АТФ
6

Пентозофосфатный путь окисления глюкозы
Локализация: молочная железа, эмбриональная и жировая ткань,
Пентозофосфатный путь окисления глюкозы
Локализация: молочная железа, эмбриональная и жировая ткань,

В пентозофосфатном пути превращения глюкозы можно выделить:
Окислительный этап
Поставляет клеткам кофермент NADPH
В пентозофосфатном пути превращения глюкозы можно выделить:
Окислительный этап
Поставляет клеткам кофермент NADPH
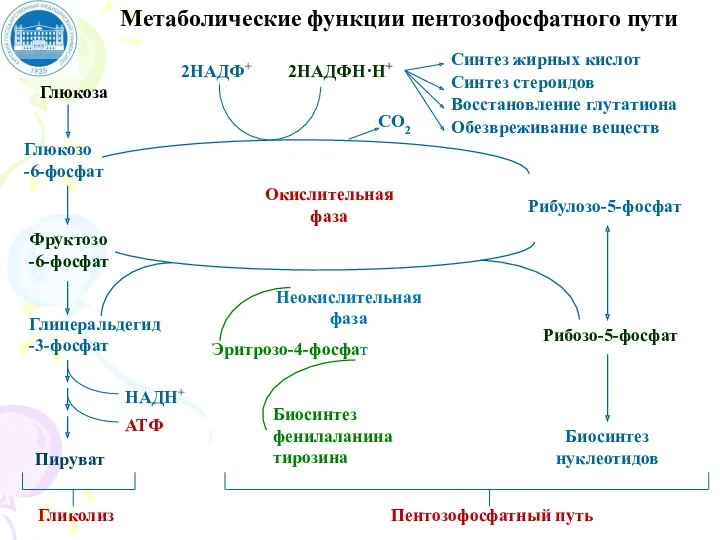
Глюкоза
Глюкозо
-6-фосфат
Фруктозо
-6-фосфат
Глицеральдегид-3-фосфат
Гликолиз
2НАДФ+
2НАДФН·Н+
Окислительная фаза
Синтез жирных кислот
Синтез стероидов
Восстановление глутатиона
Неокислительная фаза
Биосинтез
нуклеотидов
Рибулозо-5-фосфат
Рибозо-5-фосфат
Метаболические функции пентозофосфатного пути
СО2
Обезвреживание веществ
Пируват
НАДН+
АТФ
Пентозофосфатный
Глюкоза
Глюкозо
-6-фосфат
Фруктозо
-6-фосфат
Глицеральдегид-3-фосфат
Гликолиз
2НАДФ+
2НАДФН·Н+
Окислительная фаза
Синтез жирных кислот
Синтез стероидов
Восстановление глутатиона
Неокислительная фаза
Биосинтез
нуклеотидов
Рибулозо-5-фосфат
Рибозо-5-фосфат
Метаболические функции пентозофосфатного пути
СО2
Обезвреживание веществ
Пируват
НАДН+
АТФ
Пентозофосфатный

Условия протекания пентозофосфатного пути окисления глюкозы
Судьба глюкозо-6-фосфата — вступит ли он
Условия протекания пентозофосфатного пути окисления глюкозы
Судьба глюкозо-6-фосфата — вступит ли он
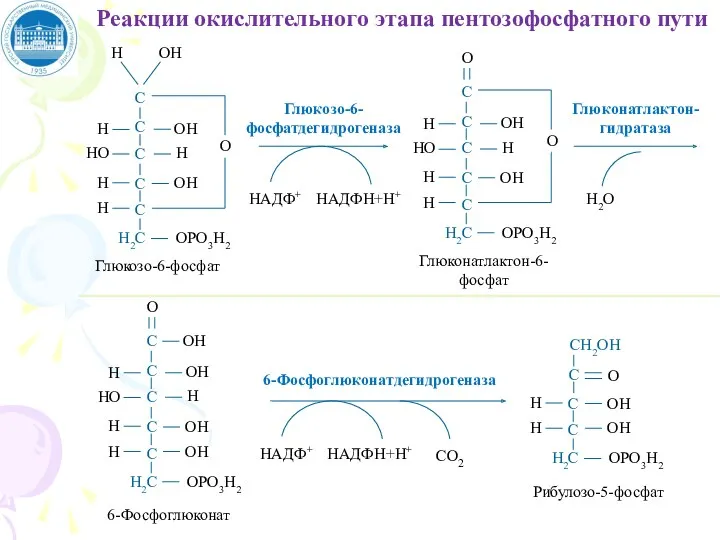
НАДФ+
НАДФН+Н+
Глюкозо-6-фосфатдегидрогеназа
Глюконатлактон-гидратаза
Н2О
НАДФ+
НАДФН+Н+
6-Фосфоглюконатдегидрогеназа
СО2
Реакции окислительного этапа пентозофосфатного пути
НАДФ+
НАДФН+Н+
Глюкозо-6-фосфатдегидрогеназа
Глюконатлактон-гидратаза
Н2О
НАДФ+
НАДФН+Н+
6-Фосфоглюконатдегидрогеназа
СО2
Реакции окислительного этапа пентозофосфатного пути

Патология пентозофосфатного пути окисления глюкозы
Генетический дефект фермента глюкозо-6-фосфатдегидрогеназы является причиной:
Патология пентозофосфатного пути окисления глюкозы
Генетический дефект фермента глюкозо-6-фосфатдегидрогеназы является причиной:

Лекция № 5
Специфические пути катаболизма.
Общий путь катаболизма: окислительное декарбоксилирование пирувата
Лекция № 5
Специфические пути катаболизма.
Общий путь катаболизма: окислительное декарбоксилирование пирувата

Специфические и общий пути катаболизма
Специфические и общий пути катаболизма
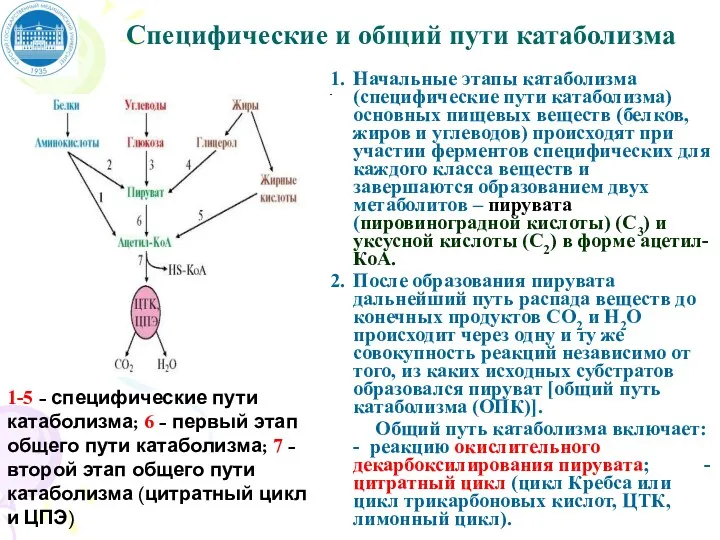
Специфические и общий пути катаболизма
Начальные этапы катаболизма (специфические пути катаболизма) основных
Специфические и общий пути катаболизма
Начальные этапы катаболизма (специфические пути катаболизма) основных

Суммарное уравнение
реакции окислительного декарбоксилирования пирувата:
Эту реакцию катализирует сложно организованный пируватдегидрогеназный
Суммарное уравнение
реакции окислительного декарбоксилирования пирувата:
Эту реакцию катализирует сложно организованный пируватдегидрогеназный
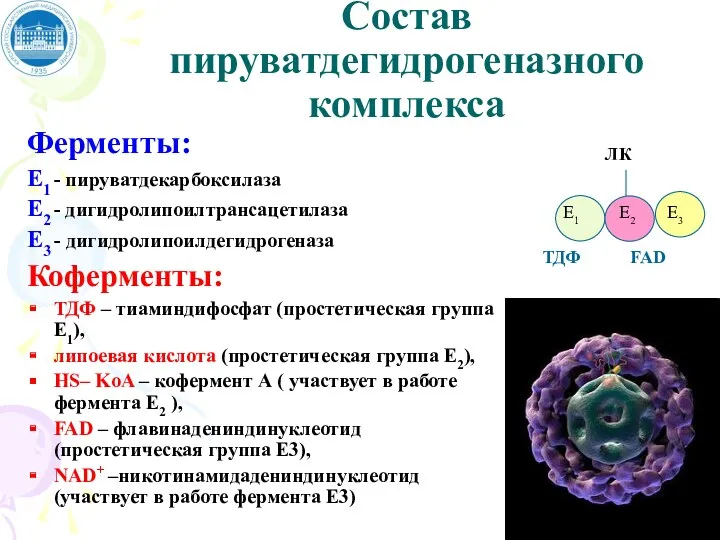
Состав
пируватдегидрогеназного комплекса
Ферменты:
Е1 - пируватдекарбоксилаза
Е2 - дигидролипоилтрансацетилаза
Е3 -
Состав
пируватдегидрогеназного комплекса
Ферменты:
Е1 - пируватдекарбоксилаза
Е2 - дигидролипоилтрансацетилаза
Е3 -
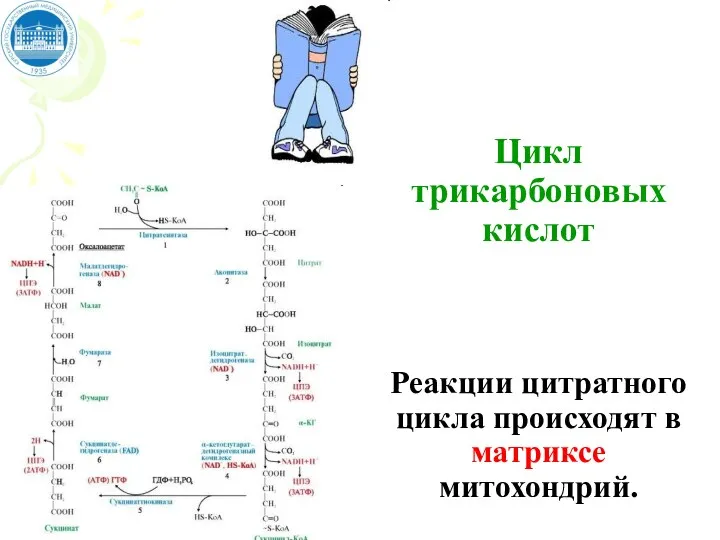
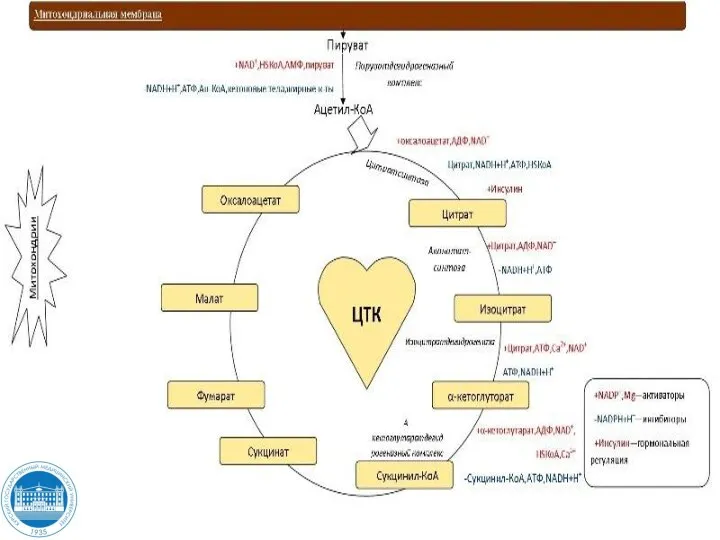
Цикл трикарбоновых кислот
Реакции цитратного цикла происходят в матриксе митохондрий.
Цикл трикарбоновых кислот
Реакции цитратного цикла происходят в матриксе митохондрий.

Цикл трикарбоновых кислот
Цикл Кребса — это ключевой этап дыхания всех клеток,
Цикл трикарбоновых кислот
Цикл Кребса — это ключевой этап дыхания всех клеток,

Цитратный цикл
(цикл трикарбоновых кислот) представляет собой совокупность
8 последовательных химических
Цитратный цикл (цикл трикарбоновых кислот) представляет собой совокупность 8 последовательных химических
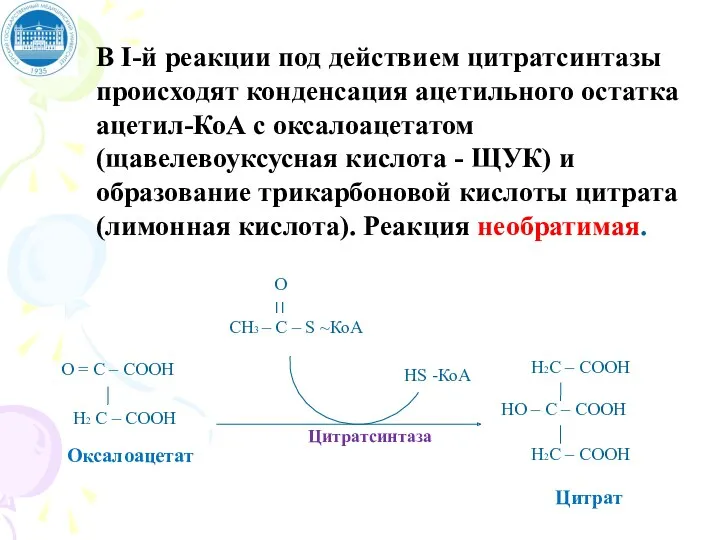
В I-й реакции под действием цитратсинтазы происходят конденсация ацетильного остатка ацетил-КоА
В I-й реакции под действием цитратсинтазы происходят конденсация ацетильного остатка ацетил-КоА
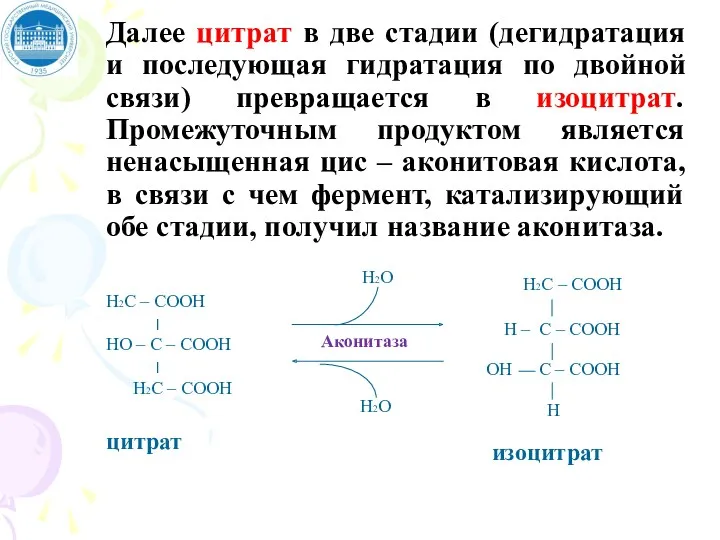
Далее цитрат в две стадии (дегидратация и последующая гидратация по двойной
Далее цитрат в две стадии (дегидратация и последующая гидратация по двойной
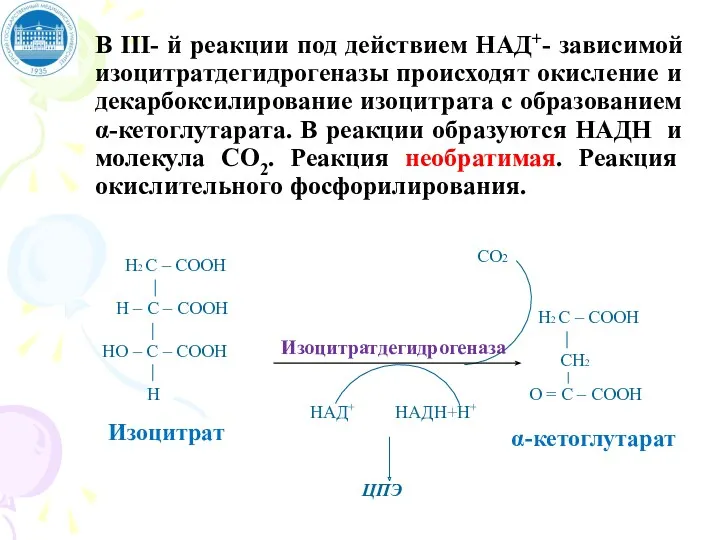
В III- й реакции под действием НАД+- зависимой изоцитратдегидрогеназы происходят окисление
В III- й реакции под действием НАД+- зависимой изоцитратдегидрогеназы происходят окисление
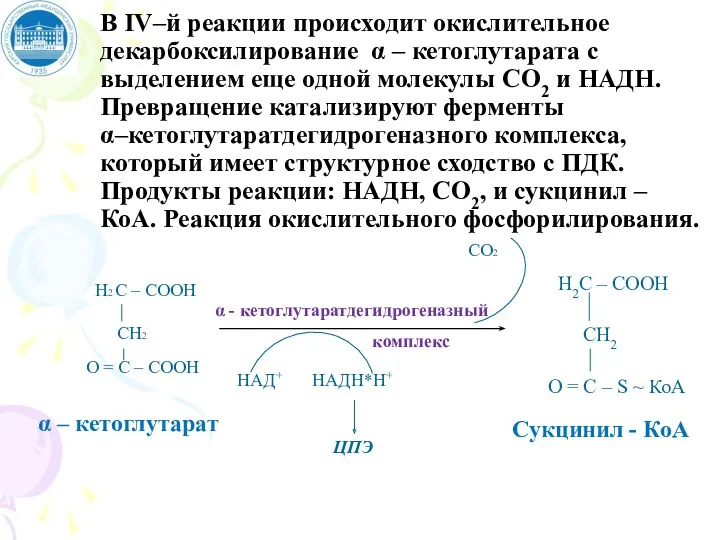
В IV–й реакции происходит окислительное декарбоксилирование α – кетоглутарата с выделением
В IV–й реакции происходит окислительное декарбоксилирование α – кетоглутарата с выделением
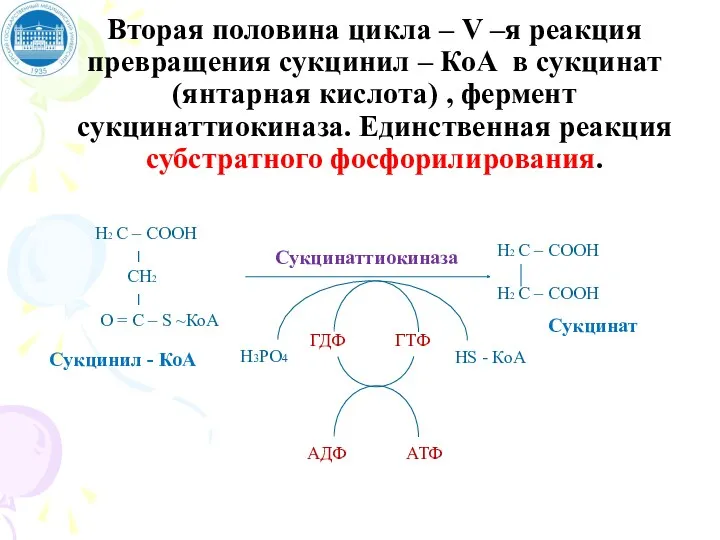
Вторая половина цикла – V –я реакция превращения сукцинил – КоА
Вторая половина цикла – V –я реакция превращения сукцинил – КоА
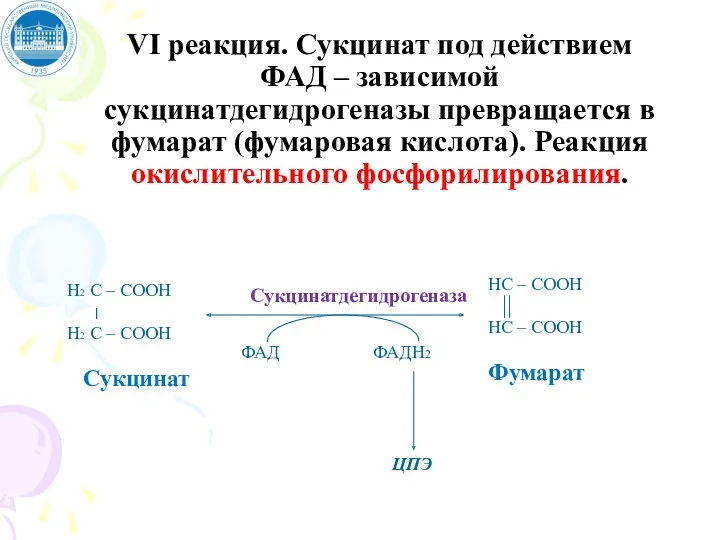
VI реакция. Сукцинат под действием ФАД – зависимой сукцинатдегидрогеназы превращается в
VI реакция. Сукцинат под действием ФАД – зависимой сукцинатдегидрогеназы превращается в
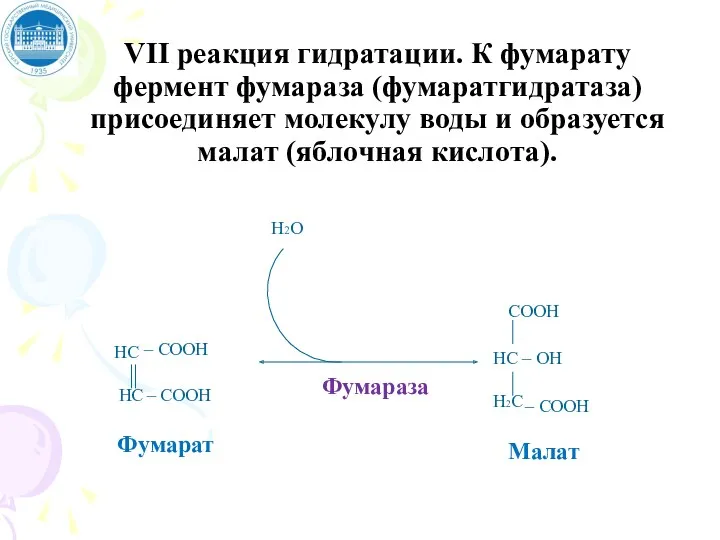
VII реакция гидратации. К фумарату фермент фумараза (фумаратгидратаза) присоединяет молекулу воды
VII реакция гидратации. К фумарату фермент фумараза (фумаратгидратаза) присоединяет молекулу воды
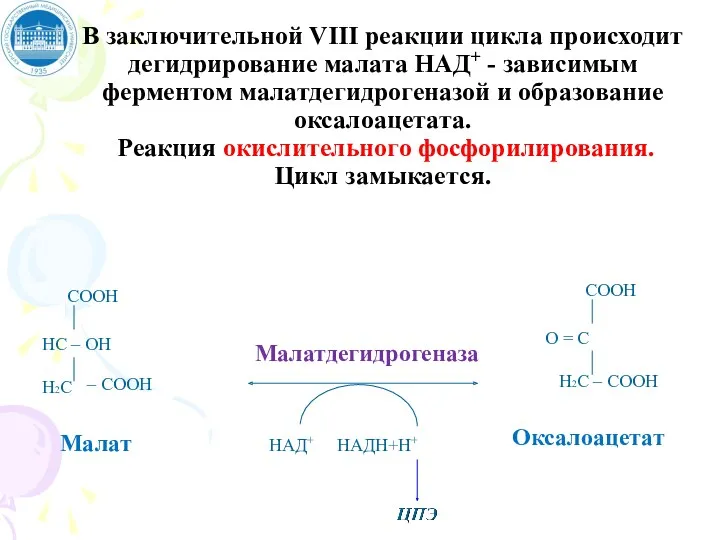
В заключительной VIII реакции цикла происходит дегидрирование малата НАД+ - зависимым
В заключительной VIII реакции цикла происходит дегидрирование малата НАД+ - зависимым

Функции цикла ЦТК
Интегративная функция — цикл является связующим звеном между реакциями
Функции цикла ЦТК
Интегративная функция — цикл является связующим звеном между реакциями

Функции цикла ЦТК
Водорододонорная функция — цикл Кребса поставляет на дыхательную цепь
Функции цикла ЦТК
Водорододонорная функция — цикл Кребса поставляет на дыхательную цепь

Таким образом, в ОПК происходит распад 3 – углеродного соединения пирувата
Таким образом, в ОПК происходит распад 3 – углеродного соединения пирувата

Баланс АТФ при окислении
глюкозы до С2О и Н2О
I. Гликолиз
А) 1.
Баланс АТФ при окислении
глюкозы до С2О и Н2О
I. Гликолиз
А) 1.

Мнемоническое правило
Для более легкого запоминания кислот, участвующих в цикле Кребса, существует
Мнемоническое правило
Для более легкого запоминания кислот, участвующих в цикле Кребса, существует

СИНТЕЗ ГЛЮКОЗЫ - ГЛЮКОНЕОГЕНЕЗ
СИНТЕЗ ГЛЮКОЗЫ - ГЛЮКОНЕОГЕНЕЗ

Глюконеогенез - это процесс синтеза глюкозы из веществ неуглеводной природы.
Глюконеогенез - это процесс синтеза глюкозы из веществ неуглеводной природы.

Важнейшей функцией глюконеогенеза является:
поддержание уровня глюкозы в крови в период длительного
Важнейшей функцией глюконеогенеза является:
поддержание уровня глюкозы в крови в период длительного

Глюконеогенез протекает
главным образом в печени
менее интенсивно - в корковом веществе
Глюконеогенез протекает
главным образом в печени
менее интенсивно - в корковом веществе

Включение различных субстратов в глюконеогенез зависит от физиологического состояния организма:
лактат является
Включение различных субстратов в глюконеогенез зависит от физиологического состояния организма:
лактат является
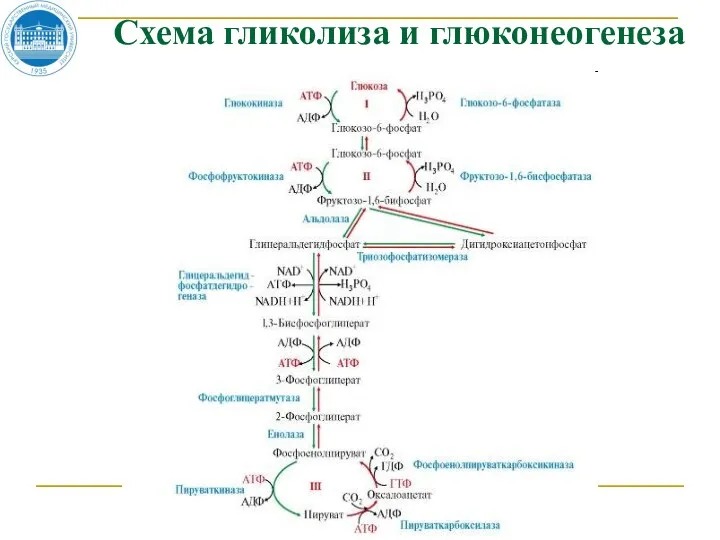
Схема гликолиза и глюконеогенеза
Схема гликолиза и глюконеогенеза
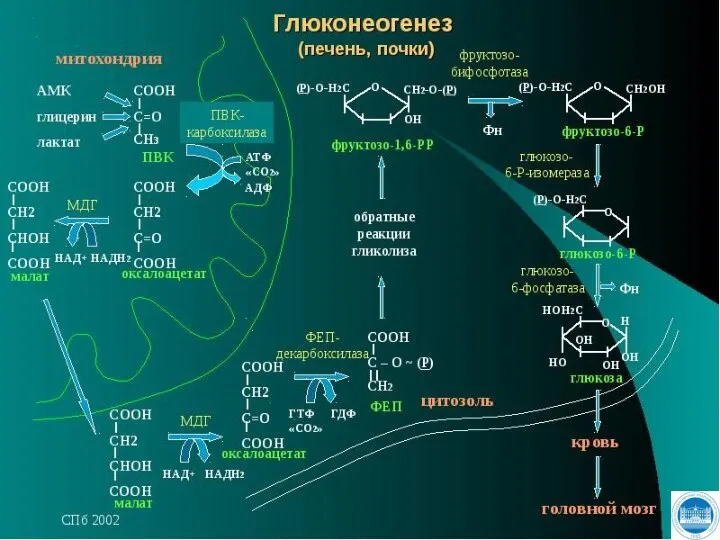

Глюконеогенез
Большинство реакций гликолиза и глюконеогенеза являются обратимыми и катализируются одними и
Глюконеогенез
Большинство реакций гликолиза и глюконеогенеза являются обратимыми и катализируются одними и
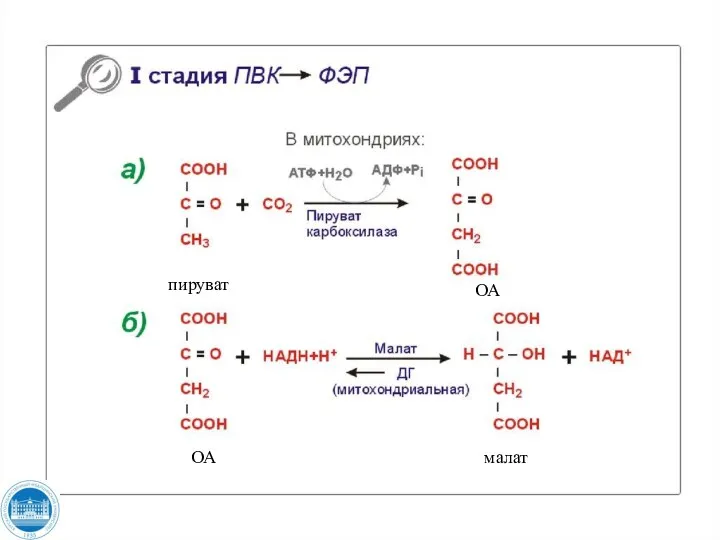
пируват
ОА
ОА
малат
пируват
ОА
ОА
малат
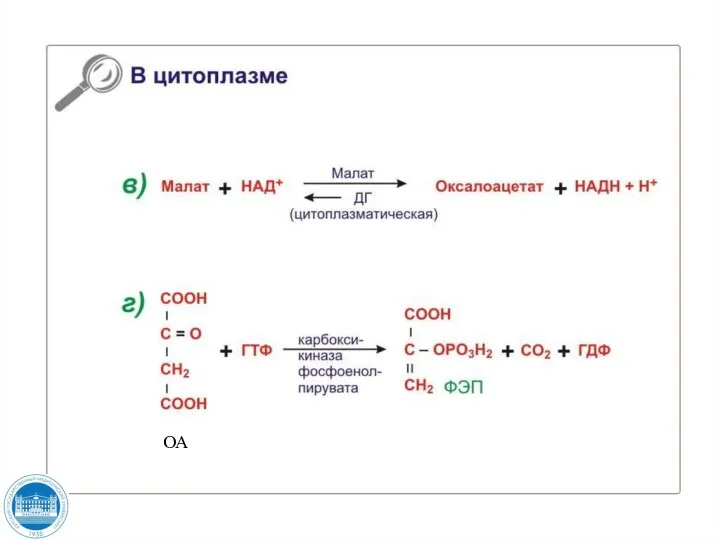
ОА
ОА

Цикл Кори
Использование лактата в качестве субстрата в глюконеогенезе связано с транспортом
Цикл Кори
Использование лактата в качестве субстрата в глюконеогенезе связано с транспортом

Благодарю за внимание
Благодарю за внимание

Лекция № 6
Глюконеогенез (продолжение).
Обмен гликогена.
Особенности обмена углеводов в различных органах и
Лекция № 6
Глюконеогенез (продолжение).
Обмен гликогена.
Особенности обмена углеводов в различных органах и
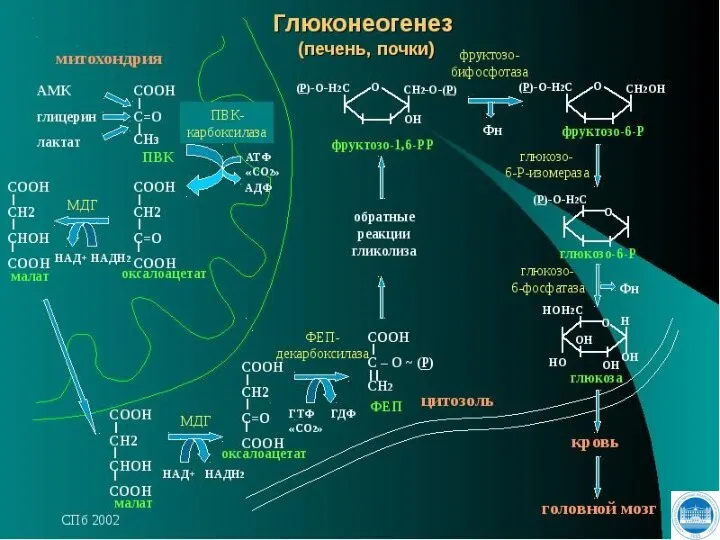
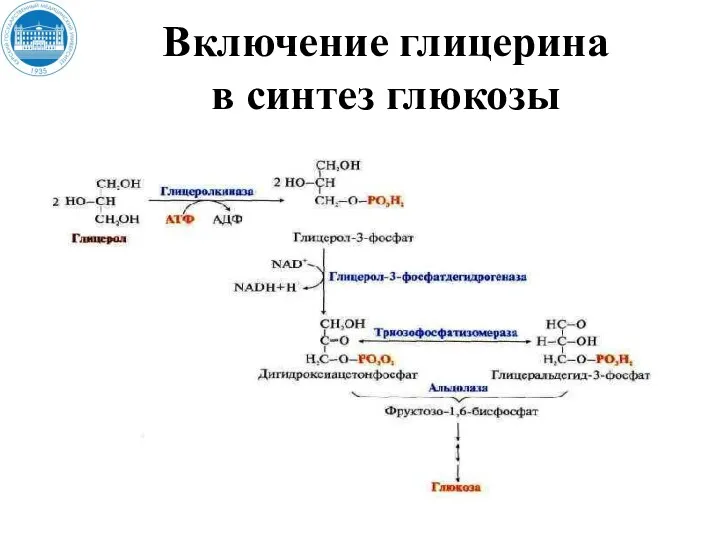
Включение глицерина
в синтез глюкозы
Включение глицерина
в синтез глюкозы
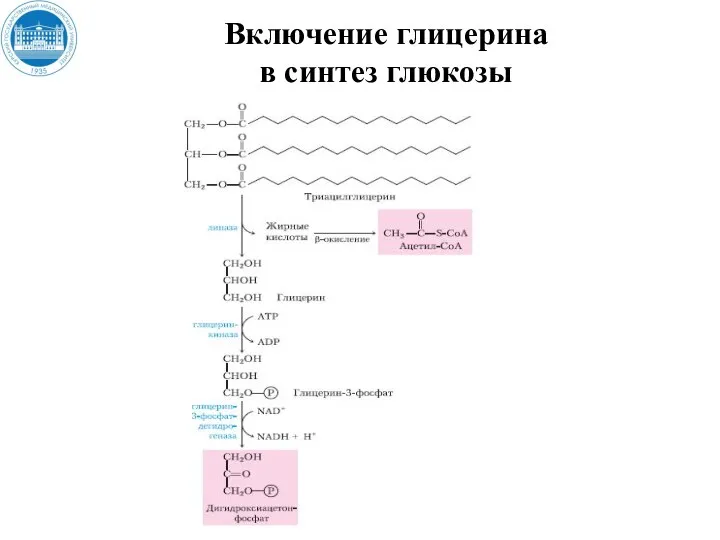
Включение глицерина
в синтез глюкозы
Включение глицерина
в синтез глюкозы
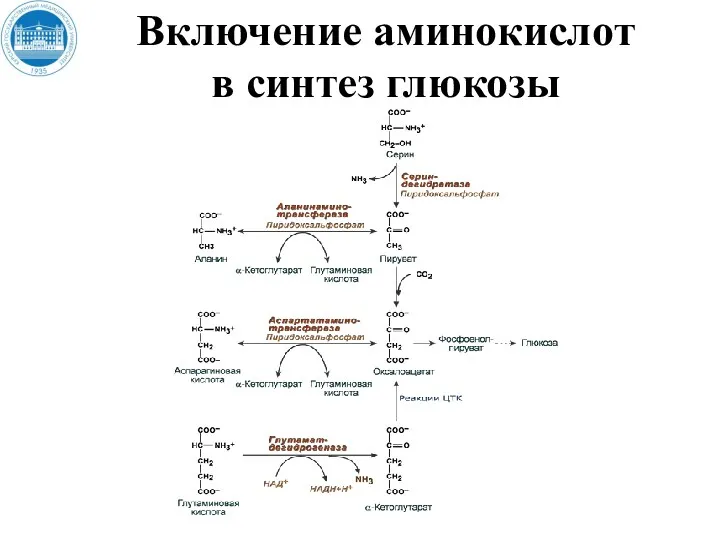
Включение аминокислот
в синтез глюкозы
Включение аминокислот
в синтез глюкозы

СИНТЕЗ ГЛИКОГЕНА (ГЛИКОГЕНОГЕНЕЗ), МОБИЛИЗАЦИЯ ГЛИКОГЕНА (ГЛИКОГЕНОЛИЗ).
СИНТЕЗ ГЛИКОГЕНА (ГЛИКОГЕНОГЕНЕЗ), МОБИЛИЗАЦИЯ ГЛИКОГЕНА (ГЛИКОГЕНОЛИЗ).
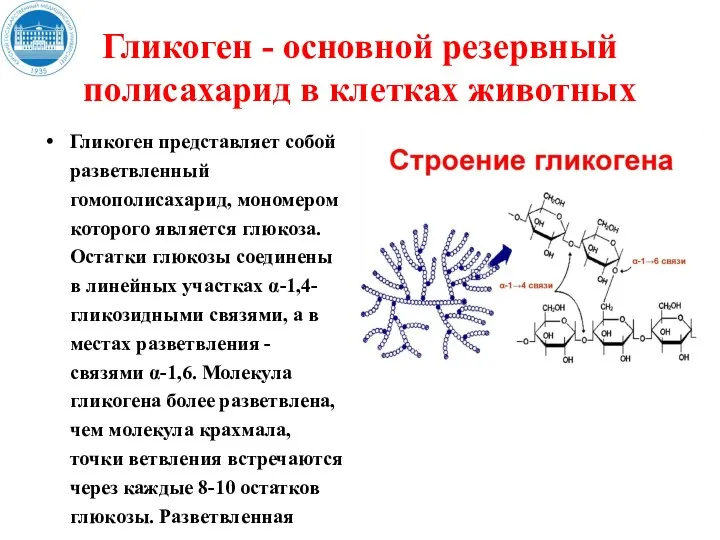
Гликоген - основной резервный полисахарид в клетках животных
Гликоген представляет собой разветвленный
Гликоген - основной резервный полисахарид в клетках животных
Гликоген представляет собой разветвленный
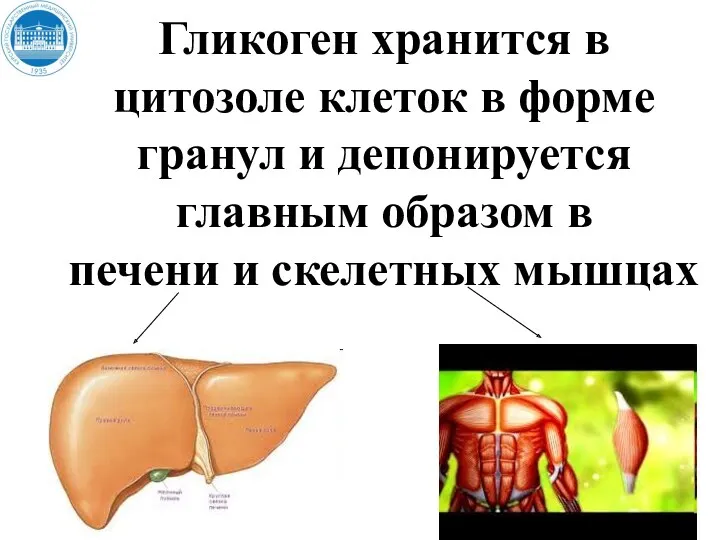
Гликоген хранится в цитозоле клеток в форме гранул и депонируется главным
Гликоген хранится в цитозоле клеток в форме гранул и депонируется главным

Гранулы гликогена плохо растворимы в воде и не влияют на осмотическое
Гранулы гликогена плохо растворимы в воде и не влияют на осмотическое

Синтез гликогена
Гликоген синтезируется в период пищеварения (абсорбтивный период: 1-2 часа после
Синтез гликогена
Гликоген синтезируется в период пищеварения (абсорбтивный период: 1-2 часа после

Синтез гликогена
Когда длина синтезируемой цепи увеличивается на 11-12 остатков глюкозы, фермент
Синтез гликогена
Когда длина синтезируемой цепи увеличивается на 11-12 остатков глюкозы, фермент
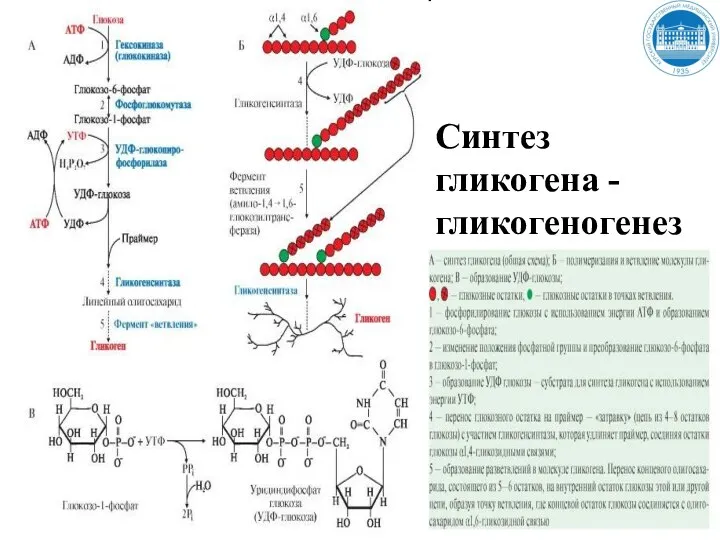
Синтез гликогена - гликогеногенез
Синтез гликогена - гликогеногенез

Мобилизация (распад) гликогена
Мобилизация (распад) гликогена происходит в интервалах
Мобилизация (распад) гликогена
Мобилизация (распад) гликогена происходит в интервалах

Мобилизация (распад) гликогена
Мобилизация гликогена в печени отличается от
Мобилизация (распад) гликогена
Мобилизация гликогена в печени отличается от
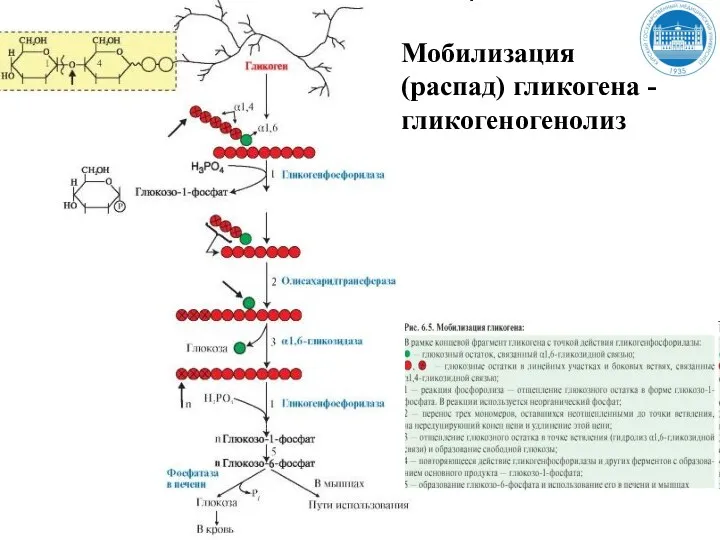
Мобилизация (распад) гликогена - гликогеногенолиз
Мобилизация (распад) гликогена - гликогеногенолиз

Гормоны, обеспечивающие переключение метаболических путей:
Переключение процессов синтеза и мобилизации гликогена в
Гормоны, обеспечивающие переключение метаболических путей:
Переключение процессов синтеза и мобилизации гликогена в

Регуляция метаболизма гликогена в печени
Под влиянием инсулина происходит:
ускорение транспорта глюкозы в
Регуляция метаболизма гликогена в печени
Под влиянием инсулина происходит:
ускорение транспорта глюкозы в

Регуляция метаболизма гликогена в печени
Под влиянием инсулина происходит:
активация фосфопротеинфосфатазой гранул гликогена,
Регуляция метаболизма гликогена в печени
Под влиянием инсулина происходит:
активация фосфопротеинфосфатазой гранул гликогена,

Первичным сигналом для синтеза инсулина и глюкагона является изменение концентрации глюкозы
Первичным сигналом для синтеза инсулина и глюкагона является изменение концентрации глюкозы

Регуляция метаболизма гликогена в печени
В период пищеварения концентрация глюкозы в
Регуляция метаболизма гликогена в печени
В период пищеварения концентрация глюкозы в

Особенности обмена глюкозы в различных тканях и органах
Особенности обмена глюкозы в различных тканях и органах
ОСОБЕННОСТИ УГЛЕВОДНОГО ОБМЕНА В ПЕЧЕНИ (ГЕПАТОЦИТЫ)
1. Превращение фруктозы и галактозы в
ОСОБЕННОСТИ УГЛЕВОДНОГО ОБМЕНА В ПЕЧЕНИ (ГЕПАТОЦИТЫ)
1. Превращение фруктозы и галактозы в

Обмен углеводов в печени
Одной из важнейших функций печени в процессах обмена
Обмен углеводов в печени
Одной из важнейших функций печени в процессах обмена

Обмен углеводов в печени
В печени катаболизм глюкозы представлен 2 процессами: 1)
Обмен углеводов в печени
В печени катаболизм глюкозы представлен 2 процессами: 1)

Обмен углеводов в печени
Гликолиз обеспечивает энергией клеточные реакции фосфорилирования, синтез белка;
Обмен углеводов в печени
Гликолиз обеспечивает энергией клеточные реакции фосфорилирования, синтез белка;

Обмен углеводов в печени
При аэробных условиях происходит сочетание гликолиза, протекающего в
Обмен углеводов в печени
При аэробных условиях происходит сочетание гликолиза, протекающего в

Обмен углеводов в печени
В печени протекают гликогенез и гликогенолиз. Эти процессы
Обмен углеводов в печени
В печени протекают гликогенез и гликогенолиз. Эти процессы

Обмен углеводов в мышцах
Цель мышечной клетки – наиболее эффективно использовать
Обмен углеводов в мышцах
Цель мышечной клетки – наиболее эффективно использовать

Обмен углеводов в мышцах
Цитоплазма мышечных клеток содержит в высоких концентрациях
Обмен углеводов в мышцах
Цитоплазма мышечных клеток содержит в высоких концентрациях

Обмен углеводов в мышцах
В мышцах идет гликогенез, мышца осуществляет лишь
Обмен углеводов в мышцах
В мышцах идет гликогенез, мышца осуществляет лишь
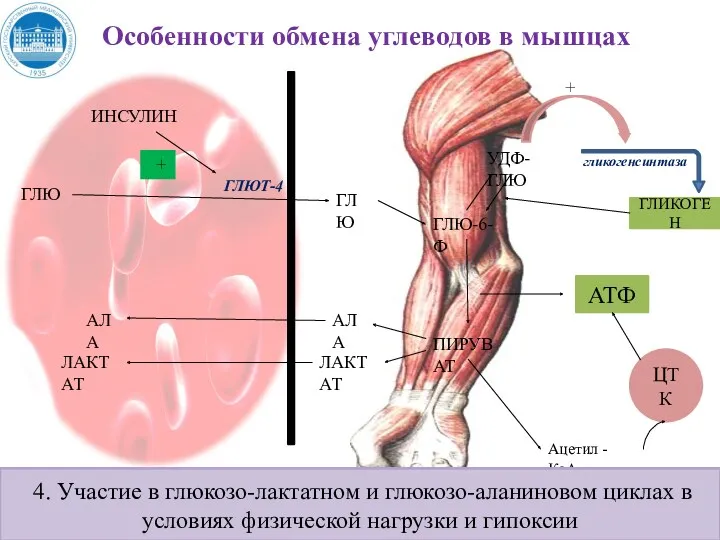
Особенности обмена углеводов в мышцах
ИНСУЛИН
ГЛЮ
ГЛЮ
ГЛЮТ-4
ГЛЮ-6-Ф
ПИРУВАТ
АТФ
ЦТК
Ацетил - КоА
1. Активное использование глюкозы
Особенности обмена углеводов в мышцах
ИНСУЛИН
ГЛЮ
ГЛЮ
ГЛЮТ-4
ГЛЮ-6-Ф
ПИРУВАТ
АТФ
ЦТК
Ацетил - КоА
1. Активное использование глюкозы

Обмен углеводов в мышцах
Фосфорилирование глюкозы в мышцах происходит под дейстием гексокиназы,
Обмен углеводов в мышцах
Фосфорилирование глюкозы в мышцах происходит под дейстием гексокиназы,
Обмен углеводов
в сердечной мышце
Ни медленно сокращающаяся гладкая мышечная ткань,
Обмен углеводов
в сердечной мышце
Ни медленно сокращающаяся гладкая мышечная ткань,
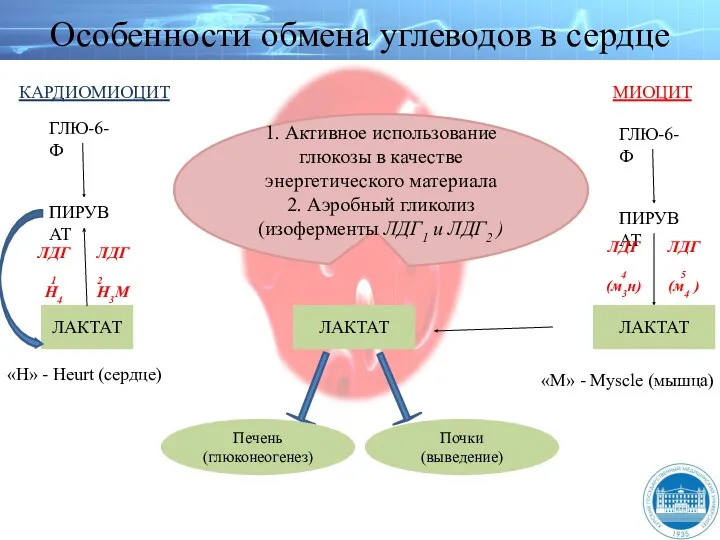
Особенности обмена углеводов в сердце
ЛАКТАТ
Печень (глюконеогенез)
Почки (выведение)
ЛАКТАТ
ЛАКТАТ
ПИРУВАТ
ГЛЮ-6-Ф
ЛДГ4
(м3н)
ЛДГ5
(м4 )
«М» - Myscle (мышца)
ГЛЮ-6-Ф
ПИРУВАТ
ЛДГ1
Н4
ЛДГ2
Н3М
«Н»
Особенности обмена углеводов в сердце
ЛАКТАТ
Печень (глюконеогенез)
Почки (выведение)
ЛАКТАТ
ЛАКТАТ
ПИРУВАТ
ГЛЮ-6-Ф
ЛДГ4
(м3н)
ЛДГ5
(м4 )
«М» - Myscle (мышца)
ГЛЮ-6-Ф
ПИРУВАТ
ЛДГ1
Н4
ЛДГ2
Н3М
«Н»
Кровь
Глю-6-ф
Глю
Кет. тела
Ацетил-КоА
ЖК
ГФЛ
Ацетилхолин
Сфингофосфо-
липиды
Мембраны
нейронов
и миелиновый
футляр
СО2
АТФ
ЦТК
Гликолипиды
КТ
2-3%
ПЕПТОЗЫ
НАДФНН+
НК
97-98%
ГЛУ
ГОМК, ГАМК
ГЛН
NH3 (обезвреживание)
Синтез сложных липидов
10% углеродных атомов
Кровь
Глю-6-ф
Глю
Кет. тела
Ацетил-КоА
ЖК
ГФЛ
Ацетилхолин
Сфингофосфо-
липиды
Мембраны
нейронов
и миелиновый
футляр
СО2
АТФ
ЦТК
Гликолипиды
КТ
2-3%
ПЕПТОЗЫ
НАДФНН+
НК
97-98%
ГЛУ
ГОМК, ГАМК
ГЛН
NH3 (обезвреживание)
Синтез сложных липидов
10% углеродных атомов

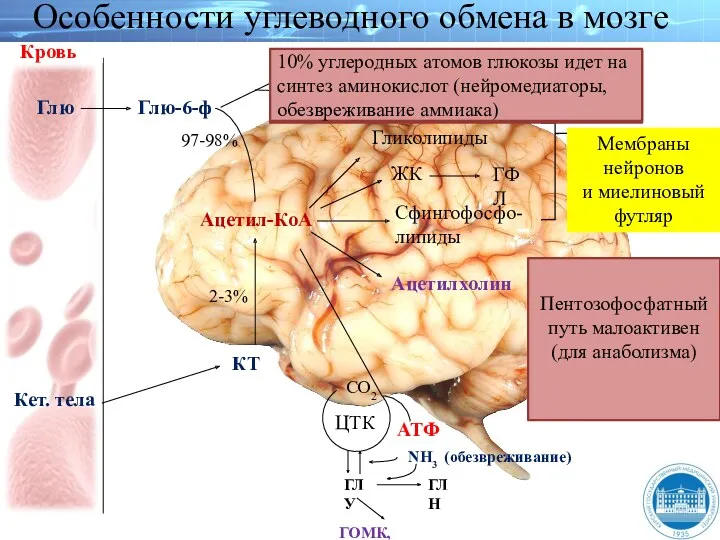
Обмен углеводов в мозге
По сравнению со всеми органами тела функций
Обмен углеводов в мозге
По сравнению со всеми органами тела функций

Обмен углеводов в мозге
Высокая активность митохондриальных ферментов цикла лимонной кислоты
Обмен углеводов в мозге
Высокая активность митохондриальных ферментов цикла лимонной кислоты
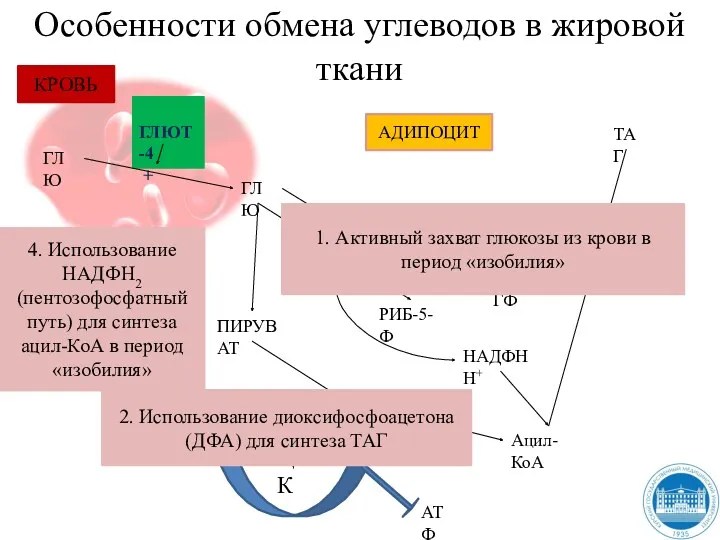
Особенности обмена углеводов в жировой ткани
ГЛЮТ-4
+
ГЛЮ
ГЛЮ
ДФА
α-ГФ
ТАГ
Ацил-КоА
АДИПОЦИТ
ПИРУВАТ
Ацетил-КоА
ЦТК
АТФ
РИБ-5-Ф
НАДФНН+
КРОВЬ
1. Активный захват глюкозы из
Особенности обмена углеводов в жировой ткани
ГЛЮТ-4
+
ГЛЮ
ГЛЮ
ДФА
α-ГФ
ТАГ
Ацил-КоА
АДИПОЦИТ
ПИРУВАТ
Ацетил-КоА
ЦТК
АТФ
РИБ-5-Ф
НАДФНН+
КРОВЬ
1. Активный захват глюкозы из

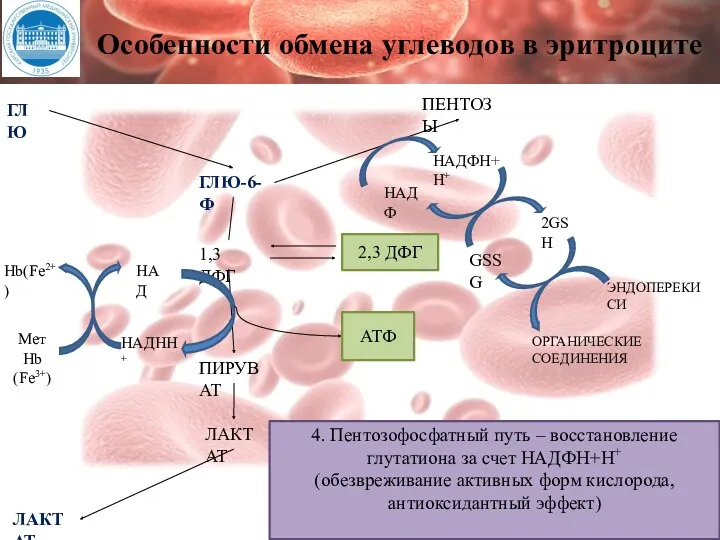
Обмен углеводов в эритроцитах
Эритроциты не содержат ядра, митохондрий. В эритроците не
Обмен углеводов в эритроцитах
Эритроциты не содержат ядра, митохондрий. В эритроците не
Особенности обмена углеводов в эритроците
ГЛЮ
ГЛЮ-6-Ф
1,3 ДФГ
ПИРУВАТ
ЛАКТАТ
ЛАКТАТ
НАД
НАДНН+
Hb(Fe2+)
Мет Hb
(Fe3+)
АТФ
2,3 ДФГ
ПЕНТОЗЫ
НАДФН+Н+
НАДФ
2GSH
GSSG
ОРГАНИЧЕСКИЕ СОЕДИНЕНИЯ
ЭНДОПЕРЕКИСИ
1. Анаэробный гликолиз
Особенности обмена углеводов в эритроците
ГЛЮ
ГЛЮ-6-Ф
1,3 ДФГ
ПИРУВАТ
ЛАКТАТ
ЛАКТАТ
НАД
НАДНН+
Hb(Fe2+)
Мет Hb
(Fe3+)
АТФ
2,3 ДФГ
ПЕНТОЗЫ
НАДФН+Н+
НАДФ
2GSH
GSSG
ОРГАНИЧЕСКИЕ СОЕДИНЕНИЯ
ЭНДОПЕРЕКИСИ
1. Анаэробный гликолиз
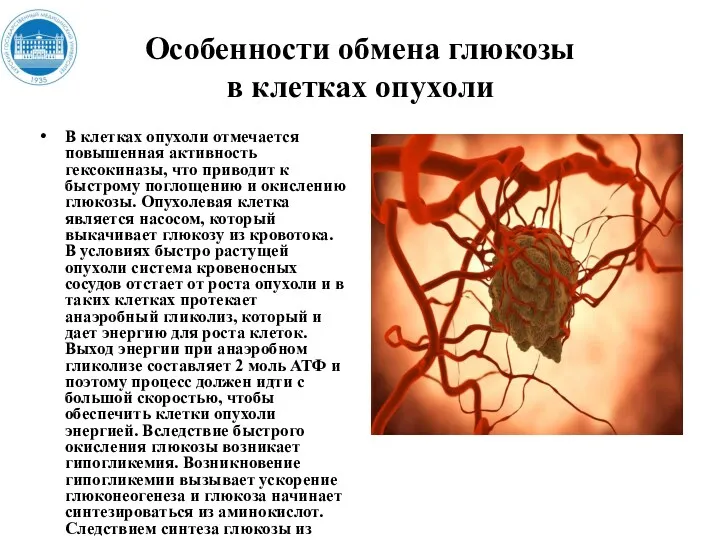
Особенности обмена глюкозы
в клетках опухoли
В клетках опухоли отмечается повышенная активность
Особенности обмена глюкозы
в клетках опухoли
В клетках опухоли отмечается повышенная активность
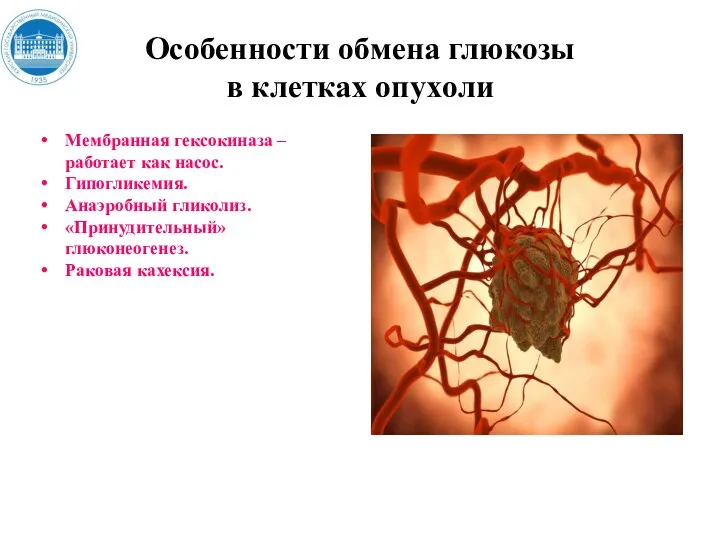
Особенности обмена глюкозы
в клетках опухoли
Мембранная гексокиназа – работает как насос.
Гипогликемия.
Анаэробный
Особенности обмена глюкозы
в клетках опухoли
Мембранная гексокиназа – работает как насос.
Гипогликемия.
Анаэробный

Благодарю за внимание
Благодарю за внимание

Лекция №7
Регуляция
и
патология углеводного обмена
Лекция №7
Регуляция
и
патология углеводного обмена

Регуляция обмена
углеводов состоит из регуляции:
Соотношение между процессами катаболизма и
Регуляция обмена
углеводов состоит из регуляции:
Соотношение между процессами катаболизма и

Регуляции гликолиза
Основное значение гликолиза - синтез АТФ, поэтому его скорость
Регуляции гликолиза
Основное значение гликолиза - синтез АТФ, поэтому его скорость

Регуляции гликолиза
Вовлечение глюкозных остатков в процесс гликолиза обеспечивает важная реакция и
Регуляции гликолиза
Вовлечение глюкозных остатков в процесс гликолиза обеспечивает важная реакция и

Регуляции гликолиза
В печени преобладает фермент – глюкокиназа, которая не ингибируется глюкозо-6-фосфатом.
Регуляции гликолиза
В печени преобладает фермент – глюкокиназа, которая не ингибируется глюкозо-6-фосфатом.
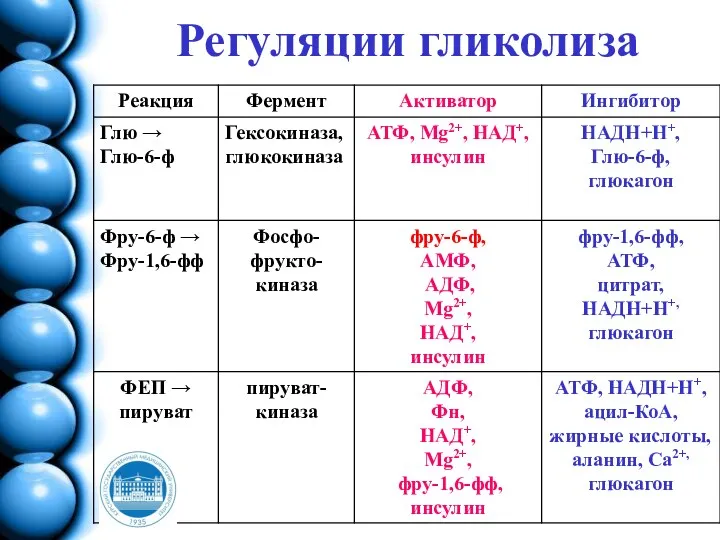
Регуляции гликолиза
Регуляции гликолиза

Гормональная регуляция гликолиза
Глюкагон и инсулин влияют на синтез ключевых ферментов, используя
Гормональная регуляция гликолиза
Глюкагон и инсулин влияют на синтез ключевых ферментов, используя
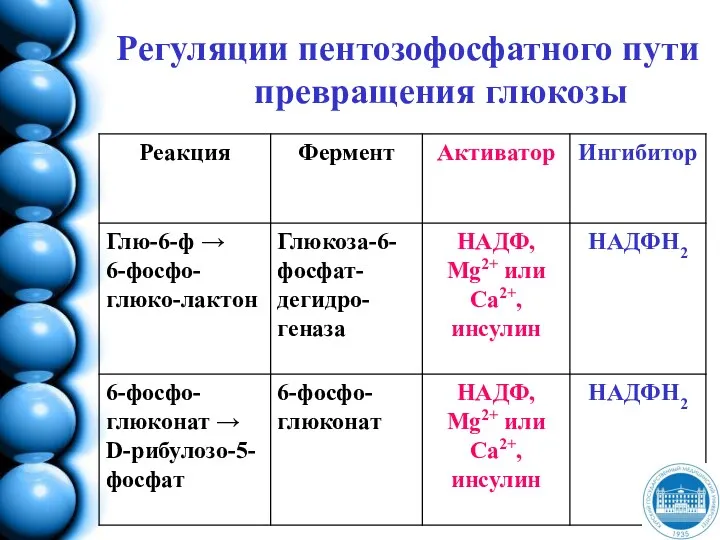
Регуляции пентозофосфатного пути превращения глюкозы
Регуляции пентозофосфатного пути превращения глюкозы
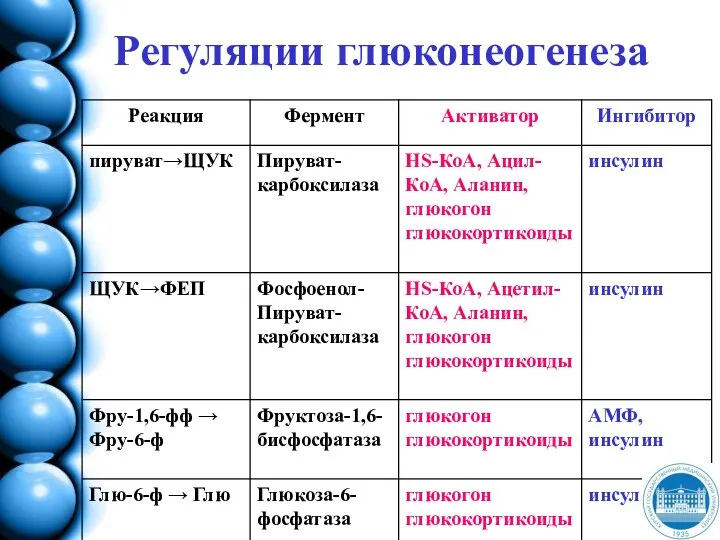
Регуляции глюконеогенеза
Регуляции глюконеогенеза

Гормональная регуляция глюконеогенеза
В постабсорбтивный период глюкагон повышает транскрипцию генов и синтез
Гормональная регуляция глюконеогенеза
В постабсорбтивный период глюкагон повышает транскрипцию генов и синтез
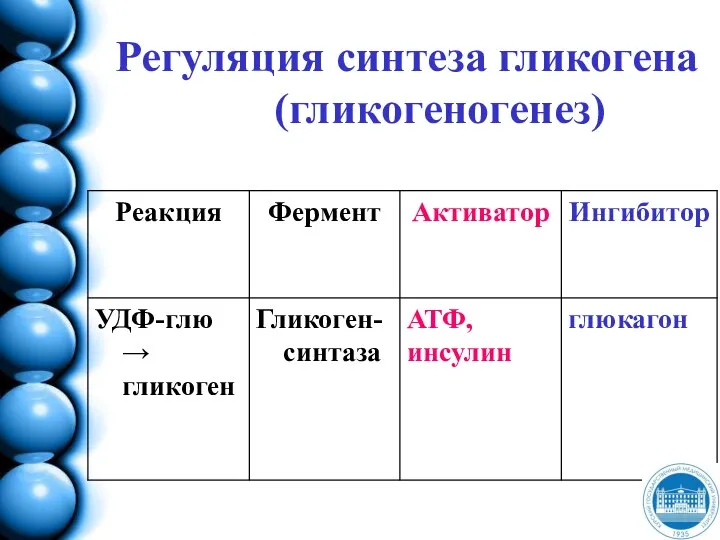
Регуляция синтеза гликогена
(гликогеногенез)
Регуляция синтеза гликогена
(гликогеногенез)
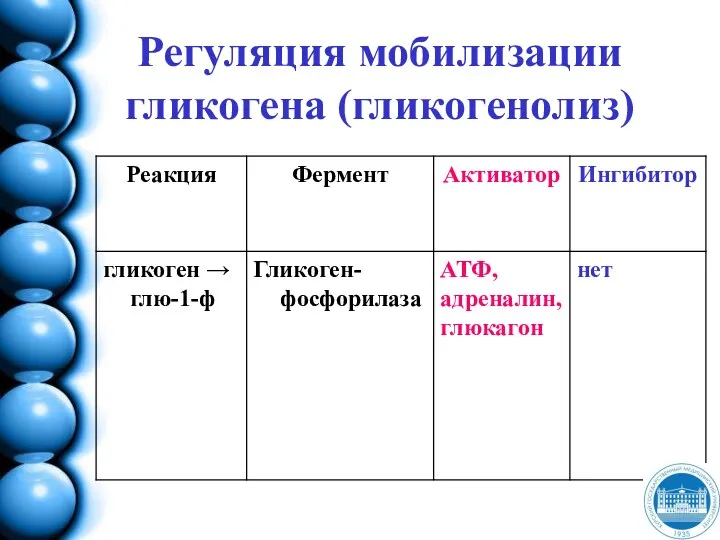
Регуляция мобилизации гликогена (гликогенолиз)
Регуляция мобилизации гликогена (гликогенолиз)
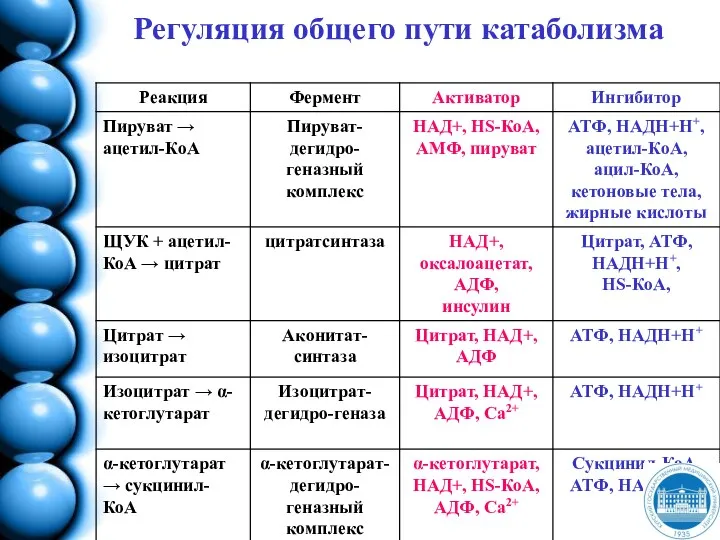
Регуляция общего пути катаболизма
Регуляция общего пути катаболизма

Механизм действия инсулина – гипогликемического гормона
Повышает проницаемость клеточных мембран для глюкозы,
Механизм действия инсулина – гипогликемического гормона
Повышает проницаемость клеточных мембран для глюкозы,

Механизм действия инсулина
Подавляет синтез ферментов глюконеогенеза, препятствует избыточному катаболизму жиров и
Механизм действия инсулина
Подавляет синтез ферментов глюконеогенеза, препятствует избыточному катаболизму жиров и

Инсулин активирует:
Ферменты гликолиза: гексокиназу, фосфофруктокиназу, пируваткиназу.
Ферменты пентозофосфатного пути: глюкозо-6-фосфатдегидрогеназу, 6-фосфоглюконатдегидрогеназу.
Ферменты гликогенеза
Инсулин активирует:
Ферменты гликолиза: гексокиназу, фосфофруктокиназу, пируваткиназу.
Ферменты пентозофосфатного пути: глюкозо-6-фосфатдегидрогеназу, 6-фосфоглюконатдегидрогеназу.
Ферменты гликогенеза

Механизм действия адреналина и глюкагона
Усиливают распад гликогена в мышцах и печени,
Механизм действия адреналина и глюкагона
Усиливают распад гликогена в мышцах и печени,

Механизм действия глюкокортикоидов
Усиливают глюконеогенез за счет индукции синтеза в клетках
Механизм действия глюкокортикоидов
Усиливают глюконеогенез за счет индукции синтеза в клетках

Гипергликемические гормоны:
Адреналин, глюкогон – активация фосфорилазы.
Кортикостероиды – активация (усиление синтеза) ферментов
Гипергликемические гормоны:
Адреналин, глюкогон – активация фосфорилазы.
Кортикостероиды – активация (усиление синтеза) ферментов

Регуляция скоростей синтеза и распада гликогена в печени поддерживает постоянство концентрации
Регуляция скоростей синтеза и распада гликогена в печени поддерживает постоянство концентрации
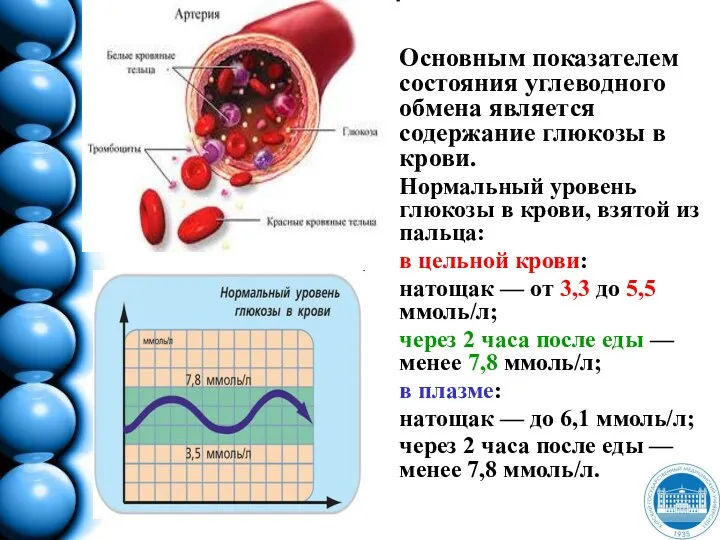
Основным показателем состояния углеводного обмена является содержание глюкозы в крови.
Нормальный
Основным показателем состояния углеводного обмена является содержание глюкозы в крови.
Нормальный
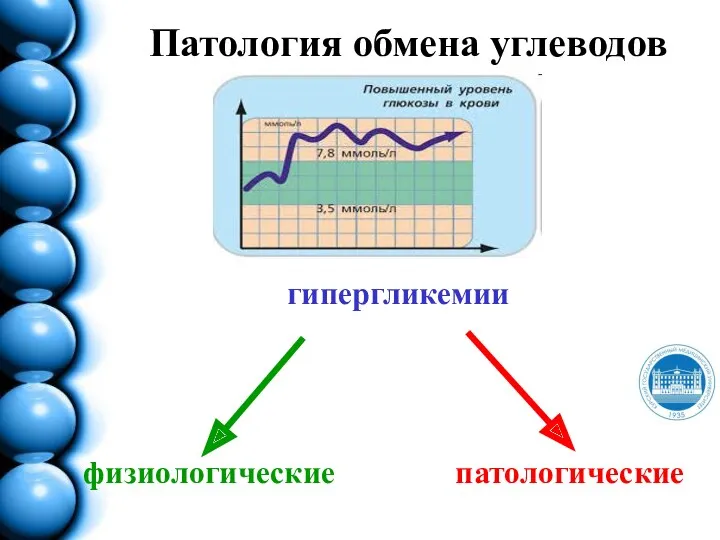
Патология обмена углеводов
гипергликемии
физиологические патологические
Патология обмена углеводов
гипергликемии
физиологические патологические

Патология обмена углеводов
К физиологическим гипергликемиям относятся:
алиментарные, возникающие при одномоментном приеме
Патология обмена углеводов
К физиологическим гипергликемиям относятся:
алиментарные, возникающие при одномоментном приеме

Патология обмена углеводов
Патологические гипергликемии обусловлены нарушением оптимального соотношения между секрецией гормонов
Патология обмена углеводов
Патологические гипергликемии обусловлены нарушением оптимального соотношения между секрецией гормонов

Клинические рекомендации Сахарный диабет 1 типа у взрослых, 2019 https://cr.minzdrav.gov.ru/recomend/286_2
Клинические
Клинические рекомендации Сахарный диабет 1 типа у взрослых, 2019 https://cr.minzdrav.gov.ru/recomend/286_2 Клинические

В основе сахарного диабета лежат следующие молекулярные дефекты:
Нарушение превращения проинсулина в
В основе сахарного диабета лежат следующие молекулярные дефекты:
Нарушение превращения проинсулина в

В основе сахарного диабета лежат следующие молекулярные дефекты:
Дефект рецепторов инсулина. У
В основе сахарного диабета лежат следующие молекулярные дефекты:
Дефект рецепторов инсулина. У

Важнейшими биохимическими признаками сахарного диабета являются:
Гипергликемия. В результате недостатка инсулина нарушается
Важнейшими биохимическими признаками сахарного диабета являются:
Гипергликемия. В результате недостатка инсулина нарушается
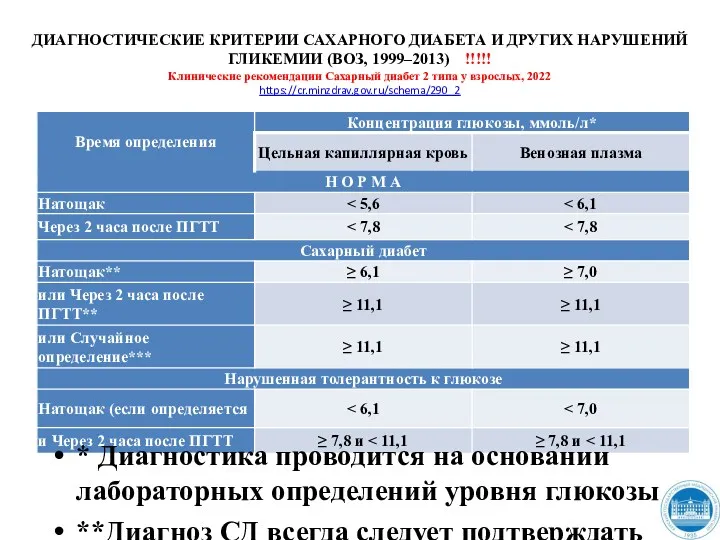
ДИАГНОСТИЧЕСКИЕ КРИТЕРИИ САХАРНОГО ДИАБЕТА И ДРУГИХ НАРУШЕНИЙ ГЛИКЕМИИ (ВОЗ, 1999–2013) !!!!!
Клинические
ДИАГНОСТИЧЕСКИЕ КРИТЕРИИ САХАРНОГО ДИАБЕТА И ДРУГИХ НАРУШЕНИЙ ГЛИКЕМИИ (ВОЗ, 1999–2013) !!!!! Клинические
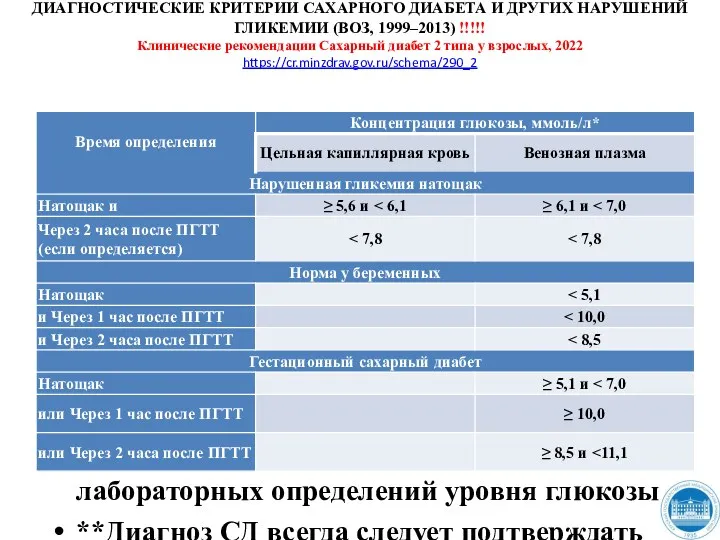
ДИАГНОСТИЧЕСКИЕ КРИТЕРИИ САХАРНОГО ДИАБЕТА И ДРУГИХ НАРУШЕНИЙ ГЛИКЕМИИ (ВОЗ, 1999–2013) !!!!!
Клинические
ДИАГНОСТИЧЕСКИЕ КРИТЕРИИ САХАРНОГО ДИАБЕТА И ДРУГИХ НАРУШЕНИЙ ГЛИКЕМИИ (ВОЗ, 1999–2013) !!!!! Клинические

Время определения концентрация глюкозы !!!!!
Перевод глюкозы крови из ммоль/л в мг/дл:
1
Время определения концентрация глюкозы !!!!!
Перевод глюкозы крови из ммоль/л в мг/дл:
1

ПГТТ – пероральный глюкозотолерантный тест. Проводится в случае сомнительных значений гликемии
ПГТТ – пероральный глюкозотолерантный тест. Проводится в случае сомнительных значений гликемии

ОСТРЫЕ ОСЛОЖНЕНИЯ САХАРНОГО ДИАБЕТА
Диабетический кетоацидоз (дка, диабетическая кетоацидотическая кома)
Гиперосмолярное гипергликемическое состояние
ОСТРЫЕ ОСЛОЖНЕНИЯ САХАРНОГО ДИАБЕТА
Диабетический кетоацидоз (дка, диабетическая кетоацидотическая кома)
Гиперосмолярное гипергликемическое состояние

1. Диабетический кетоацидоз (дка, диабетическая кетоацидотическая кома) !
ДКА – требующая экстренной
1. Диабетический кетоацидоз (дка, диабетическая кетоацидотическая кома) !
ДКА – требующая экстренной

Лабораторные изменения: диагностика и дифференциальная диагностика ДКА
Общий анализ мочи: Глюкозурия, кетонурия,
Лабораторные изменения: диагностика и дифференциальная диагностика ДКА
Общий анализ мочи: Глюкозурия, кетонурия,

2. ГИПЕРОСМОЛЯРНОЕ ГИПЕРГЛИКЕМИЧЕСКОЕ СОСТОЯНИЕ (ГГС)!
ГГС – острая декомпенсация СД, с резко
2. ГИПЕРОСМОЛЯРНОЕ ГИПЕРГЛИКЕМИЧЕСКОЕ СОСТОЯНИЕ (ГГС)!
ГГС – острая декомпенсация СД, с резко

Лабораторные изменения: диагностика и дифференциальная диагностика ГГС
Общий анализ мочи: Общий анализ
Лабораторные изменения: диагностика и дифференциальная диагностика ГГС
Общий анализ мочи: Общий анализ

3. МОЛОЧНОКИСЛЫЙ АЦИДОЗ (ЛАКТАТАЦИДОЗ) !
Лактатацидоз – метаболический ацидоз с большой анионной
3. МОЛОЧНОКИСЛЫЙ АЦИДОЗ (ЛАКТАТАЦИДОЗ) !
Лактатацидоз – метаболический ацидоз с большой анионной

Лабораторные изменения: диагностика и дифференциальная диагностика лактатацидоза
Биохимический анализ крови: Лактат >
Лабораторные изменения: диагностика и дифференциальная диагностика лактатацидоза
Биохимический анализ крови: Лактат >

4. ГИПОГЛИКЕМИЯ И ГИПОГЛИКЕМИЧЕСКАЯ КОМА !
Гипогликемия – уровень глюкозы плазмы <
4. ГИПОГЛИКЕМИЯ И ГИПОГЛИКЕМИЧЕСКАЯ КОМА !
Гипогликемия – уровень глюкозы плазмы <

Лабораторные изменения: диагностика и дифференциальная диагностика гипогликемии и гипогликемической комы
Биохимический
Лабораторные изменения: диагностика и дифференциальная диагностика гипогликемии и гипогликемической комы
Биохимический

Гликогеновые болезни - это группа наследственных заболеваний, причинами которых являются дефекты ферментов,
Гликогеновые болезни - это группа наследственных заболеваний, причинами которых являются дефекты ферментов,
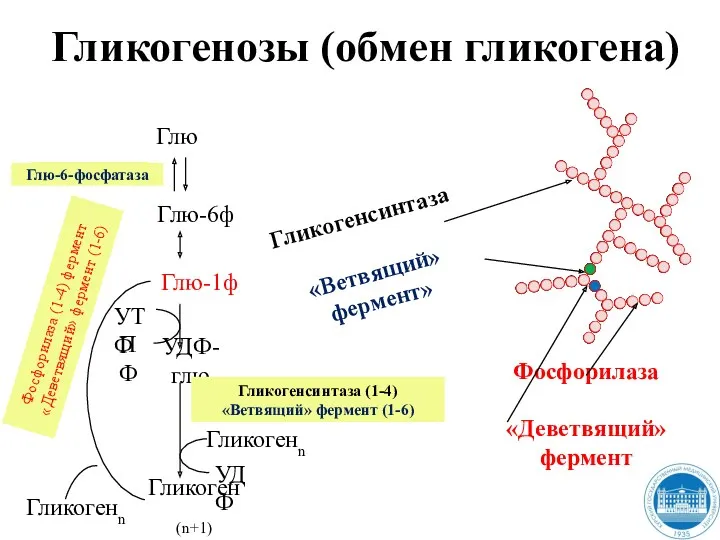
Гликогенозы (обмен гликогена)
Глю
Глю-6ф
Глю-1ф
УДФ-глю
Гликоген(n+1)
Гликогенсинтаза (1-4) «Ветвящий» фермент (1-6)
Гликогенn
УДФ
Глю-6-фосфатаза
УТФ
ПФ
Гликогенn
Фосфорилаза (1-4) фермент
«Деветвящий» фермент (1-6)
Гликогенсинтаза
Гликогенозы (обмен гликогена)
Глю
Глю-6ф
Глю-1ф
УДФ-глю
Гликоген(n+1)
Гликогенсинтаза (1-4) «Ветвящий» фермент (1-6)
Гликогенn
УДФ
Глю-6-фосфатаза
УТФ
ПФ
Гликогенn
Фосфорилаза (1-4) фермент
«Деветвящий» фермент (1-6)
Гликогенсинтаза

Гликогенозы
Всего известно 12 типов гликогенозов. По патогенетическому признаку гликогенозы делят:
печеночные – 0,
Гликогенозы
Всего известно 12 типов гликогенозов. По патогенетическому признаку гликогенозы делят:
печеночные – 0,
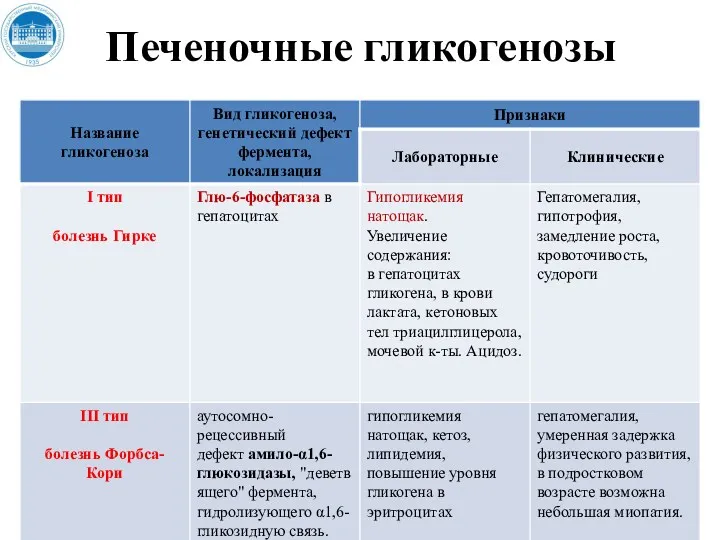
Печеночные гликогенозы
Печеночные гликогенозы

Печеночные гликогенозы
Печеночные гликогенозы

Мышечные гликогенозы
Мышечные гликогенозы

Смешанные гликогенозы
Смешанные гликогенозы
 Классификация лиан. Лианы как жизненная форма
Классификация лиан. Лианы как жизненная форма Сучасна клітинна теорія. Клітина – елементарна цілісна жива система
Сучасна клітинна теорія. Клітина – елементарна цілісна жива система 7 самых ядовитых растений России
7 самых ядовитых растений России Обмен веществ и энергии. Характеристика метаболизма
Обмен веществ и энергии. Характеристика метаболизма Отделы Плауновидные, Хвощевидные, Папоротниковидные, особенности строения и жизнедеятельности
Отделы Плауновидные, Хвощевидные, Папоротниковидные, особенности строения и жизнедеятельности Бактериофаги. Особенности бактериофагов
Бактериофаги. Особенности бактериофагов Нуклеиновые кислоты. Биология. 9 класс
Нуклеиновые кислоты. Биология. 9 класс Многообразие паукообразных, их роль в природе
Многообразие паукообразных, их роль в природе Западно-алтайский национальный природный заповедник
Западно-алтайский национальный природный заповедник Вплив електричного поля на живі організми
Вплив електричного поля на живі організми Путешествие в удивительный мир растений
Путешествие в удивительный мир растений Репродукція клітин
Репродукція клітин Многообразие паукообразных, их роль в природе
Многообразие паукообразных, их роль в природе Биотехнология. Новые направления в биологии
Биотехнология. Новые направления в биологии Насекомые. Кто где живет
Насекомые. Кто где живет Ферменты. Лекции 1-4
Ферменты. Лекции 1-4 Ботаника. Отделы растений
Ботаника. Отделы растений Аутекологічні особливості Вльшанки в НПП Гомільшанські ліси
Аутекологічні особливості Вльшанки в НПП Гомільшанські ліси Микробиология. Физиология микроорганизмов
Микробиология. Физиология микроорганизмов Селекция. Приручение животных
Селекция. Приручение животных Отряд хищные
Отряд хищные Медузы. Стадии жизненного цикла
Медузы. Стадии жизненного цикла Значение бактерий в природе и в жизни человека
Значение бактерий в природе и в жизни человека Анализирующее скрещивание. Дигибридное скрещивание. Третий закон Менделя
Анализирующее скрещивание. Дигибридное скрещивание. Третий закон Менделя Популяция как форма существования вида
Популяция как форма существования вида Авторская методическая разработка по теме Слуховой анализатор.Строение уха.
Авторская методическая разработка по теме Слуховой анализатор.Строение уха. История эволюционизма
История эволюционизма Биоэнергетика. Биологическое окисление. Биологические виды энергии
Биоэнергетика. Биологическое окисление. Биологические виды энергии