- Факторы патогенности микроорганизмов

Содержание
- 2. «инфекция» (заражение) совокупность биологических процессов, происходящих в макроорганизме при внедрении в него патогенных микроорганизмов, независимо от
- 3. Инфекция Инфекционные болезни рассматривают как явления, включающие биологический и социальный факторы. Так, механизмы передачи инфекционных болезней,
- 4. Инфекция Отличия от других заболеваний Заразительность (контагиозность) Цикличность (периоды) Развитие противоинфекционного иммунитета Инкубационный период
- 5. Патогенные микроорганизмы Характерными свойствами патогенных микроорганизмов являются специфичность (способность вызывать определённую инфекционную болезнь после проникновения в
- 6. Место проникновения возбудителя называется входными воротами. Как правило это -ткани, лишенные физиологической защиты против конкретного вида
- 7. Инфицирующая доза возбудителя Инфицирующая доза возбудителя – минимальное количество микробных клеток, способных вызвать инфекционный процесс. Величина
- 8. Инфицирующая доза Для высоковирулентного возбудителя Yersinia pestis (чума) достаточноао несколько бактериальных клеток. Shigella dysenteriae – десятки
- 9. Периоды инфекционного заболевания 1 период - Инкубационный- от момента заражения до проявления клинических симптомов Локализация возбудителя
- 10. Периоды инфекционного заболевания 4-й период - Исход заболевания (outcome) - Реконвалесценция Переход в хроническую форму Формирование
- 11. Периоды инфекционного заболевания 2-й период - Продромальный (prodrome)-это проявление “общих симптомов”- дискомфорт, усталость, озноб. Клинически -
- 12. В настоящее время наблюдается переход от традиционного представления о бактериях как строго одноклеточных организмах к представлению
- 13. Пути проникновения возбудителя в макроорганизм
- 14. Факторы патогенности микроорганизмов Факторы адгезии и колонизации Факторы инвазии Антифагоцитарные факторы Факторы, нарушающие иммунную защиту Токсические
- 15. Адгезия происходит на поверхности слизистых оболочек различных органов и систем. Адгезия начинается как обратимый процесс, затем
- 16. Жгутики способствуют приближению к поверхности клетки
- 17. Холерные вибрионы
- 18. Адгезия. На клетке хозяина имеются рецепторы - разнообразные молекулы (гликолипиды, маннозные остатки, протеогликаны). Рецепторами для адгезинов
- 19. Факторы адгезии Адгезия завершается лиганд-рецепторным взаимодействием. Это высокоспецифичный процесс При котором адгезины комплементарны рецепторам клетки. Со
- 20. Колонизация эпителия трахеи Bordetella pertussis (клетки без ресничек свободны от бактерий)
- 21. Факторы адгезии У грамотрицательных бактерий функцию распознавания и прикрепления бактерий чаще осуществляют пили или фимбрии. Они
- 22. Фимбрии у гонококков. Количество 100-500. Состоят из пилина. У грамотрицательных бактерий факторами адгезии служат фимбрии (фимбриальные
- 23. (А)Электронная микрофотография негативно контрастированных E coli. Показаны извитые жгутики и многочисленные короткие тонкие и более ригидные
- 24. Пили E.coli
- 25. Адгезины Афимбриальные адгезины – филаментозный гемаглютинин у Bordetella pertussis, ответственный за прикрепление к реснитчатому эпителию дыхательных
- 26. Адгезия Колонизация эпителия трахеи Bordetella pertussis (клетки без ресничек свободны от бактерий)
- 27. Факторы адгезии у грамположительных бактерий Тейхоевые и липотейхоевые кислоты, наружные белки клеточной стенки
- 28. Figure 2-9. Структура тейхоевых кислот (A) Рибитол тейхоевая кислота с повторяющимися фрагментами связанными 1,5-фосфодиэфирными связями D-рибитола
- 29. Адгезия У грамположительных бактерий – Тейхоевые и липотейхоевые кислоты. Фибронектин связывающие белки ( стафилококки, стрептококки). М-протеин
- 30. Streptococcus pyogenes. Cell surface fibrils
- 31. M protein and fimbriae of Group A streptococci –адгезия и защита от фагоцитоза
- 32. Уропатогенные эшерихии экспрессируют два вида ворсинок: Р-ворсинки и ворсинки I типа, связываются с разными рецепторами Адгезия
- 33. Инвазия-распространения микроорганизмов в межклеточных пространствах тканей организма хозяина и проникновения их внутрь его клеток.
- 34. Инвазия При инвазии рецепторами эукариотических клеток являются их мембранные молекулы, основная функция которых - межклеточные взаимодействия.
- 35. Инвазия Адгезия - сигнал к синтезу белков (IpaB, IpaC и IpaD), выполняющих функции инвазинов. Их транспорт
- 36. Yersinia spp., Salmonella spp. и Shigella spp. осуществляют инвазию кишечного эпителия, основными "воротами" являются М-клетки. Одной
- 37. Инвазия Шигеллы мигрирует в подслизистый слой, в область лимфоидных фолликулов, где подвергается фагоцитозу мононуклеарными фагоцитами. Шигеллы
- 38. Механизм бактериальной инвазии у некоторых Грам-отрицательных бактерий
- 39. (D)Сканирующая электронная микрофотография энтеропатогенных Е. coli, прикрепляющихся к опороподобным клеточным выростам на поверхности HeLa клеток. (E)
- 40. С образования биопленок начинается развитие любой инфекции. Биопленки -тонкий слой микроорганизмов с секретированными ими полимерами, который
- 41. Биопленки Это основной фенотип почти всех бактерий в естественных условиях обитания, как во внешней среде, так
- 42. Биопленки Существование бактерий в виде биопленок усиливает свою защиту от фагоцитоза, ультрафиолетового излучения, вирусов и дегидратации,
- 43. Адгезия микроорганизмов к имплантируемым устройствам. Ни один из используемых для создания имплантируемых устройств материалов не является
- 44. Формирование биопленок Колонизация (объекты окружающей среды, клапаны сердца, зубная эмаль и другое, катетеры,….) Резистентость фагоцитозу Резистентость
- 45. Факторы инвазии Инвазия –проникновение возбудителя через слизистые и соединительнотканные барьеры Агрессия – подавление естественной резистентности и
- 46. Факторы инвазии Инвазия-распространения микроорганизмов в межклеточных пространствах тканей организма хозяина и проникновения их внутрь его клеток.
- 47. Факторы инвазии Гиалуронидаза – деполимеризует гиалуроновую кислоту, высокополимерное соединение, состоящий из остатков N - ацетилглюкозамина и
- 48. Факторы инвазии Нейраминидаза- гидролизует гликозидные связи в гликопротеидах, ганглиозидах, отщепляет от них остатки сиаловых (нейраминовых кислот),
- 49. Факторы инвазии и агрессии Лецитиназа – гидролизует лецитин (фосфоглицерид фосфатидилхолин) основной компонент мембран млекопитающих, разрушает липиды
- 50. Лецитиназная активность
- 51. Протеолитические ферменты. Основной целью протеолитических ферментов, образуемых бактериями, являются сигнальные и эффекторные молекулы иммунной защиты Коагулаза
- 52. IgA протеазы – гидролиз секреторных иммуноглобулинов Neisseria meningitidis сериновая протеаза Haemophilus spp. сериновая протеаза Streptococcus spp.
- 53. Ферменты. ДНК-аза – гидролиз молекул ДНК, разрыв фосфодиэфирных связей распад ДНК и РНК молекул на олигонуклеотиды
- 54. Тест на ДНК-зу.
- 55. Тест на плазмокоагулазу
- 56. Ферменты Уреаза - распад мочевины, аммиак вызывает защелачивание среды, прямой токсический эффект. Токсичен для центральной нервной
- 57. Антифагоцитарные факторы
- 58. Стадии фагоцитоза
- 60. Антифагоцитарные факторы Имеют поверхностную локализацию – капсулы, капсулоподобные структуры Не являются жизненно важными для бактериальной клетки
- 61. Антифагоцитарные факторы Защита от фагоцитоза может происходить на различных стадиях процесса: На стадии узнавания-поглощения Капсулы, капсулоподобный
- 63. Число(фигура) 11. Негативное контрастирование Streptococcus pyogenes при электронной микроскопии (28,000X). Ореол вокруг цепочки клеток - капсула
- 64. Колонии Bacillus anthracis. Рост слизистых или мукоидных бактериальные колонии - обычно свидетельствует о продукции капсул. В
- 65. Бактериальные капсулы, контрастированные китайской тушью, рассматриваемые в световой микроскоп. Это - истинная капсула, обособленный слой полисахаридов,
- 66. Антифагоцитарные факторы Капсула –метод Бурри-Гинса
- 70. Антифагоцитарные факторы Выживание микробных клеток после поглощения фагоцитом. Препятствие слиянию фагосомы с лизосомой – корд-фактор микобактерий
- 71. Незавершенный фагоцитоз
- 72. Инвазия нефагоцитирующих клеток Активная инвазия клеток, не относящихся к фагоцитам, прежде всего эпителиальных: внутри таких клеток
- 73. Уклонение от иммунного ответа Вариабельность антигенных свойств Антигенная мимикрия Образование L-форм Экранирование антигенных детерминант с помощью
- 74. Streptococcus sp
- 75. Pseudomonas
- 76. Pseudomonas aeruginosa
- 77. Бактериальные токсины Оказывают непосредственное патологическое действие Экзотоксины (белковые токсины)– выделяются преимущественно в окружающую среду. Эндотоксины- связаны
- 78. Бактериальные токсины Характерные свойства белковых токсинов Токсичность Специфичность Термолабильность Иммуногенны-образуют анатоксины
- 79. Бактериальные токсины Простые – полипептидная цепь Сложные – несколько связанных полипептидных цепей, соединенных между собой. Простые
- 80. Классификация по механизму действия Ингибируют синтез белка- цитотоксины Повреждают клеточные мембраны-мембранотоксины Нарушают передачу сигналов – функциональные
- 81. Механизм действия токсинов Нарушающие синтез белка Дифтерийный токсин –простой. Обладает Рибозил-трансферазной активностью, переносит ADF-рибозу На мишень
- 83. Токсины, нарушающие синтез белка Шига-токсин – Субъединица А, обладающая ферментативной активностью, действует как N-гликозидаза, отщепляя единичный
- 84. Порообразующие токсины. Бактериальные токсины, функционирующие посредством вставки в плазматическую мембрану хозяина и формирующие в ней трансмембранные
- 85. Токсины, повреждающие клеточные мембраны. Порообразующие-гемолизины и лейкоцидин. Могут повреждать моноциты, тромбоциты. Альфа токсин стафилококков Нарушающие целостность
- 86. Типы гемолиза на кровяном агаре
- 88. β-гемолитические стрептококки группы А (Streptococcus pyogenes)
- 89. Функциональные блокаторы (активаторы путей метаболизма вторичных мессенджеров Нарушающие функцию аданилатциклазы – Холерный токсин –сложный токсин, состоит
- 90. Холерный токсин
- 91. Нейротоксины C.botulinum (BoNT серотипов A vG) и C.tetani -протеазы Нейротоксины синтезируются в виде неактивных полипептидов с
- 92. Клеточные мишени -группа белков, необходимых для соединения синаптических пузырьков с пресинаптическими плазматическими мембранами с последующим высвобождением
- 93. Нейротоксин Столбнячный токсин поражает два вида нейронов. Он связывается с рецепторами пресинаптической мембраны моторных нейронов, затем
- 94. Протеолитические токсины нейротоксины Обладает протеазной активностью, разрушает белок синаптобревин, блокирует систему торможения –судороги Ботулотоксин – действует
- 95. Токсины-суперантигены, активаторы иммунного ответа Иммуностимулирующий потенциал токсинов является следствием их способности связывать различные участки белков главного
- 96. Токсины-суперантигены
- 97. Эндотоксин Сложный липополисахаридный комплекс, содержится в клеточной стенке грамотрицательных бактерий и выделяется в окружающую среду при
- 98. Эндотоксины Липид А Центральный олигосахарид О-антиген
- 100. Скачать презентацию

«инфекция» (заражение)
совокупность биологических процессов, происходящих в макроорганизме при внедрении в
«инфекция» (заражение)
совокупность биологических процессов, происходящих в макроорганизме при внедрении в

Инфекция
Инфекционные болезни рассматривают как явления, включающие биологический и социальный факторы. Так,
Инфекция
Инфекционные болезни рассматривают как явления, включающие биологический и социальный факторы. Так,

Инфекция
Отличия от других заболеваний
Заразительность (контагиозность)
Цикличность (периоды)
Развитие противоинфекционного
Инфекция
Отличия от других заболеваний
Заразительность (контагиозность)
Цикличность (периоды)
Развитие противоинфекционного

Патогенные микроорганизмы
Характерными свойствами патогенных микроорганизмов являются специфичность (способность вызывать определённую инфекционную
Патогенные микроорганизмы
Характерными свойствами патогенных микроорганизмов являются специфичность (способность вызывать определённую инфекционную

Место проникновения возбудителя называется входными воротами.
Как правило это -ткани, лишенные физиологической
Место проникновения возбудителя называется входными воротами.
Как правило это -ткани, лишенные физиологической

Инфицирующая доза возбудителя
Инфицирующая доза возбудителя – минимальное количество микробных клеток, способных
Инфицирующая доза возбудителя
Инфицирующая доза возбудителя – минимальное количество микробных клеток, способных

Инфицирующая доза
Для высоковирулентного возбудителя Yersinia pestis (чума) достаточноао несколько бактериальных клеток.
Инфицирующая доза
Для высоковирулентного возбудителя Yersinia pestis (чума) достаточноао несколько бактериальных клеток.

Периоды инфекционного заболевания
1 период - Инкубационный- от момента заражения до проявления
Периоды инфекционного заболевания
1 период - Инкубационный- от момента заражения до проявления

Периоды инфекционного заболевания
4-й период - Исход заболевания (outcome) -
Реконвалесценция
Переход в
Периоды инфекционного заболевания
4-й период - Исход заболевания (outcome) -
Реконвалесценция
Переход в

Периоды инфекционного заболевания
2-й период - Продромальный (prodrome)-это проявление “общих симптомов”- дискомфорт,
Периоды инфекционного заболевания
2-й период - Продромальный (prodrome)-это проявление “общих симптомов”- дискомфорт,

В настоящее время наблюдается переход от традиционного представления о бактериях как
В настоящее время наблюдается переход от традиционного представления о бактериях как

Пути проникновения возбудителя в макроорганизм
Пути проникновения возбудителя в макроорганизм

Факторы патогенности микроорганизмов
Факторы адгезии и колонизации
Факторы инвазии
Антифагоцитарные факторы
Факторы, нарушающие иммунную защиту
Токсические
Факторы патогенности микроорганизмов
Факторы адгезии и колонизации
Факторы инвазии
Антифагоцитарные факторы
Факторы, нарушающие иммунную защиту
Токсические

Адгезия происходит на поверхности слизистых оболочек различных органов и систем.
Адгезия начинается
Адгезия происходит на поверхности слизистых оболочек различных органов и систем.
Адгезия начинается
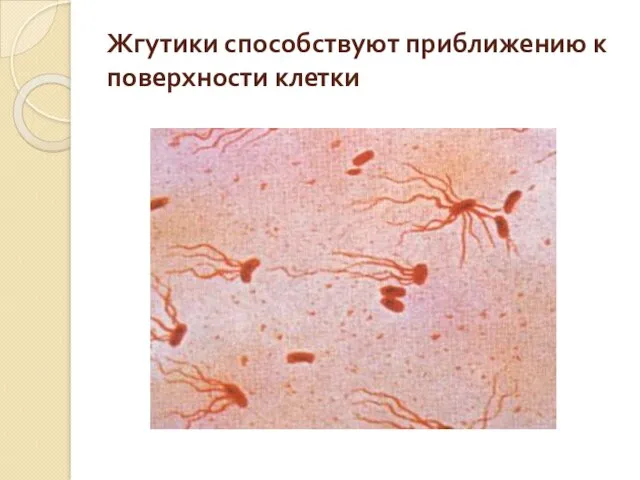
Жгутики способствуют приближению к поверхности клетки
Жгутики способствуют приближению к поверхности клетки

Холерные вибрионы
Холерные вибрионы

Адгезия.
На клетке хозяина имеются рецепторы - разнообразные молекулы (гликолипиды, маннозные остатки,
Адгезия.
На клетке хозяина имеются рецепторы - разнообразные молекулы (гликолипиды, маннозные остатки,

Факторы адгезии
Адгезия завершается лиганд-рецепторным взаимодействием. Это высокоспецифичный процесс
При котором адгезины комплементарны
Факторы адгезии
Адгезия завершается лиганд-рецепторным взаимодействием. Это высокоспецифичный процесс
При котором адгезины комплементарны
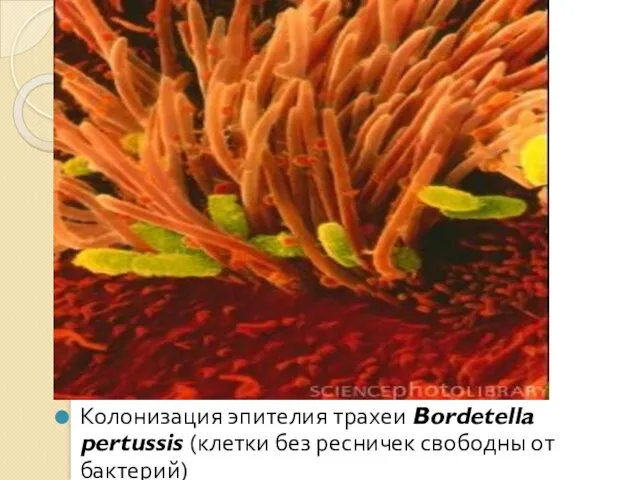
Колонизация эпителия трахеи Bordetella pertussis (клетки без ресничек свободны от бактерий)
Колонизация эпителия трахеи Bordetella pertussis (клетки без ресничек свободны от бактерий)

Факторы адгезии
У грамотрицательных бактерий функцию распознавания и прикрепления бактерий чаще осуществляют
Факторы адгезии
У грамотрицательных бактерий функцию распознавания и прикрепления бактерий чаще осуществляют

Фимбрии у гонококков. Количество 100-500. Состоят из пилина.
У грамотрицательных бактерий факторами
Фимбрии у гонококков. Количество 100-500. Состоят из пилина.
У грамотрицательных бактерий факторами
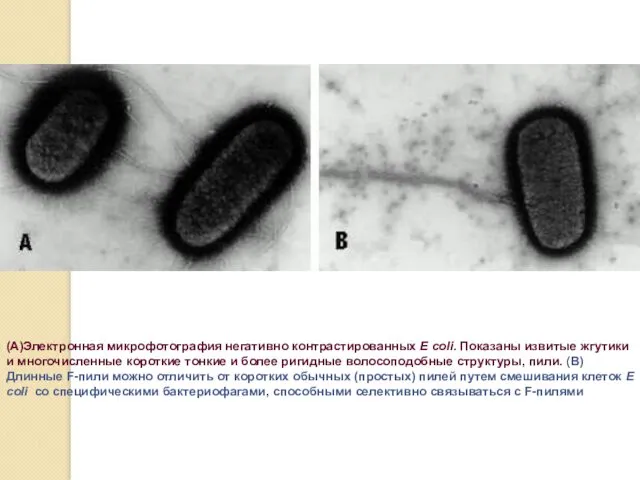
(А)Электронная микрофотография негативно контрастированных E coli. Показаны извитые жгутики и многочисленные
(А)Электронная микрофотография негативно контрастированных E coli. Показаны извитые жгутики и многочисленные
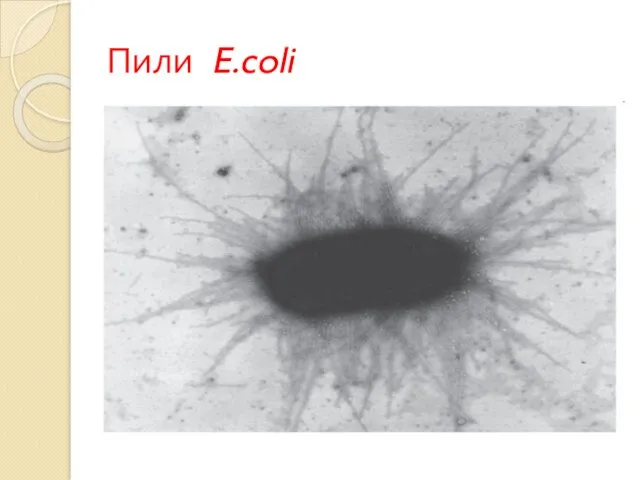
Пили E.coli
Пили E.coli

Адгезины
Афимбриальные адгезины – филаментозный гемаглютинин у Bordetella pertussis, ответственный за прикрепление
Адгезины
Афимбриальные адгезины – филаментозный гемаглютинин у Bordetella pertussis, ответственный за прикрепление

Адгезия
Колонизация эпителия трахеи Bordetella pertussis (клетки без ресничек свободны от бактерий)
Адгезия
Колонизация эпителия трахеи Bordetella pertussis (клетки без ресничек свободны от бактерий)
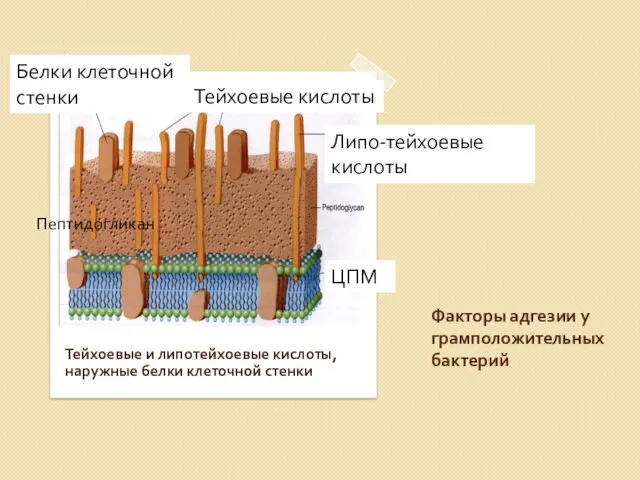
Факторы адгезии у грамположительных бактерий
Тейхоевые и липотейхоевые кислоты, наружные белки клеточной
Факторы адгезии у грамположительных бактерий
Тейхоевые и липотейхоевые кислоты, наружные белки клеточной

Figure 2-9. Структура тейхоевых кислот (A) Рибитол тейхоевая кислота с повторяющимися
Figure 2-9. Структура тейхоевых кислот (A) Рибитол тейхоевая кислота с повторяющимися

Адгезия
У грамположительных бактерий –
Тейхоевые и липотейхоевые кислоты.
Фибронектин связывающие белки ( стафилококки,
Адгезия
У грамположительных бактерий –
Тейхоевые и липотейхоевые кислоты.
Фибронектин связывающие белки ( стафилококки,

Streptococcus pyogenes.
Cell surface fibrils
Streptococcus pyogenes.
Cell surface fibrils
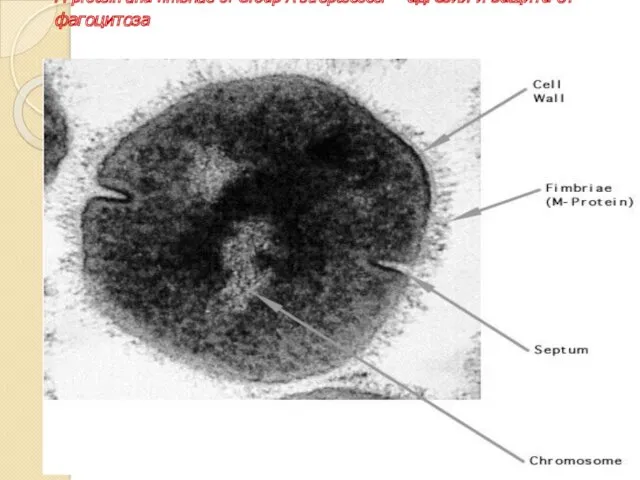
M protein and fimbriae of Group A streptococci –адгезия и защита
M protein and fimbriae of Group A streptococci –адгезия и защита

Уропатогенные эшерихии экспрессируют два вида ворсинок: Р-ворсинки и ворсинки I типа,
Уропатогенные эшерихии экспрессируют два вида ворсинок: Р-ворсинки и ворсинки I типа,

Инвазия-распространения микроорганизмов в межклеточных пространствах тканей организма хозяина и проникновения их
Инвазия-распространения микроорганизмов в межклеточных пространствах тканей организма хозяина и проникновения их

Инвазия
При инвазии рецепторами эукариотических клеток являются их мембранные молекулы, основная функция
Инвазия
При инвазии рецепторами эукариотических клеток являются их мембранные молекулы, основная функция

Инвазия
Адгезия - сигнал к синтезу белков (IpaB, IpaC и IpaD), выполняющих
Инвазия
Адгезия - сигнал к синтезу белков (IpaB, IpaC и IpaD), выполняющих

Yersinia spp., Salmonella spp. и Shigella spp. осуществляют инвазию кишечного эпителия,
Yersinia spp., Salmonella spp. и Shigella spp. осуществляют инвазию кишечного эпителия,

Инвазия
Шигеллы мигрирует в подслизистый слой, в область лимфоидных фолликулов, где подвергается
Инвазия
Шигеллы мигрирует в подслизистый слой, в область лимфоидных фолликулов, где подвергается
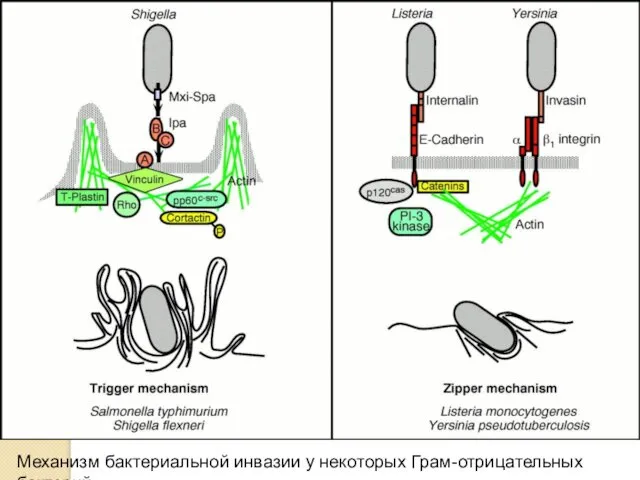
Механизм бактериальной инвазии у некоторых Грам-отрицательных бактерий
Механизм бактериальной инвазии у некоторых Грам-отрицательных бактерий
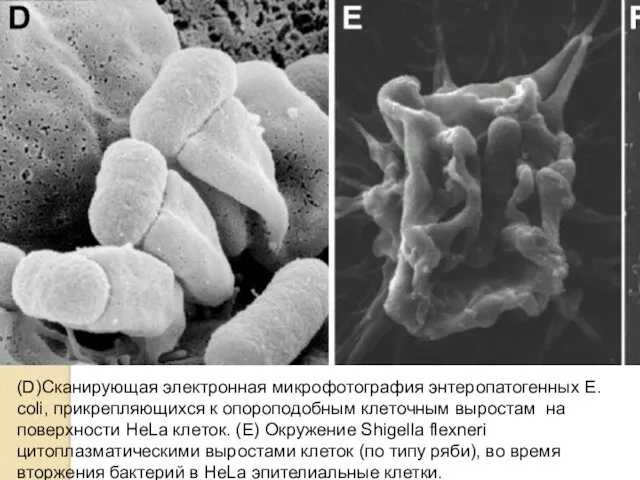
(D)Сканирующая электронная микрофотография энтеропатогенных Е. coli, прикрепляющихся к опороподобным клеточным выростам
(D)Сканирующая электронная микрофотография энтеропатогенных Е. coli, прикрепляющихся к опороподобным клеточным выростам

С образования биопленок начинается развитие любой инфекции.
Биопленки -тонкий слой микроорганизмов с
С образования биопленок начинается развитие любой инфекции.
Биопленки -тонкий слой микроорганизмов с

Биопленки
Это основной фенотип почти всех бактерий в естественных условиях обитания, как
Биопленки
Это основной фенотип почти всех бактерий в естественных условиях обитания, как

Биопленки
Существование бактерий в виде биопленок усиливает свою защиту от фагоцитоза, ультрафиолетового
Биопленки
Существование бактерий в виде биопленок усиливает свою защиту от фагоцитоза, ультрафиолетового

Адгезия микроорганизмов к имплантируемым устройствам.
Ни один из используемых для создания имплантируемых
Адгезия микроорганизмов к имплантируемым устройствам.
Ни один из используемых для создания имплантируемых
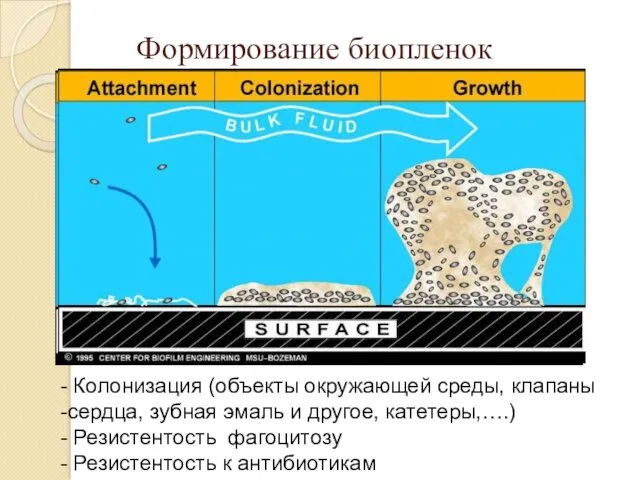
Формирование биопленок
Колонизация (объекты окружающей среды, клапаны
сердца, зубная эмаль и
Формирование биопленок
Колонизация (объекты окружающей среды, клапаны
сердца, зубная эмаль и

Факторы инвазии
Инвазия –проникновение возбудителя через слизистые и соединительнотканные барьеры
Агрессия – подавление
Факторы инвазии
Инвазия –проникновение возбудителя через слизистые и соединительнотканные барьеры
Агрессия – подавление

Факторы инвазии
Инвазия-распространения микроорганизмов в межклеточных пространствах тканей организма хозяина и проникновения
Факторы инвазии
Инвазия-распространения микроорганизмов в межклеточных пространствах тканей организма хозяина и проникновения

Факторы инвазии
Гиалуронидаза – деполимеризует гиалуроновую кислоту, высокополимерное соединение, состоящий из остатков
Факторы инвазии
Гиалуронидаза – деполимеризует гиалуроновую кислоту, высокополимерное соединение, состоящий из остатков

Факторы инвазии
Нейраминидаза- гидролизует гликозидные связи в гликопротеидах, ганглиозидах, отщепляет от них
Факторы инвазии
Нейраминидаза- гидролизует гликозидные связи в гликопротеидах, ганглиозидах, отщепляет от них

Факторы инвазии и агрессии
Лецитиназа – гидролизует лецитин (фосфоглицерид фосфатидилхолин) основной компонент
Факторы инвазии и агрессии
Лецитиназа – гидролизует лецитин (фосфоглицерид фосфатидилхолин) основной компонент

Лецитиназная активность
Лецитиназная активность

Протеолитические ферменты.
Основной целью протеолитических ферментов, образуемых бактериями, являются сигнальные и
Протеолитические ферменты.
Основной целью протеолитических ферментов, образуемых бактериями, являются сигнальные и
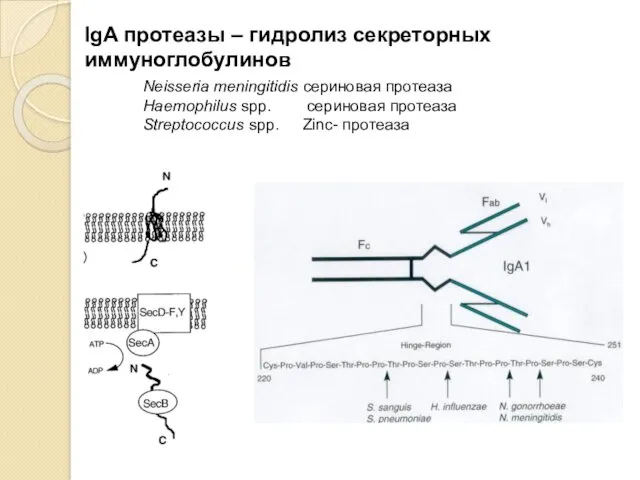
IgA протеазы – гидролиз секреторных иммуноглобулинов
Neisseria meningitidis сериновая протеаза
Haemophilus spp. сериновая протеаза
Streptococcus
IgA протеазы – гидролиз секреторных иммуноглобулинов
Neisseria meningitidis сериновая протеаза
Haemophilus spp. сериновая протеаза
Streptococcus

Ферменты.
ДНК-аза – гидролиз молекул ДНК, разрыв фосфодиэфирных связей распад ДНК
Ферменты.
ДНК-аза – гидролиз молекул ДНК, разрыв фосфодиэфирных связей распад ДНК
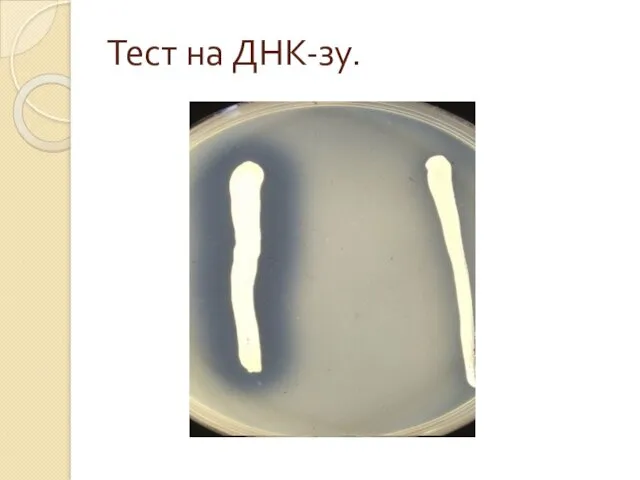
Тест на ДНК-зу.
Тест на ДНК-зу.

Тест на плазмокоагулазу
Тест на плазмокоагулазу

Ферменты
Уреаза - распад мочевины, аммиак вызывает защелачивание среды, прямой токсический эффект.
Ферменты
Уреаза - распад мочевины, аммиак вызывает защелачивание среды, прямой токсический эффект.

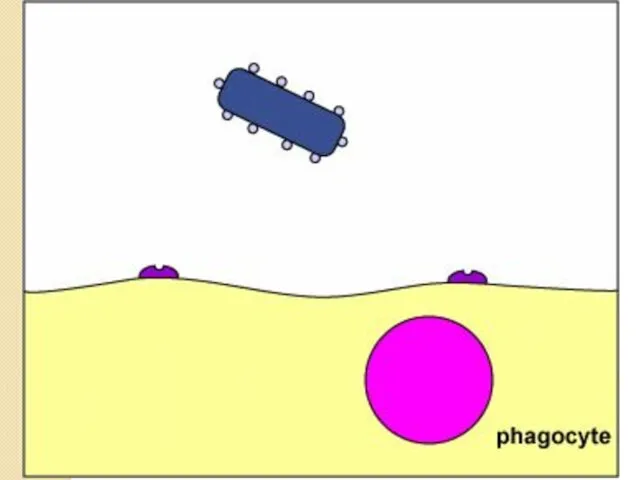
Антифагоцитарные факторы
Антифагоцитарные факторы

Стадии фагоцитоза
Стадии фагоцитоза

Антифагоцитарные факторы
Имеют поверхностную локализацию – капсулы, капсулоподобные структуры
Не являются жизненно важными
Антифагоцитарные факторы
Имеют поверхностную локализацию – капсулы, капсулоподобные структуры
Не являются жизненно важными

Антифагоцитарные факторы
Защита от фагоцитоза может происходить на различных стадиях процесса:
На стадии
Антифагоцитарные факторы
Защита от фагоцитоза может происходить на различных стадиях процесса:
На стадии

Число(фигура) 11. Негативное контрастирование Streptococcus pyogenes при электронной микроскопии (28,000X). Ореол
Число(фигура) 11. Негативное контрастирование Streptococcus pyogenes при электронной микроскопии (28,000X). Ореол
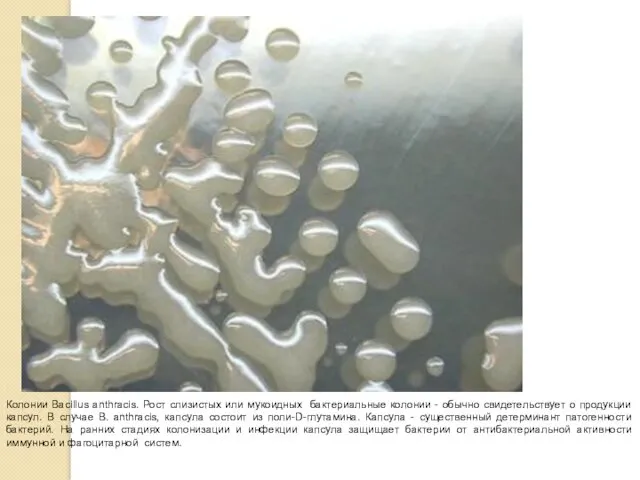
Колонии Bacillus anthracis. Рост слизистых или мукоидных бактериальные колонии - обычно
Колонии Bacillus anthracis. Рост слизистых или мукоидных бактериальные колонии - обычно
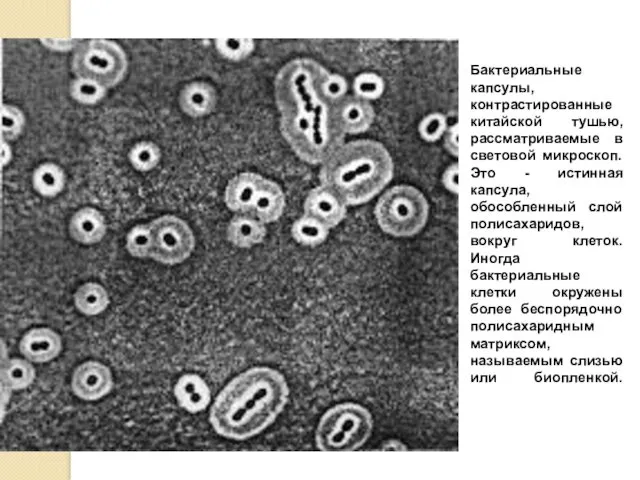
Бактериальные капсулы, контрастированные китайской тушью, рассматриваемые в световой микроскоп. Это -
Бактериальные капсулы, контрастированные китайской тушью, рассматриваемые в световой микроскоп. Это -

Антифагоцитарные факторы
Капсула –метод Бурри-Гинса
Антифагоцитарные факторы
Капсула –метод Бурри-Гинса
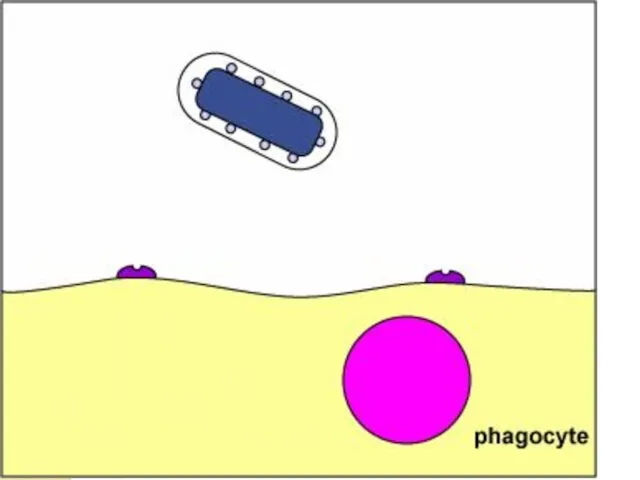


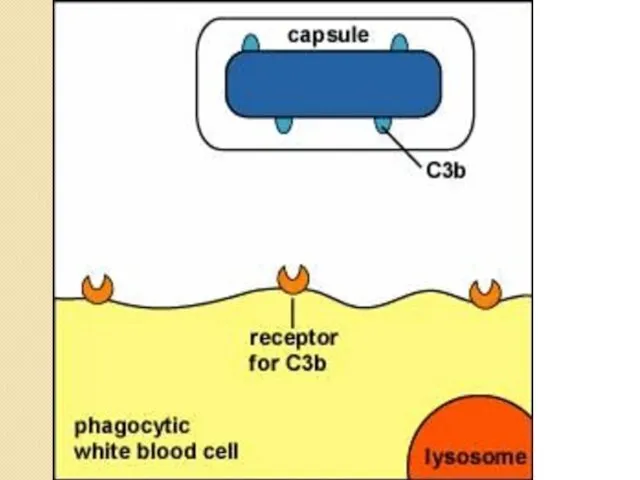
Антифагоцитарные факторы
Выживание микробных клеток после поглощения фагоцитом.
Препятствие слиянию фагосомы с лизосомой
Антифагоцитарные факторы
Выживание микробных клеток после поглощения фагоцитом.
Препятствие слиянию фагосомы с лизосомой
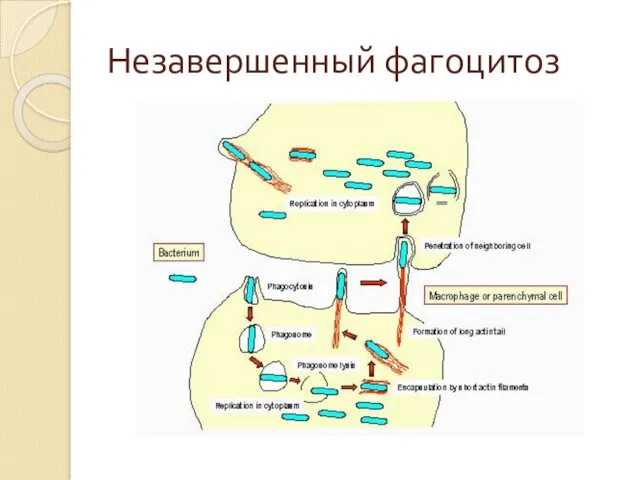
Незавершенный фагоцитоз
Незавершенный фагоцитоз

Инвазия нефагоцитирующих клеток
Активная инвазия клеток, не относящихся к фагоцитам, прежде всего
Инвазия нефагоцитирующих клеток
Активная инвазия клеток, не относящихся к фагоцитам, прежде всего

Уклонение от иммунного ответа
Вариабельность антигенных свойств
Антигенная мимикрия
Образование L-форм
Экранирование антигенных детерминант с
Уклонение от иммунного ответа
Вариабельность антигенных свойств
Антигенная мимикрия
Образование L-форм
Экранирование антигенных детерминант с
Streptococcus sp
Streptococcus sp
Pseudomonas
Pseudomonas
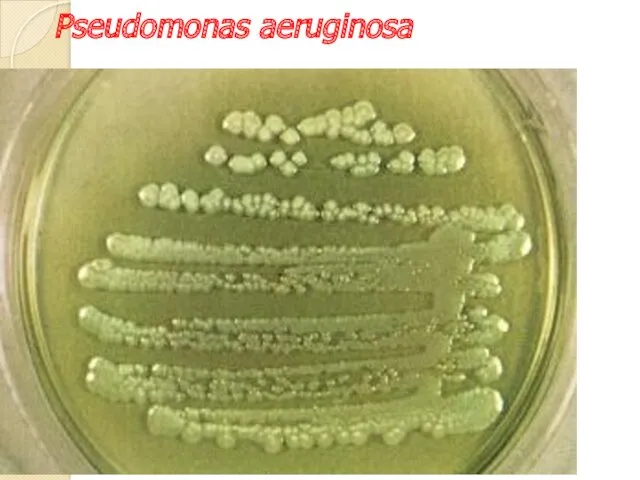
Pseudomonas aeruginosa
Pseudomonas aeruginosa

Бактериальные токсины
Оказывают непосредственное патологическое действие
Экзотоксины (белковые токсины)– выделяются преимущественно в окружающую
Бактериальные токсины
Оказывают непосредственное патологическое действие
Экзотоксины (белковые токсины)– выделяются преимущественно в окружающую

Бактериальные токсины
Характерные свойства белковых токсинов
Токсичность
Специфичность
Термолабильность
Иммуногенны-образуют анатоксины
Бактериальные токсины
Характерные свойства белковых токсинов
Токсичность
Специфичность
Термолабильность
Иммуногенны-образуют анатоксины

Бактериальные токсины
Простые – полипептидная цепь
Сложные – несколько связанных полипептидных цепей, соединенных
Бактериальные токсины
Простые – полипептидная цепь
Сложные – несколько связанных полипептидных цепей, соединенных

Классификация по механизму действия
Ингибируют синтез белка- цитотоксины
Повреждают клеточные мембраны-мембранотоксины
Нарушают передачу
Классификация по механизму действия
Ингибируют синтез белка- цитотоксины
Повреждают клеточные мембраны-мембранотоксины
Нарушают передачу
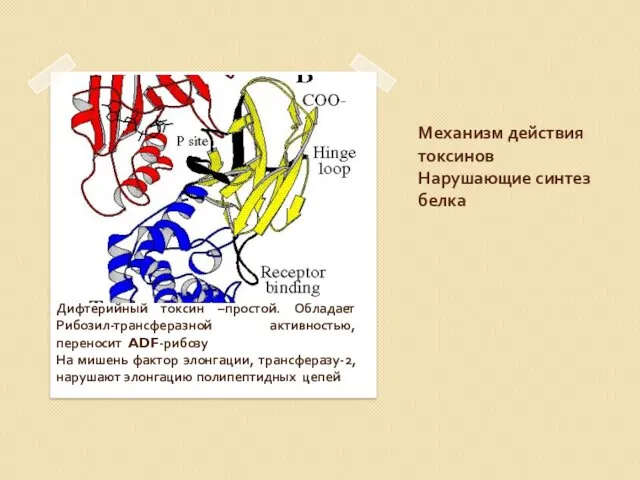
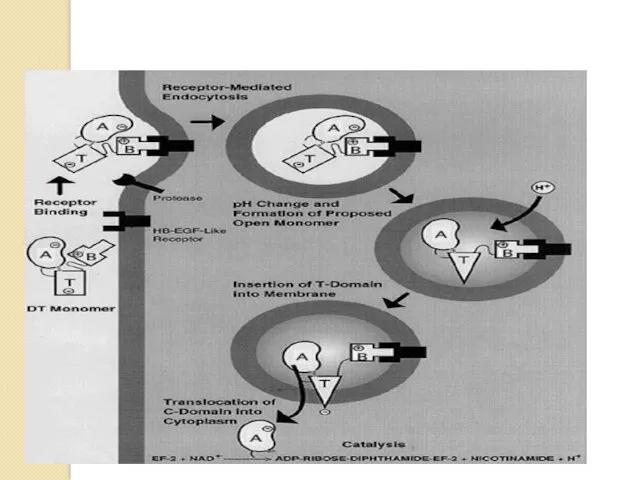
Механизм действия токсинов
Нарушающие синтез белка
Дифтерийный токсин –простой. Обладает Рибозил-трансферазной активностью, переносит
Механизм действия токсинов
Нарушающие синтез белка
Дифтерийный токсин –простой. Обладает Рибозил-трансферазной активностью, переносит

Токсины, нарушающие синтез белка
Шига-токсин – Субъединица А, обладающая ферментативной активностью, действует
Токсины, нарушающие синтез белка
Шига-токсин – Субъединица А, обладающая ферментативной активностью, действует

Порообразующие токсины.
Бактериальные токсины, функционирующие посредством вставки в плазматическую мембрану хозяина и
Порообразующие токсины.
Бактериальные токсины, функционирующие посредством вставки в плазматическую мембрану хозяина и

Токсины, повреждающие клеточные мембраны.
Порообразующие-гемолизины и лейкоцидин.
Могут повреждать моноциты, тромбоциты. Альфа токсин
Токсины, повреждающие клеточные мембраны.
Порообразующие-гемолизины и лейкоцидин.
Могут повреждать моноциты, тромбоциты. Альфа токсин

Типы гемолиза на кровяном агаре
Типы гемолиза на кровяном агаре

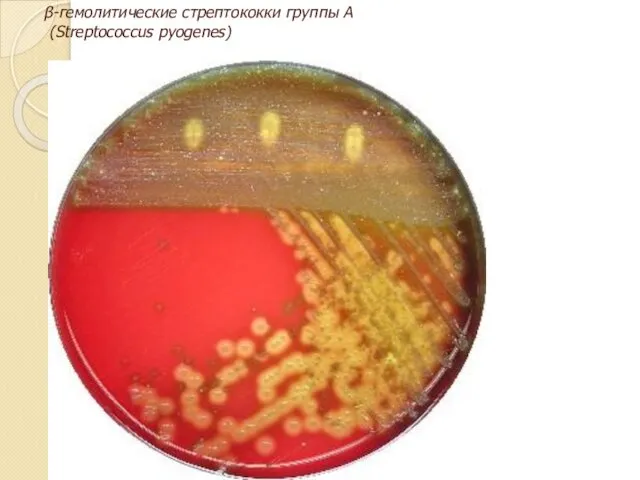
β-гемолитические стрептококки группы А
(Streptococcus pyogenes)
β-гемолитические стрептококки группы А
(Streptococcus pyogenes)

Функциональные блокаторы (активаторы путей метаболизма вторичных мессенджеров
Нарушающие функцию аданилатциклазы –
Холерный токсин
Функциональные блокаторы (активаторы путей метаболизма вторичных мессенджеров
Нарушающие функцию аданилатциклазы –
Холерный токсин

Холерный токсин
Холерный токсин

Нейротоксины C.botulinum (BoNT серотипов A vG) и C.tetani -протеазы
Нейротоксины синтезируются в
Нейротоксины C.botulinum (BoNT серотипов A vG) и C.tetani -протеазы
Нейротоксины синтезируются в
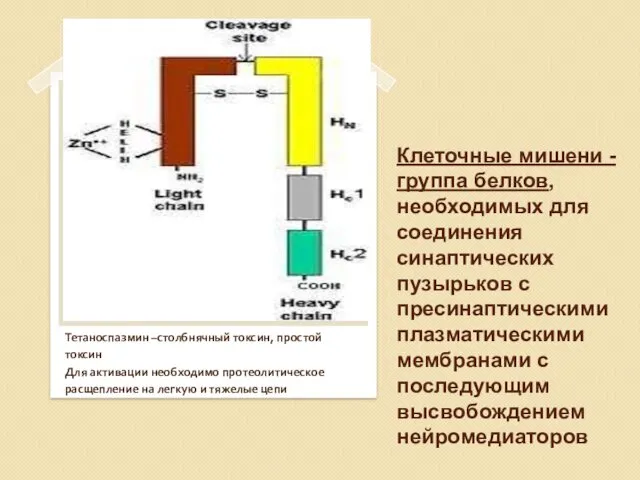
Клеточные мишени -группа белков, необходимых для соединения синаптических пузырьков с пресинаптическими
Клеточные мишени -группа белков, необходимых для соединения синаптических пузырьков с пресинаптическими

Нейротоксин
Столбнячный токсин поражает два вида нейронов. Он связывается с рецепторами пресинаптической
Нейротоксин
Столбнячный токсин поражает два вида нейронов. Он связывается с рецепторами пресинаптической

Протеолитические токсины нейротоксины
Обладает протеазной активностью, разрушает белок синаптобревин, блокирует систему торможения
Протеолитические токсины нейротоксины
Обладает протеазной активностью, разрушает белок синаптобревин, блокирует систему торможения

Токсины-суперантигены, активаторы иммунного ответа
Иммуностимулирующий потенциал токсинов является следствием их способности связывать
Токсины-суперантигены, активаторы иммунного ответа
Иммуностимулирующий потенциал токсинов является следствием их способности связывать
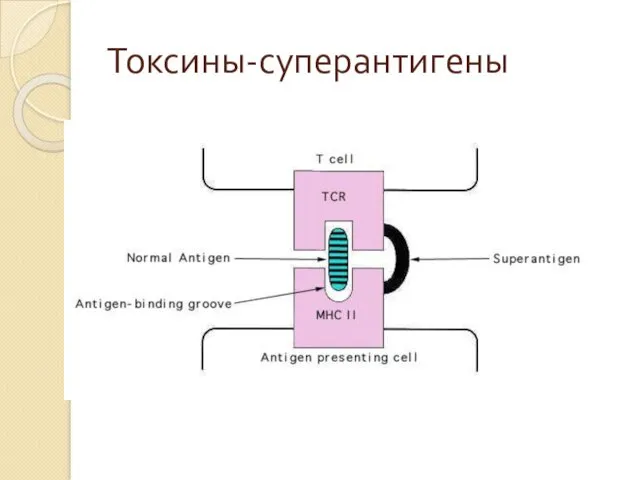
Токсины-суперантигены
Токсины-суперантигены

Эндотоксин
Сложный липополисахаридный комплекс, содержится в клеточной стенке грамотрицательных бактерий и выделяется
Эндотоксин
Сложный липополисахаридный комплекс, содержится в клеточной стенке грамотрицательных бактерий и выделяется
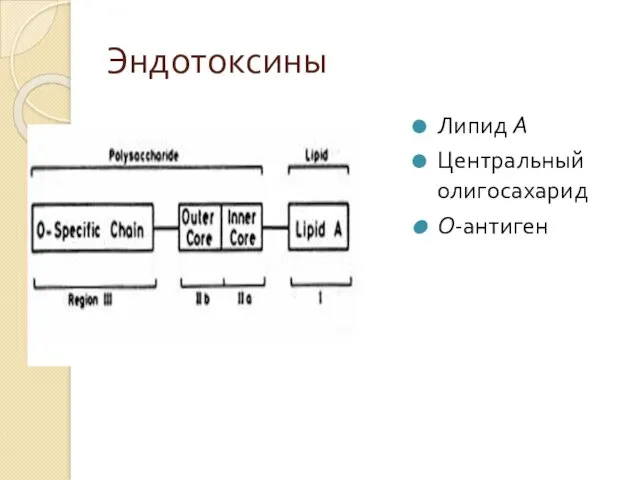
Эндотоксины
Липид А
Центральный олигосахарид
О-антиген
Эндотоксины
Липид А
Центральный олигосахарид
О-антиген
 Обмен липидов-2. Строение хиломикрона
Обмен липидов-2. Строение хиломикрона Свечение моря
Свечение моря Cladocera
Cladocera Отдел Моховидные
Отдел Моховидные ПРЕЗЕНТАЦИЯ ДЛЯ ИНТЕРАКТИВНОЙ ДОСКИ. Капуста. Семейство Крестоцветные (В двух частях)
ПРЕЗЕНТАЦИЯ ДЛЯ ИНТЕРАКТИВНОЙ ДОСКИ. Капуста. Семейство Крестоцветные (В двух частях) ЧМН - черепно–мозговые нервы
ЧМН - черепно–мозговые нервы Паразитизм, как явление. (Лекция 2)
Паразитизм, как явление. (Лекция 2) Водоросли (5 класс)
Водоросли (5 класс) Тура қанаттылар отряды. Шегіртке тұқымдасы
Тура қанаттылар отряды. Шегіртке тұқымдасы Neuropsychomodulating action of microbiota
Neuropsychomodulating action of microbiota Наекомые рекордсмены
Наекомые рекордсмены Загальна характеристика стафіло- стрепто- диплококів, їх роль в патології тварин. Лабораторна діагностика кокових інфекцій
Загальна характеристика стафіло- стрепто- диплококів, їх роль в патології тварин. Лабораторна діагностика кокових інфекцій Пищеварительная система. Ротовая полость
Пищеварительная система. Ротовая полость Клонирование генов
Клонирование генов Нервная регуляция
Нервная регуляция General characteristics of 6 Kingdoms
General characteristics of 6 Kingdoms Строение глаза
Строение глаза Ткани. Эпителиальная ткань. Соединительная ткань. Мышечная ткань
Ткани. Эпителиальная ткань. Соединительная ткань. Мышечная ткань Грибы как царство живой природы. Грибы нашего леса
Грибы как царство живой природы. Грибы нашего леса Семейство лютиковые
Семейство лютиковые Лабораторная диагностика паразитарных заболеваний. Занятие 11
Лабораторная диагностика паразитарных заболеваний. Занятие 11 Слуховая система человека
Слуховая система человека Многообразие грибов как одного из компонентов экосистем
Многообразие грибов как одного из компонентов экосистем Животный мир Беларуси
Животный мир Беларуси Гипотеза А.И. Опарина о происхождении жизни на Земле
Гипотеза А.И. Опарина о происхождении жизни на Земле Необычные грибы
Необычные грибы Каких животных называем копытные?
Каких животных называем копытные? Витамины. Определение, классификация
Витамины. Определение, классификация