- Физиология дыхания. Лекция 24

Содержание
- 2. Дыхательный центр. Под дыхательным центром в узком (анатомическом) смысле понимают совокупность нейронов специфических (дыхательных) ядер продолговатого
- 3. Дыхательный центр выполняет две основные функции в системе дыхания: моторную, или двигательную, которая проявляется в виде
- 4. Локализация и функциональные свойства дыхательных нейронов. Нейроны дыхательного центра локализованы в дорсомедиальной и вентролатеральной областях продолговатого
- 5. На рис. 44 схематично изображены паттерны электрической активности дыхательных нейронов. Нейроны дыхательного центра в зависимости от
- 6. Другие области локализации дыхательных нейронов. В мосту находятся два ядра дыхательных нейронов, которые называют пневмотаксическим центром.
- 7. Диафрагмальные мотонейроны образуют диафрагмальный нерв. Нейроны расположены узким столбом в медиальной части вентральных рогов от СIII
- 8. Генерация дыхательного ритма. Спонтанная активность нейронов дыхательного центра начинает появляться к концу периода внутриутробного развития. В
- 9. Стартовый разряд ранних инспираторных нейронов начинает активировать полные инспираторные нейроны, которые способны возбуждать друг друга. Полные
- 10. Вторая половина фазы выдоха, или фаза активной экспирации, полностью зависит от механизма ритмогенеза инспираторной и постинспираторной
- 11. Рефлекторная регуляция дыхания. Рефлекторная регуляция дыхания осуществляется благодаря тому, что нейроны дыхательного центра имеют связи с
- 12. Рефлексы с гортани и трахеи. Многочисленные нервные окон¬чания расположены между эпителиальными клетками слизистой оболочки гортани и
- 13. Рефлексы с J-рецепторов. В альвеолярных перегородках в кон¬такте с капиллярами находятся особые J-рецепторы. Эти рецепторы особенно
- 14. Проприоцептивный контроль дыхания. Рецепторы суставов грудной клетки посылают импульсы в кору больших полушарий и являются единственным
- 15. Артериальные (периферические) хеморецепторы. Периферические хеморецепторы находятся в каротидных и аортальных тельцах. Сигналы от артериальных хеморецепторов по
- 16. Центральные хеморецепторы. Окончательно не установлено местоположение центральных хеморецепторов. Исследователи считают, что такие хеморецепторы находятся в ростральных
- 17. Реакция дыхания на СО2. Гиперкапния и ацидоз стимулируют, а гипокапния и алкалоз тормозят центральные хеморецепторы. При
- 18. Координация дыхания с другими функциями организма. В отличие от других физиологических функций организма дыхание находится под
- 19. Корковая регуляция дыхания. Деятельность всей совокупности нейронов, образующих ДЦ, необходима для сохранения нормального дыхания. Однако в
- 20. 24. 2 Особенности дыхания и его регуляция при мышечной работе, при пониженном и повышенном атмосферном давлении.
- 21. За исключением циркуляторной гипоксии, происходящей в случае недостаточного притока крови к отдельным органам, остальные формы гипоксии
- 22. Особенности дыхания при пониженном атмосферном давлении. Наиболее изученной формой гипоксии является гипоксемическая гипоксия, особенно ее дыхательная
- 23. Первоначальная гипоксическая стимуляция дыхания, возникающая при подъеме на высоту, приводит к вымыванию из крови СО2 и
- 24. Особенности дыхания при повышенном атмосферном давлении. В то время, как низкое атмосферное давление ведет к химическим
- 25. Дыхание при мышечной работе. Интенсивность дыхания тесно связана с интенсивностью окислительных процессов: глубина и частота дыхательных
- 26. Одной из причин увеличения легочной вентиляции при интенсивной мышечной работе является накопление молочной кислоты в тканях
- 27. Дыхание чистым кислородом. Гипербарическая оксигенация. В клинической практике иногда возникает потребность в повышении РО2 в артериальной
- 28. 24.3. Характеристика функциональной системы, поддерживающей постоянство газового состава крови и ее схема. Для поддержания нормального уровня
- 30. Скачать презентацию

Дыхательный центр.
Под дыхательным центром в узком (анатомическом) смысле понимают совокупность
Дыхательный центр.
Под дыхательным центром в узком (анатомическом) смысле понимают совокупность

Дыхательный центр выполняет две основные функции в системе дыхания: моторную, или
Дыхательный центр выполняет две основные функции в системе дыхания: моторную, или

Локализация и функциональные свойства дыхательных нейронов.
Нейроны дыхательного центра локализованы в
Локализация и функциональные свойства дыхательных нейронов.
Нейроны дыхательного центра локализованы в
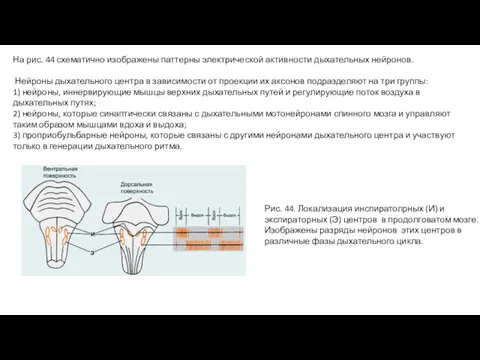
На рис. 44 схематично изображены паттерны электрической активности дыхательных нейронов.
Нейроны
На рис. 44 схематично изображены паттерны электрической активности дыхательных нейронов.
Нейроны

Другие области локализации дыхательных нейронов.
В мосту находятся два ядра дыхательных
Другие области локализации дыхательных нейронов.
В мосту находятся два ядра дыхательных

Диафрагмальные мотонейроны образуют диафрагмальный нерв. Нейроны расположены узким столбом в медиальной
Диафрагмальные мотонейроны образуют диафрагмальный нерв. Нейроны расположены узким столбом в медиальной

Генерация дыхательного ритма.
Спонтанная активность нейронов дыхательного центра начинает появляться к
Генерация дыхательного ритма.
Спонтанная активность нейронов дыхательного центра начинает появляться к

Стартовый разряд ранних инспираторных нейронов начинает активировать полные инспираторные нейроны, которые
Стартовый разряд ранних инспираторных нейронов начинает активировать полные инспираторные нейроны, которые

Вторая половина фазы выдоха, или фаза активной экспирации, полностью зависит от
Вторая половина фазы выдоха, или фаза активной экспирации, полностью зависит от

Рефлекторная регуляция дыхания. Рефлекторная регуляция дыхания осуществляется благодаря тому, что нейроны
Рефлекторная регуляция дыхания. Рефлекторная регуляция дыхания осуществляется благодаря тому, что нейроны

Рефлексы с гортани и трахеи. Многочисленные нервные окон¬чания расположены между эпителиальными
Рефлексы с гортани и трахеи. Многочисленные нервные окон¬чания расположены между эпителиальными

Рефлексы с J-рецепторов. В альвеолярных перегородках в кон¬такте с капиллярами
Рефлексы с J-рецепторов. В альвеолярных перегородках в кон¬такте с капиллярами

Проприоцептивный контроль дыхания. Рецепторы суставов грудной клетки посылают импульсы в кору
Проприоцептивный контроль дыхания. Рецепторы суставов грудной клетки посылают импульсы в кору

Артериальные (периферические) хеморецепторы.
Периферические хеморецепторы находятся в каротидных и аортальных тельцах.
Артериальные (периферические) хеморецепторы.
Периферические хеморецепторы находятся в каротидных и аортальных тельцах.

Центральные хеморецепторы.
Окончательно не установлено местоположение центральных хеморецепторов. Исследователи считают, что
Центральные хеморецепторы.
Окончательно не установлено местоположение центральных хеморецепторов. Исследователи считают, что

Реакция дыхания на СО2.
Гиперкапния и ацидоз стимулируют, а гипокапния и
Реакция дыхания на СО2.
Гиперкапния и ацидоз стимулируют, а гипокапния и

Координация дыхания с другими функциями организма.
В отличие от других физиологических
Координация дыхания с другими функциями организма.
В отличие от других физиологических

Корковая регуляция дыхания.
Деятельность всей совокупности нейронов, образующих ДЦ, необходима для
Корковая регуляция дыхания.
Деятельность всей совокупности нейронов, образующих ДЦ, необходима для

24. 2 Особенности дыхания и его регуляция при мышечной работе,
24. 2 Особенности дыхания и его регуляция при мышечной работе,

За исключением циркуляторной гипоксии, происходящей в случае недостаточного притока крови
За исключением циркуляторной гипоксии, происходящей в случае недостаточного притока крови

Особенности дыхания при пониженном атмосферном давлении.
Наиболее изученной формой гипоксии является гипоксемическая
Особенности дыхания при пониженном атмосферном давлении.
Наиболее изученной формой гипоксии является гипоксемическая

Первоначальная гипоксическая стимуляция дыхания, возникающая при подъеме на высоту, приводит к
Первоначальная гипоксическая стимуляция дыхания, возникающая при подъеме на высоту, приводит к

Особенности дыхания при повышенном атмосферном давлении.
В то время, как низкое
Особенности дыхания при повышенном атмосферном давлении.
В то время, как низкое

Дыхание при мышечной работе.
Интенсивность дыхания тесно связана с интенсивностью окислительных
Дыхание при мышечной работе.
Интенсивность дыхания тесно связана с интенсивностью окислительных

Одной из причин увеличения легочной вентиляции при интенсивной мышечной работе является
Одной из причин увеличения легочной вентиляции при интенсивной мышечной работе является

Дыхание чистым кислородом.
Гипербарическая оксигенация. В клинической практике иногда возникает потребность
Дыхание чистым кислородом.
Гипербарическая оксигенация. В клинической практике иногда возникает потребность
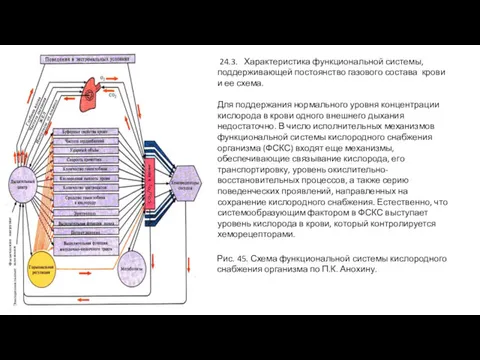
24.3. Характеристика функциональной системы, поддерживающей постоянство газового состава крови и
24.3. Характеристика функциональной системы, поддерживающей постоянство газового состава крови и
 Эволюция черепа, жаберного и челюстного аппарата позвоночных
Эволюция черепа, жаберного и челюстного аппарата позвоночных Особенности пищеварения у собак и котов. Питательные и биологически активные вещества
Особенности пищеварения у собак и котов. Питательные и биологически активные вещества Растениеводство. Системы земледелия
Растениеводство. Системы земледелия Биологические и хозяйственные особенности кур
Биологические и хозяйственные особенности кур Формирование целостной картины мира. Играем на лугу. Луг и его обитатели
Формирование целостной картины мира. Играем на лугу. Луг и его обитатели Копитні ссавці
Копитні ссавці Задания ЕГЭ по теме Клетка
Задания ЕГЭ по теме Клетка Применение активных дрожжей в кормлений животных
Применение активных дрожжей в кормлений животных Влияние света на проращивание семян (часть 1)
Влияние света на проращивание семян (часть 1) Ошущение. Возникновение ощущений
Ошущение. Возникновение ощущений Тип Членистоногие, класс Паукообразные
Тип Членистоногие, класс Паукообразные Птицы Африки
Птицы Африки Стрес та його чинники. Адаптація людини до стресу
Стрес та його чинники. Адаптація людини до стресу Биология – наука о живой природе
Биология – наука о живой природе Презентация по теме Проектная деятельность
Презентация по теме Проектная деятельность Взаимодействие микроорганизмов с человеком и животными
Взаимодействие микроорганизмов с человеком и животными Молекулярні механізми скорочення м’язового волокна
Молекулярні механізми скорочення м’язового волокна Ядовитые растения
Ядовитые растения Факторы среды, влияющие на размножение и формирование пола у животных. (Лекция 3)
Факторы среды, влияющие на размножение и формирование пола у животных. (Лекция 3) Красная книга Таштагольского района
Красная книга Таштагольского района Фенологические наблюдения за природой
Фенологические наблюдения за природой Анатомия ЦНС. Задний мозг. Варолиев мост
Анатомия ЦНС. Задний мозг. Варолиев мост Экология человека
Экология человека Строение и жизнедеятельность бактерий
Строение и жизнедеятельность бактерий Молекулярно-биологические методы диагностики
Молекулярно-биологические методы диагностики Всё о мёде, способы фальсификации
Всё о мёде, способы фальсификации Электрофорез белков в полиакриламидном геле и вестерн-блоттинг
Электрофорез белков в полиакриламидном геле и вестерн-блоттинг Мышцы шеи
Мышцы шеи