Энергетика мышечного сокращения. Ферменты в толстых нитях поперечно-полосатых мышц позвоночных презентация
- Энергетика мышечного сокращения. Ферменты в толстых нитях поперечно-полосатых мышц позвоночных

Содержание
- 2. 1. Что такое электромеханическое сопряжение (ЭМС)? 2. Какая концентрация ионов кальция в покоящихся мышечных клетках? Вопросы
- 3. 7. Какова длительность ПД в скелетных мышцах, сердечной и гладких мышцах? 8. Что такое EF-hand белки?
- 4. Энергетика сокращения Источником энергии для сокращения и расслабления служит АТФ. На головках миозина есть каталитические центры,
- 5. Запасы АТФ в мышечном волокне ограничены, они обеспечивают выполнение физической нагрузки не более 1-2 с. При
- 6. 1. Фосфагенная энергосистема – первый энергетический резерв мышечного волокна. К фосфагенам относятся АТФ и КрФ (креатинфосфат).
- 7. Фосфагенная система обеспечивает мышечные усилия «взрывного» характера (спринтерский бег, прыжки, метание, подъем штанги и т.д.). Емкость
- 8. 2. гликолитическая энергосистема. В основе ее лежит расщепление анаэробно глюкозы или гликогена до молочной кислоты (выход
- 9. Высокий уровень молочной кислоты и несостоятельность щелочного резерва крови является ведущим звеном в периферических механизмах утомления:
- 10. Реакции, происходящие с участием кислорода, получили название аэробных. Окисление – аэробный путь ресинтеза АТФ протекает в
- 11. При окислении 1 молекулы глюкозы до конечных продуктов (воды и углекислого газа) ресинтезируется 36 молекул АТФ,
- 12. Пируваты (соли пировиноградной кислоты) — важные химические соединения в биохимии. Они представляют собой конечный продукт метаболизма
- 13. Электроно-транспортная цепь митохондрий является местом проведения окислительного фосфорилирования у эукариот. NADH и сукцинат, образовавшиеся в ходе
- 14. После смерти содержание АТФ в клетках быстро снижается и когда становится ниже критического, поперечные мостики миозина
- 15. Ферменты в толстых нитях поперечно-полосатых мышц позвоночных Известно, что толстая (миозиновая) нить поперечно-полосатых мышц позвоночных помимо
- 16. Полосы минорных белков в А-диске Эти полосы формируют миозин-связанные белки: С-белок, Х-белок (MyBPC), Н-белок
- 17. ЭМ исследование формы и размеров молекул С-белка Подлубная, 1990.
- 18. Х-белок в молекулярной форме в растворе, содержащем 0.3М KCI, 10 мМ K-фосфат, pH 7.0. Электронные микрофотографии
- 19. Линейные агрегаты Х-белка скелетных мышц кролика (А) и суслика (Б) в растворе 0.1 М KCl, 10
- 20. Влияние С-белка на структуру миозиновых нитей Контрольные нити миозина Подлубная, 1990.
- 21. Ферменты, связанные с толстыми нитями Как оказалось, толстые нити - это биологическая подложка для ферментативных систем,
- 22. Первый фермент – Креатинкиназа Фермент, катализирующий реакцию переноса фосфорильного остатка с креатинфосфата на ADP, в результате
- 23. Локализация креатинкиназы: в цитоплазме и М-линии саркомера. предполагается, что фермент может связываться по всей длине толстых
- 24. Функции креатинкиназы: Структурная (в М-линии) Ферментативная, заключающаяся в обеспечении быстрого ресинтеза АТФ. Активация креатинкиназы начинается после
- 25. Второй фермент – АМФ-дезаминаза (тоже ambiquitous фермент) Катализирует реакцию дезаминирования (процесс удаления аминогрупп NH2) адениловой кислоты
- 26. Функциональное значение АМФ-дезаминазы не ясно. Возможно этот фермент участвует в цикле пуриновых нуклеотидов (АТФ, АДФ, АМФ,
- 27. Третий фермент – фосфофруктокиназа или F-белок (тоже ambiquitous фермент). Фермент, катализирующий центральную реакцию гликолиза: Ф6Ф +
- 28. F-белок – фосфофруктокиназа (димер, 4нм) Где расположен в саркомере ?
- 29. F-белок связывается с миозином Подлубная, 1990.
- 30. Связывание F-белка с актином Подлубная, 1990.
- 31. Связывание F-белка в саркомере – в районе первых 2-х поперечных полос из 11-ти. Но! Возможно фермент
- 32. Итак, резюме: Значение адсорбции ферментов, участвующих в метаболизме энергии (т.е., не только трёх вышеупомянутых), на толстых
- 33. Благодарю за внимание!
- 35. Скачать презентацию

1. Что такое электромеханическое сопряжение (ЭМС)?
2. Какая концентрация ионов кальция
1. Что такое электромеханическое сопряжение (ЭМС)?
2. Какая концентрация ионов кальция

7. Какова длительность ПД в скелетных мышцах,
сердечной и гладких мышцах?
8.
7. Какова длительность ПД в скелетных мышцах,
сердечной и гладких мышцах?
8.

Энергетика сокращения
Источником энергии для сокращения и расслабления служит АТФ.
На головках
Энергетика сокращения
Источником энергии для сокращения и расслабления служит АТФ.
На головках

Запасы АТФ в мышечном волокне ограничены, они обеспечивают выполнение физической нагрузки
Запасы АТФ в мышечном волокне ограничены, они обеспечивают выполнение физической нагрузки

1. Фосфагенная энергосистема – первый энергетический резерв мышечного волокна.
К фосфагенам
1. Фосфагенная энергосистема – первый энергетический резерв мышечного волокна.
К фосфагенам

Фосфагенная система обеспечивает мышечные усилия «взрывного» характера (спринтерский бег, прыжки, метание,
Фосфагенная система обеспечивает мышечные усилия «взрывного» характера (спринтерский бег, прыжки, метание,

2. гликолитическая энергосистема.
В основе ее лежит расщепление анаэробно глюкозы или гликогена
2. гликолитическая энергосистема.
В основе ее лежит расщепление анаэробно глюкозы или гликогена

Высокий уровень молочной кислоты и несостоятельность щелочного резерва крови является ведущим
Высокий уровень молочной кислоты и несостоятельность щелочного резерва крови является ведущим

Реакции, происходящие с участием кислорода, получили название аэробных.
Окисление – аэробный
Реакции, происходящие с участием кислорода, получили название аэробных.
Окисление – аэробный

При окислении 1 молекулы глюкозы до конечных продуктов (воды и углекислого
При окислении 1 молекулы глюкозы до конечных продуктов (воды и углекислого
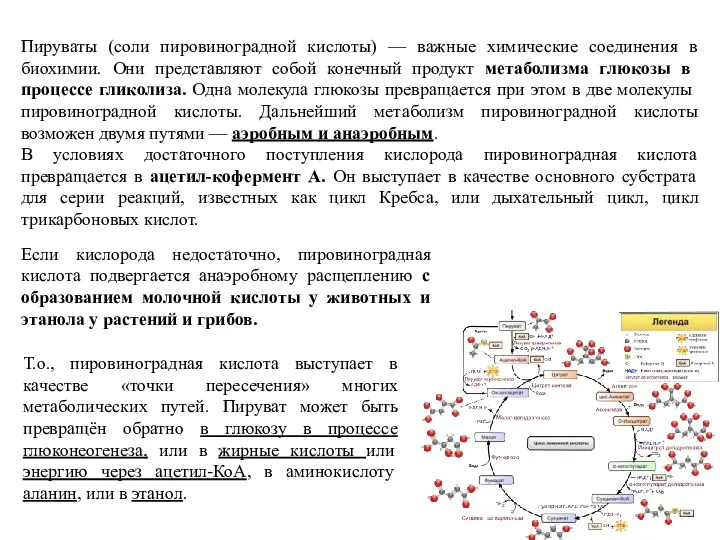
Пируваты (соли пировиноградной кислоты) — важные химические соединения в биохимии. Они
Пируваты (соли пировиноградной кислоты) — важные химические соединения в биохимии. Они
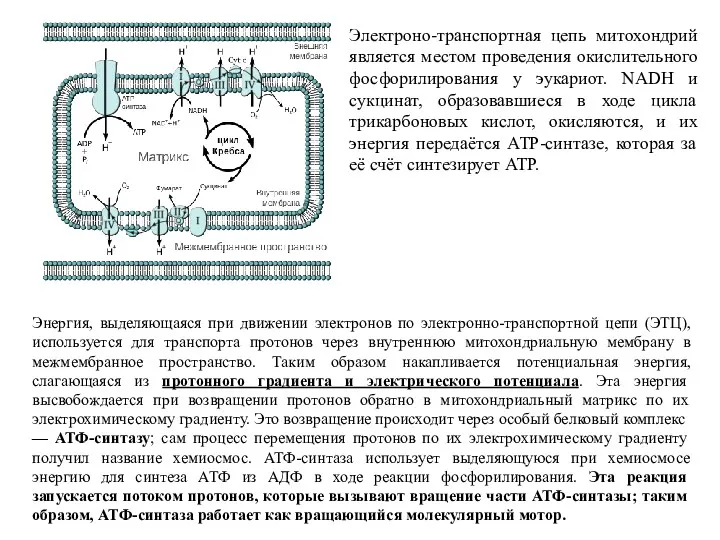
Электроно-транспортная цепь митохондрий является местом проведения окислительного фосфорилирования у эукариот. NADH
Электроно-транспортная цепь митохондрий является местом проведения окислительного фосфорилирования у эукариот. NADH

После смерти содержание АТФ в клетках быстро снижается и когда становится
После смерти содержание АТФ в клетках быстро снижается и когда становится

Ферменты в толстых нитях поперечно-полосатых
мышц позвоночных
Известно, что толстая (миозиновая)
Ферменты в толстых нитях поперечно-полосатых
мышц позвоночных
Известно, что толстая (миозиновая)
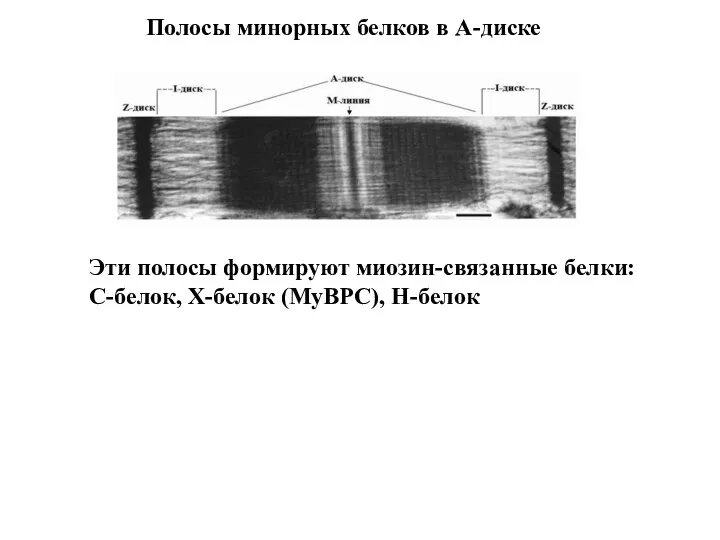
Полосы минорных белков в А-диске
Эти полосы формируют миозин-связанные белки:
С-белок, Х-белок
Полосы минорных белков в А-диске
Эти полосы формируют миозин-связанные белки:
С-белок, Х-белок

ЭМ исследование формы и размеров молекул С-белка
Подлубная, 1990.
ЭМ исследование формы и размеров молекул С-белка
Подлубная, 1990.
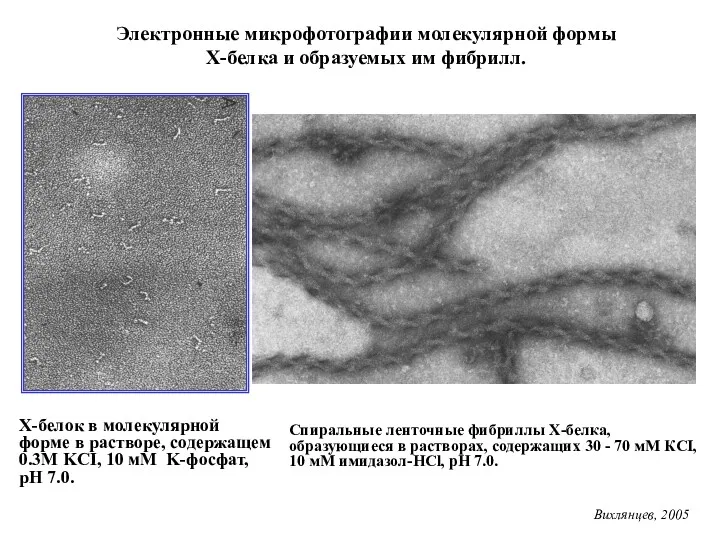
Х-белок в молекулярной форме в растворе, содержащем 0.3М KCI, 10 мМ
Х-белок в молекулярной форме в растворе, содержащем 0.3М KCI, 10 мМ
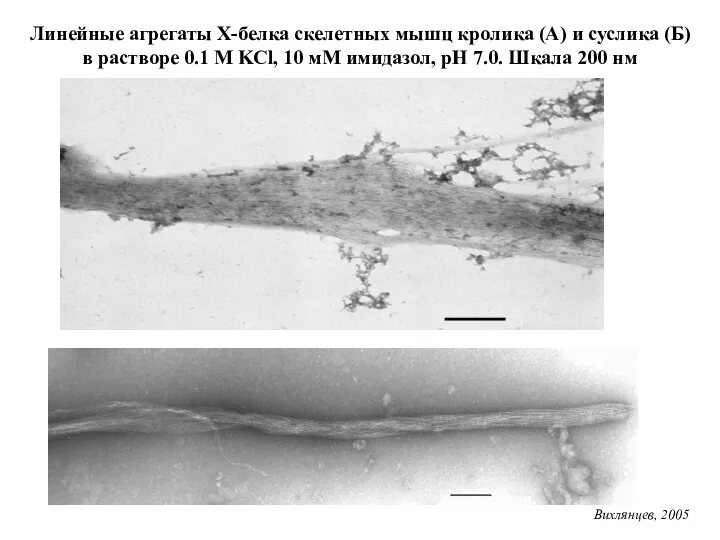
Линейные агрегаты Х-белка скелетных мышц кролика (А) и суслика (Б)
в растворе
Линейные агрегаты Х-белка скелетных мышц кролика (А) и суслика (Б)
в растворе
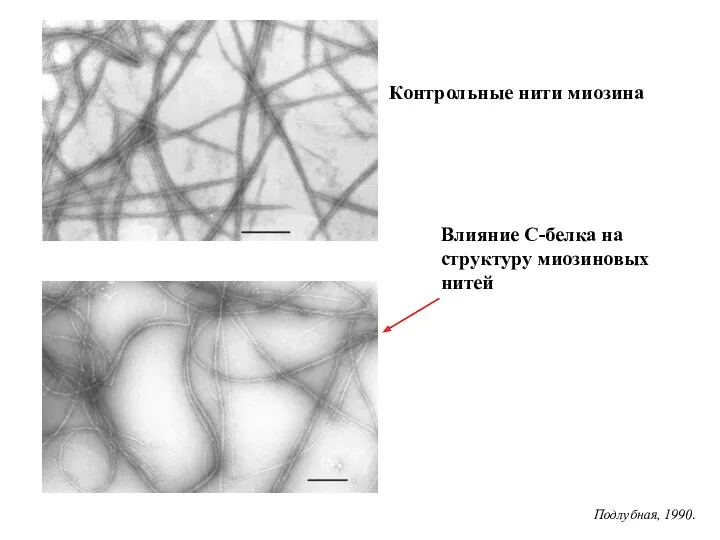
Влияние С-белка на структуру миозиновых нитей
Контрольные нити миозина
Подлубная, 1990.
Влияние С-белка на структуру миозиновых нитей
Контрольные нити миозина
Подлубная, 1990.

Ферменты, связанные с толстыми нитями
Как оказалось, толстые нити - это биологическая
Ферменты, связанные с толстыми нитями
Как оказалось, толстые нити - это биологическая

Первый фермент – Креатинкиназа
Фермент, катализирующий реакцию переноса фосфорильного остатка с
Первый фермент – Креатинкиназа
Фермент, катализирующий реакцию переноса фосфорильного остатка с

Локализация креатинкиназы: в цитоплазме и М-линии саркомера.
предполагается, что фермент может
Локализация креатинкиназы: в цитоплазме и М-линии саркомера.
предполагается, что фермент может

Функции креатинкиназы:
Структурная (в М-линии)
Ферментативная, заключающаяся в обеспечении быстрого ресинтеза АТФ.
Функции креатинкиназы:
Структурная (в М-линии)
Ферментативная, заключающаяся в обеспечении быстрого ресинтеза АТФ.

Второй фермент – АМФ-дезаминаза (тоже ambiquitous фермент)
Катализирует реакцию дезаминирования (процесс удаления
Второй фермент – АМФ-дезаминаза (тоже ambiquitous фермент)
Катализирует реакцию дезаминирования (процесс удаления

Функциональное значение АМФ-дезаминазы не ясно. Возможно этот фермент участвует в цикле
Функциональное значение АМФ-дезаминазы не ясно. Возможно этот фермент участвует в цикле

Третий фермент – фосфофруктокиназа или F-белок (тоже ambiquitous фермент).
Фермент, катализирующий центральную
Третий фермент – фосфофруктокиназа или F-белок (тоже ambiquitous фермент).
Фермент, катализирующий центральную
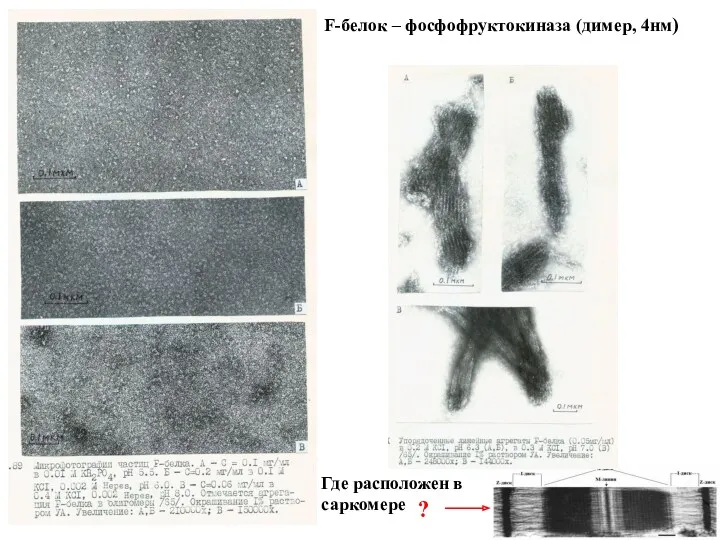
F-белок – фосфофруктокиназа (димер, 4нм)
Где расположен в саркомере
?
F-белок – фосфофруктокиназа (димер, 4нм)
Где расположен в саркомере
?
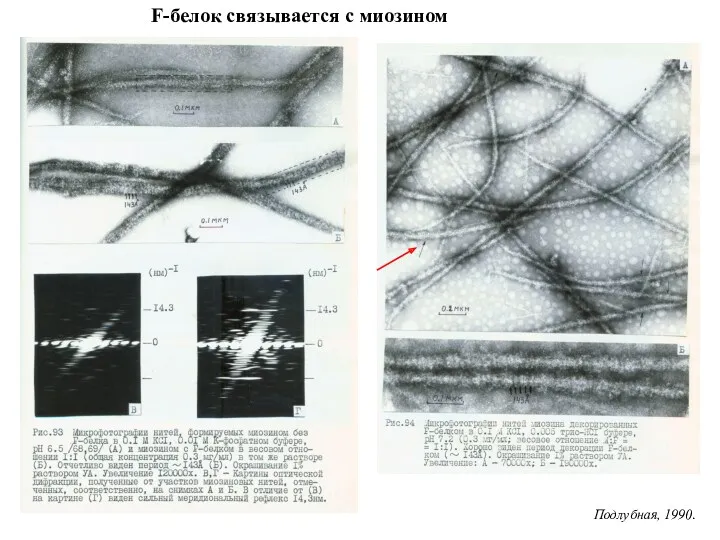
F-белок связывается с миозином
Подлубная, 1990.
F-белок связывается с миозином
Подлубная, 1990.
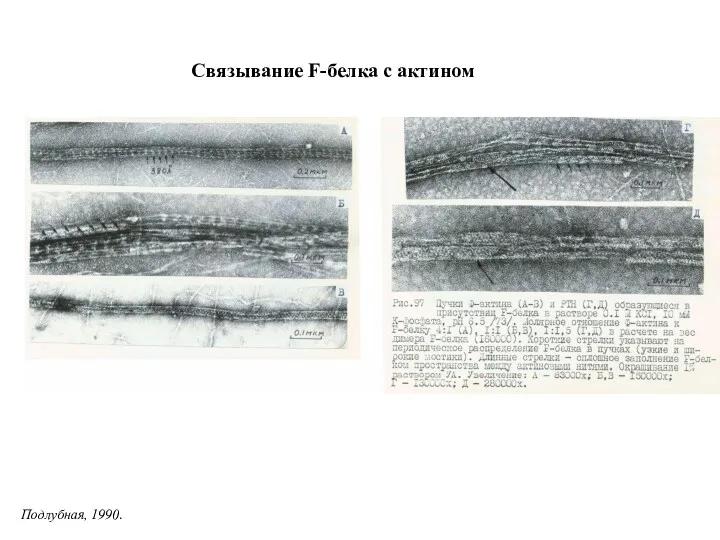
Связывание F-белка с актином
Подлубная, 1990.
Связывание F-белка с актином
Подлубная, 1990.

Связывание F-белка в саркомере – в районе первых 2-х поперечных полос
Связывание F-белка в саркомере – в районе первых 2-х поперечных полос

Итак, резюме:
Значение адсорбции ферментов, участвующих в метаболизме энергии (т.е., не
Итак, резюме:
Значение адсорбции ферментов, участвующих в метаболизме энергии (т.е., не

Благодарю
за
внимание!
Благодарю
за
внимание!
 Соединительная ткань и ее виды
Соединительная ткань и ее виды Моллюски: общая характеристика типа
Моллюски: общая характеристика типа Учение об анализаторах. Орган зрения
Учение об анализаторах. Орган зрения Цветок адениум
Цветок адениум Патрин А
Патрин А Задания линии 2 и 22 по экспериментам в КИМ ЕГЭ по биологии 2022
Задания линии 2 и 22 по экспериментам в КИМ ЕГЭ по биологии 2022 Лимфатическая система
Лимфатическая система Психология и социобиология половых различий
Психология и социобиология половых различий Презентация ц.бактерии 6кл
Презентация ц.бактерии 6кл Опасное растение борщевик
Опасное растение борщевик Внутривидовые и межвидовые взаимодействия у грибов, водорослей, высших растений. (Часть 4)
Внутривидовые и межвидовые взаимодействия у грибов, водорослей, высших растений. (Часть 4) Главные направления эволюции органического мира
Главные направления эволюции органического мира Тварина Південної Америки - вікунья
Тварина Південної Америки - вікунья Презентация по биологии на тему Класс Пресмыкающиеся. Особенности внешнего и внутреннего строения. Многообразие. Происхождение
Презентация по биологии на тему Класс Пресмыкающиеся. Особенности внешнего и внутреннего строения. Многообразие. Происхождение Черепные нервы. Обонятельные нервы
Черепные нервы. Обонятельные нервы Дезоксирибонуклеиновая кислота (ДНК)
Дезоксирибонуклеиновая кислота (ДНК) Строение клетки. Ткани
Строение клетки. Ткани Красная книга Белгородской области
Красная книга Белгородской области Физиология вегетативной нервной системы
Физиология вегетативной нервной системы Види сполучних тканин
Види сполучних тканин Эволюционная теория Ч. Дарвина
Эволюционная теория Ч. Дарвина Методы изучения наследственности человека
Методы изучения наследственности человека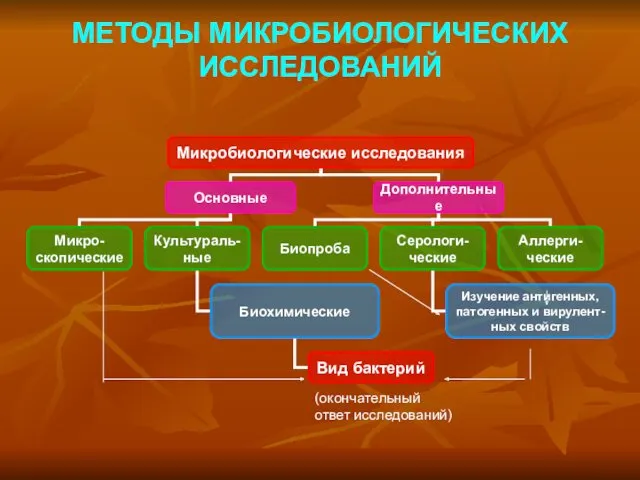 Методы микробиологических исследований
Методы микробиологических исследований Мужская половая система
Мужская половая система Действия и взаимодействия в системе Я и другой: нейробиологические аспекты
Действия и взаимодействия в системе Я и другой: нейробиологические аспекты Электрические свойства тканей организма. (Лекция 6)
Электрические свойства тканей организма. (Лекция 6) Кожа – наружный покровный орган. 8 класс
Кожа – наружный покровный орган. 8 класс В мире животных
В мире животных