- Эволюция и видообразование. Современные представления

Содержание
- 2. Главные направления эволюции (впервые сформулированы А.Н. Северцовым) Эволюция на уровне крупных таксонов называется макроэволюцией. Все современные
- 3. Главные направления эволюции
- 4. Араморфоз Это широкие анатомо-морфологические адаптации, существенно повышающие общий уровень организации рассматриваемой группы. В результате организмы получают
- 5. ПРИМЕРЫ АРОМОРФОЗОВ
- 6. Идиоадаптации - частное приспособление организмов к определённому образу жизни в конкретных условиях внешней среды. В отличие
- 7. ПРИМЕРЫ ИДИОАДАПТАЦИЙ Клювы птиц Полярная куропатка летом Полярная куропатка зимой
- 8. ПРИМЕРЫ ДЕГЕНЕРАЦИЙ Гиднора африканская Бычий и свиной цепни Омела Общая дегенерация – это изменения, которые заключаются
- 9. Соотношение направлений эволюции В природе все процессы эволюции идут непрерывно и одновременно, сочетаясь между собой и
- 10. Действие биологического прогресса и биологического регресса на популяцию Биологический прогресс Увеличение и расширение Численности особей в
- 11. Соотношения арогенезов, аллогенезов и катагенезов в ходе эволюции. Ароморфозы (арогенез) возникают в процессе эволюции сравнительно редко
- 12. Микроэволюция и видообразование Микроэволюция – это эволюция вида. В рамках учения о микроэволюции различают элементарную эволюционную
- 13. Элементарным эволюционным явлением служат мутации и рекомбинации генов. В результате этих процессов популяция становится генетически гетерогенной
- 14. Изучение закономерностей распределения генотипов в популяциях было начато А. Пирсоном (Pirson, 1904). Он показал, что при
- 15. Полное описание генетической структуры популяции вряд ли осуществимо – для этого потребовалось бы описание всех аллелей
- 16. Элементарным эволюционным материалом служат любые генотипически различные особи и группы особей т.е. мутантные организмы или носители
- 17. Элементарными факторами эволюции служит естественный отбор, мутационный процесс, популяционные волны, изоляция, дрейф генов, миграция. Естественный отбор
- 18. Формы естественного отбора
- 19. Популяционные волны или волны жизни колебание численности особей в популяции в сторону от средней численности. Популяционные
- 20. Изоляция в качестве элементарного фактора эволюции обеспечивает барьеры, исключающие репродукцию. Различают географическую, экологическую и генетическую изоляцию,
- 21. Появление новых видов Видообразование – это центральный и важнейший завершающий этап эволюции. Биологический вид – это
- 22. Механизмы видообразования Симпатрическое видообразование. Связано с расхождением групп особей одного вида и обитающих на одном ареале
- 23. Аллопатрическое видообразование. Вызывается разделением ареала вида на несколько изолированных частей. При этом на каждую такую часть
- 24. Синтетическая теория эволюции В 1930-40-е годы произошел широкий синтез генетики и дарвинизма. Генетические идеи проникли в
- 25. Дж. Хаксли Эрнст Майр Н.В. Тимофеев-Ресовский С.С. Четвериков Основоположники СТЭ
- 26. Термин «современный» или «эволюционный синтез» происходит из названия книги Дж. Хаксли «Evolution: The Modern synthesis» (1942).
- 27. Сущность синтетической теории составляет преимущественное размножение определенных генотипов и передача их потомкам. В вопросе об источнике
- 28. Основные постулаты СТЭ и её критика
- 29. Эволюционная теория прерывистого равновесия (теория пунктуализма) На рубеже 60-70-х годов ХХ века палеонтологами Элдриджем, Гулдом и
- 30. Сравнение гипотез постепенного (градуализм) и прерывистого (пунктуализм) видообразования
- 31. Теория двухкомпонентного генома В 1985 г. М.Д. Голубовский, основываясь на данных молекулярной генетики, предложил выделить в
- 32. Палеогеография Изменение геологической оболочки Земли
- 33. Морские течения и изменение климата
- 34. Экосистемная теория эволюции Основной постулат теории состоит в том, что эволюционирует не вид, а экосистема в
- 36. Скачать презентацию

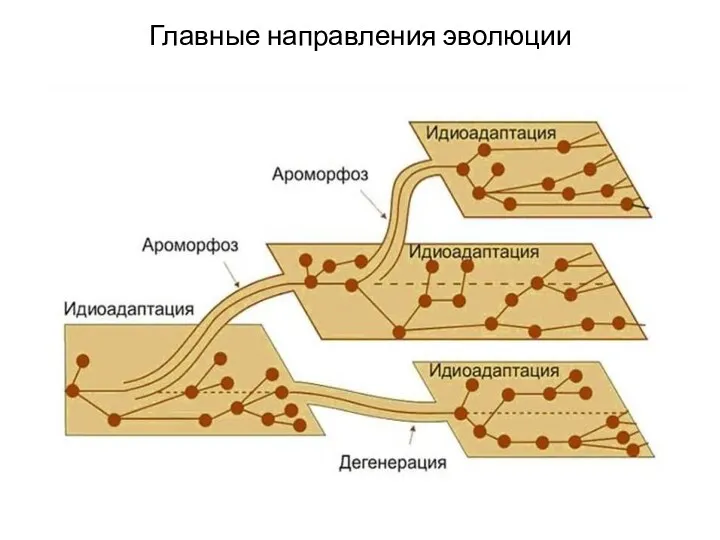
Главные направления эволюции
(впервые сформулированы А.Н. Северцовым)
Эволюция на уровне крупных таксонов называется
Главные направления эволюции
(впервые сформулированы А.Н. Северцовым)
Эволюция на уровне крупных таксонов называется
Главные направления эволюции
Главные направления эволюции

Араморфоз
Это широкие анатомо-морфологические адаптации, существенно повышающие общий уровень организации рассматриваемой группы.
Араморфоз
Это широкие анатомо-морфологические адаптации, существенно повышающие общий уровень организации рассматриваемой группы.
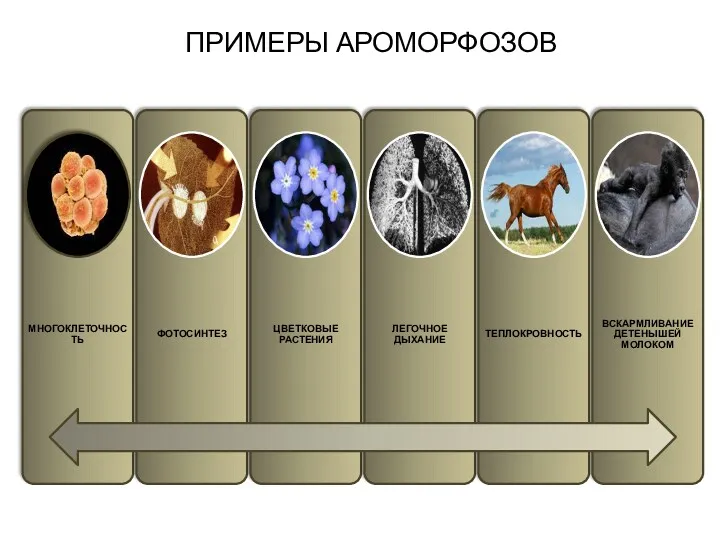
ПРИМЕРЫ АРОМОРФОЗОВ
ПРИМЕРЫ АРОМОРФОЗОВ

Идиоадаптации - частное приспособление организмов к определённому образу жизни в конкретных
Идиоадаптации - частное приспособление организмов к определённому образу жизни в конкретных

ПРИМЕРЫ ИДИОАДАПТАЦИЙ
Клювы птиц
Полярная куропатка летом
Полярная куропатка зимой
ПРИМЕРЫ ИДИОАДАПТАЦИЙ
Клювы птиц
Полярная куропатка летом
Полярная куропатка зимой

ПРИМЕРЫ ДЕГЕНЕРАЦИЙ
Гиднора африканская
Бычий и свиной цепни
Омела
Общая дегенерация – это изменения, которые
ПРИМЕРЫ ДЕГЕНЕРАЦИЙ
Гиднора африканская
Бычий и свиной цепни
Омела
Общая дегенерация – это изменения, которые

Соотношение направлений эволюции
В природе все процессы эволюции идут непрерывно и одновременно,
Соотношение направлений эволюции
В природе все процессы эволюции идут непрерывно и одновременно,
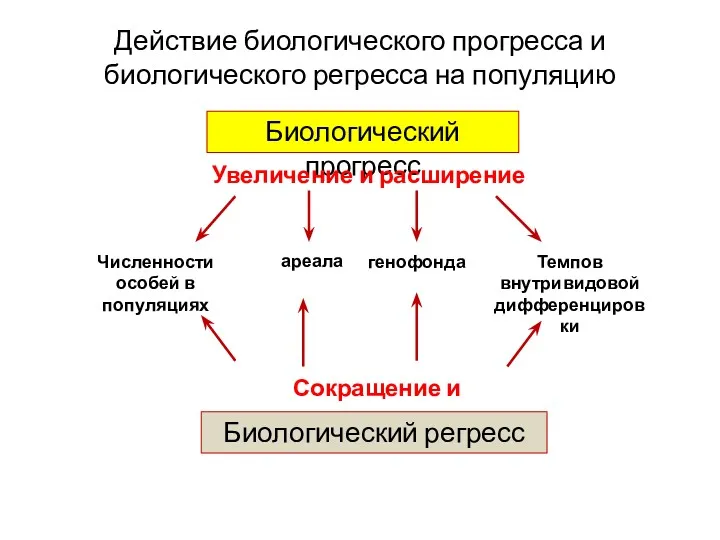
Действие биологического прогресса и биологического регресса на популяцию
Биологический прогресс
Увеличение и расширение
Численности
Действие биологического прогресса и биологического регресса на популяцию
Биологический прогресс
Увеличение и расширение
Численности
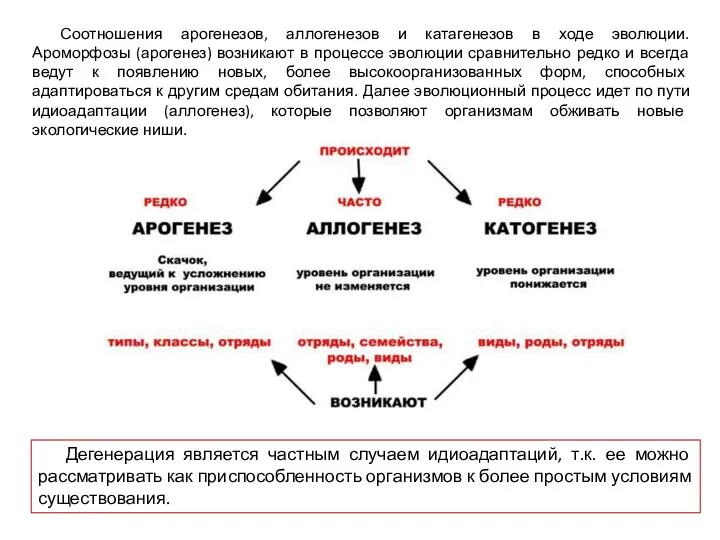
Соотношения арогенезов, аллогенезов и катагенезов в ходе эволюции.
Ароморфозы (арогенез) возникают в
Соотношения арогенезов, аллогенезов и катагенезов в ходе эволюции. Ароморфозы (арогенез) возникают в

Микроэволюция и видообразование
Микроэволюция – это эволюция вида. В рамках учения о
Микроэволюция и видообразование
Микроэволюция – это эволюция вида. В рамках учения о
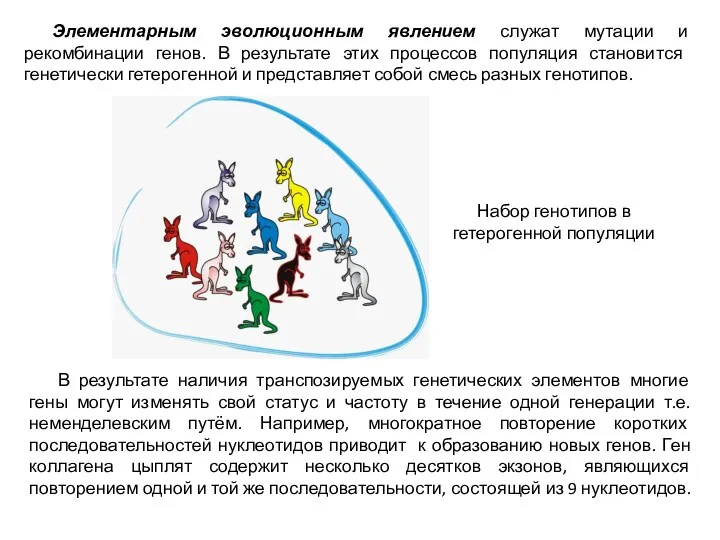
Элементарным эволюционным явлением служат мутации и рекомбинации генов. В результате этих
Элементарным эволюционным явлением служат мутации и рекомбинации генов. В результате этих

Изучение закономерностей распределения генотипов в популяциях было начато А. Пирсоном (Pirson,
Изучение закономерностей распределения генотипов в популяциях было начато А. Пирсоном (Pirson,

Полное описание генетической структуры популяции вряд ли осуществимо – для этого
Полное описание генетической структуры популяции вряд ли осуществимо – для этого
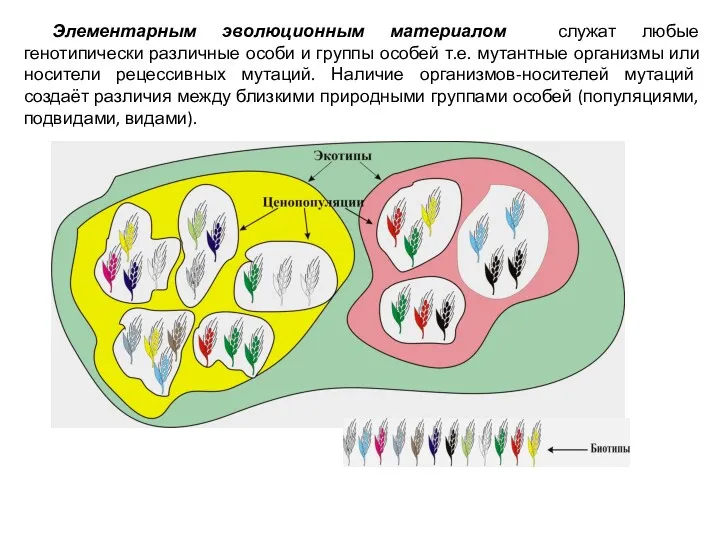
Элементарным эволюционным материалом служат любые генотипически различные особи и группы особей
Элементарным эволюционным материалом служат любые генотипически различные особи и группы особей
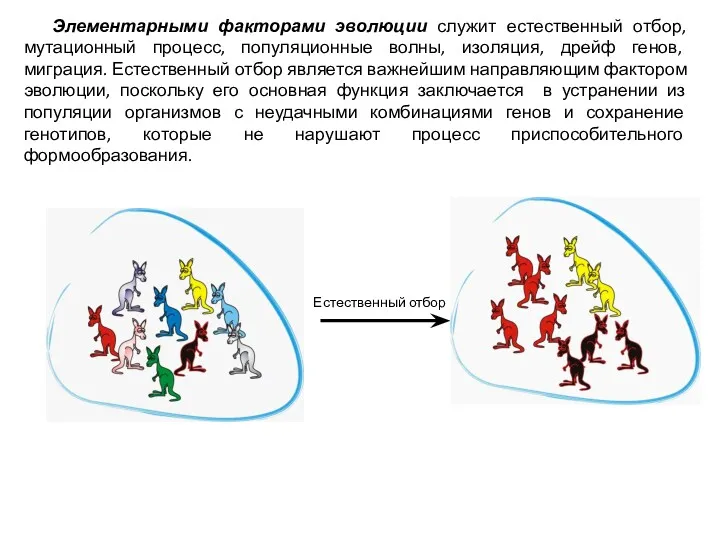
Элементарными факторами эволюции служит естественный отбор, мутационный процесс, популяционные волны, изоляция,
Элементарными факторами эволюции служит естественный отбор, мутационный процесс, популяционные волны, изоляция,

Формы естественного отбора
Формы естественного отбора
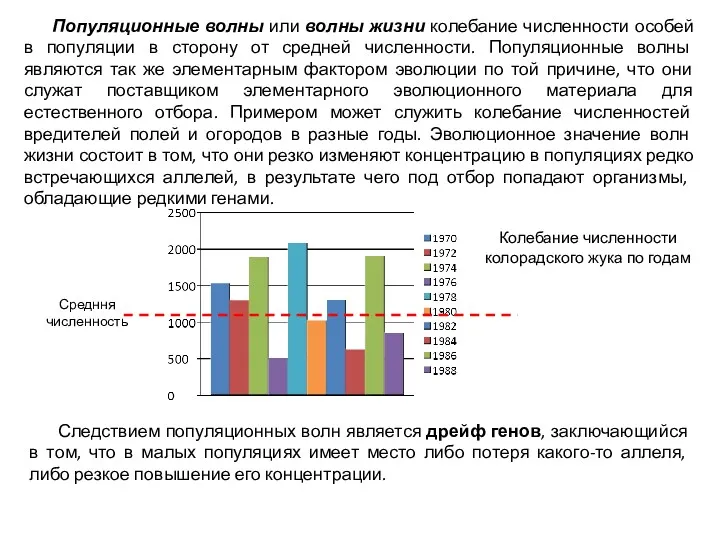
Популяционные волны или волны жизни колебание численности особей в популяции в
Популяционные волны или волны жизни колебание численности особей в популяции в

Изоляция в качестве элементарного фактора эволюции обеспечивает барьеры, исключающие репродукцию. Различают
Изоляция в качестве элементарного фактора эволюции обеспечивает барьеры, исключающие репродукцию. Различают
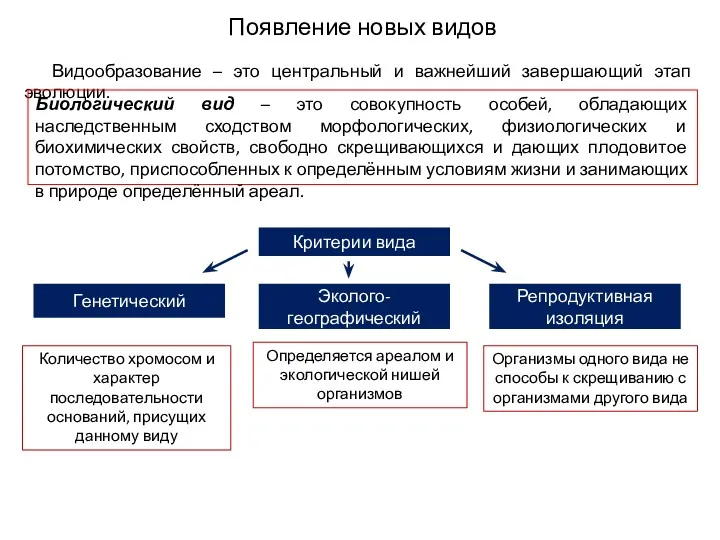
Появление новых видов
Видообразование – это центральный и важнейший завершающий этап эволюции.
Биологический
Появление новых видов
Видообразование – это центральный и важнейший завершающий этап эволюции.
Биологический
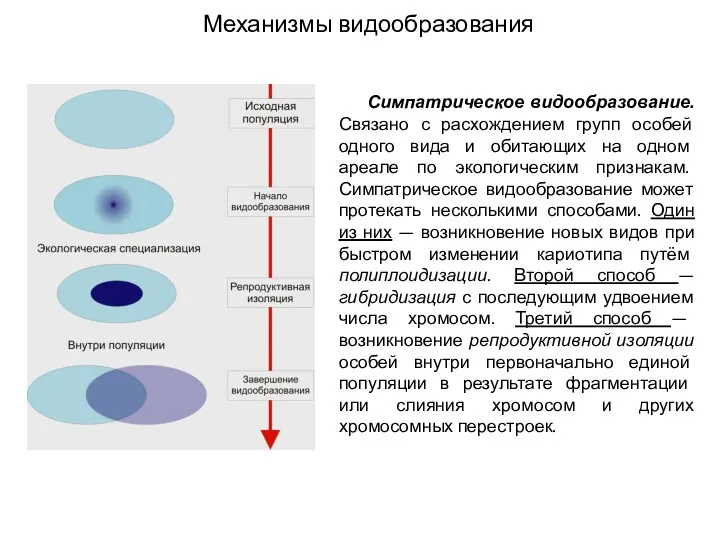
Механизмы видообразования
Симпатрическое видообразование. Связано с расхождением групп особей одного вида и
Механизмы видообразования
Симпатрическое видообразование. Связано с расхождением групп особей одного вида и
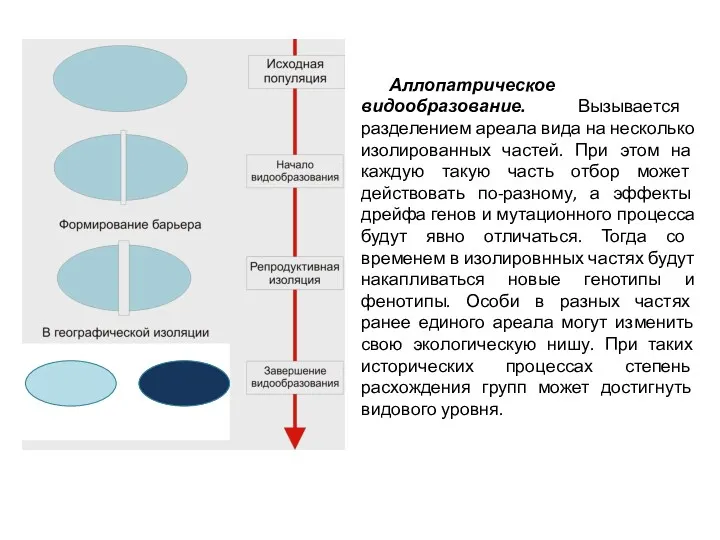
Аллопатрическое видообразование. Вызывается разделением ареала вида на несколько изолированных частей. При
Аллопатрическое видообразование. Вызывается разделением ареала вида на несколько изолированных частей. При

Синтетическая теория эволюции
В 1930-40-е годы произошел широкий синтез генетики и дарвинизма.
Синтетическая теория эволюции
В 1930-40-е годы произошел широкий синтез генетики и дарвинизма.

Дж. Хаксли
Эрнст Майр
Н.В. Тимофеев-Ресовский
С.С. Четвериков
Основоположники СТЭ
Дж. Хаксли
Эрнст Майр
Н.В. Тимофеев-Ресовский
С.С. Четвериков
Основоположники СТЭ

Термин «современный» или «эволюционный синтез» происходит из названия книги Дж. Хаксли
Термин «современный» или «эволюционный синтез» происходит из названия книги Дж. Хаксли

Сущность синтетической теории составляет преимущественное размножение определенных генотипов и передача их
Сущность синтетической теории составляет преимущественное размножение определенных генотипов и передача их

Основные постулаты СТЭ и её критика
Основные постулаты СТЭ и её критика

Эволюционная теория прерывистого равновесия
(теория пунктуализма)
На рубеже 60-70-х годов ХХ века палеонтологами
Эволюционная теория прерывистого равновесия
(теория пунктуализма)
На рубеже 60-70-х годов ХХ века палеонтологами

Сравнение гипотез постепенного (градуализм) и прерывистого (пунктуализм) видообразования
Сравнение гипотез постепенного (градуализм) и прерывистого (пунктуализм) видообразования
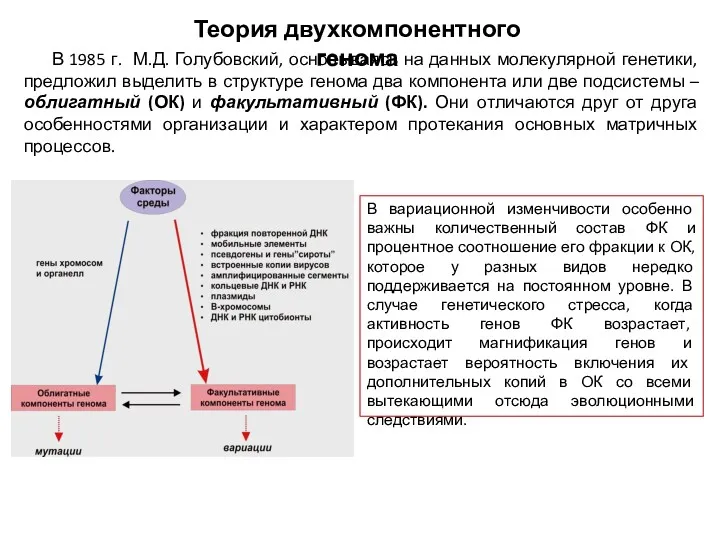
Теория двухкомпонентного генома
В 1985 г. М.Д. Голубовский, основываясь на данных молекулярной
Теория двухкомпонентного генома
В 1985 г. М.Д. Голубовский, основываясь на данных молекулярной

Палеогеография
Изменение геологической оболочки Земли
Палеогеография
Изменение геологической оболочки Земли
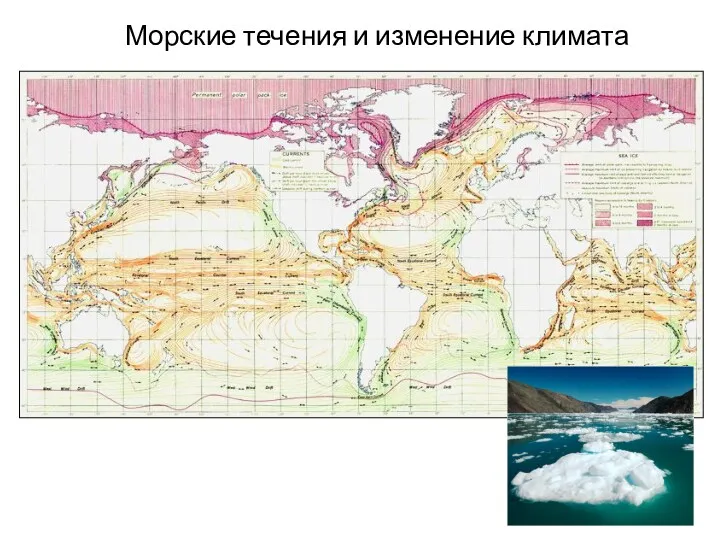
Морские течения и изменение климата
Морские течения и изменение климата
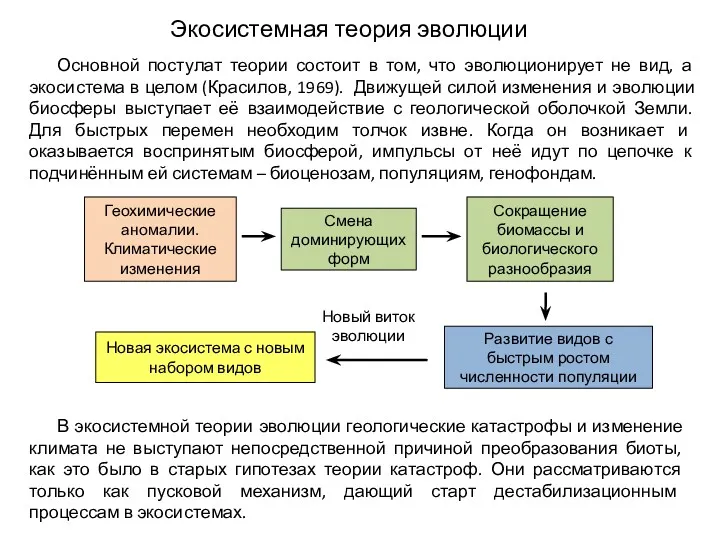
Экосистемная теория эволюции
Основной постулат теории состоит в том, что эволюционирует не
Экосистемная теория эволюции
Основной постулат теории состоит в том, что эволюционирует не
 Мужской ликбез. Все об уходе за внешностью
Мужской ликбез. Все об уходе за внешностью Ядро
Ядро Вода - основной источник жизни и здоровья
Вода - основной источник жизни и здоровья Встечаем пернатых друзей
Встечаем пернатых друзей Общие сведения о клетке. Патология клетки
Общие сведения о клетке. Патология клетки В сосновом лесу
В сосновом лесу урок биологии в 7 классе Внешнее и внутреннее строение насекомых
урок биологии в 7 классе Внешнее и внутреннее строение насекомых Систематика растений
Систематика растений Биологическое разнообразие грибов. Их практическое значение
Биологическое разнообразие грибов. Их практическое значение Вопросы по биологии
Вопросы по биологии Формы естественного отбора
Формы естественного отбора In vitro жағдайындағы өсірілетін клеткаларды биотехнологияда қолдану
In vitro жағдайындағы өсірілетін клеткаларды биотехнологияда қолдану Антропогенез. Расы. Расизм. Часть 2
Антропогенез. Расы. Расизм. Часть 2 Вестибулярная сенсорная система
Вестибулярная сенсорная система Строение и работа сердца
Строение и работа сердца Механизмы эволюционного процесса. Главные направления эволюции
Механизмы эволюционного процесса. Главные направления эволюции Лекарственные и съедобные растения,произрастающие в Якутии
Лекарственные и съедобные растения,произрастающие в Якутии Физиологические механизмы автоматии сердца. Микроциркуляция. Регуляция периферического кровообращения
Физиологические механизмы автоматии сердца. Микроциркуляция. Регуляция периферического кровообращения Деление клетки. Митоз
Деление клетки. Митоз Эволюция кровеносной системы животных
Эволюция кровеносной системы животных Семейство Кошачьи
Семейство Кошачьи День птиц. Конкурсная программа
День птиц. Конкурсная программа Презентация по биологии Мхи, папоротники, хвощи и плауны для 6 класса.
Презентация по биологии Мхи, папоротники, хвощи и плауны для 6 класса. Обитатели солёных водоёмов
Обитатели солёных водоёмов Типы развития насекомых. Насекомые с неполным превращением
Типы развития насекомых. Насекомые с неполным превращением Основы разведения животных
Основы разведения животных Жасушалардың қартаюы және сомалық жасушалардың жасушалық өлу түрлер
Жасушалардың қартаюы және сомалық жасушалардың жасушалық өлу түрлер Extinction of animals
Extinction of animals