- Физиология микроорганизмов

Содержание
- 2. Определение понятий Питание бактерий Пластический обмен Энергетический обмен. Дыхание бактерий. Рост и размножение бактерий Культивирование бактерий
- 3. Физиологический период Луи́ Пасте́р Роберт Кох (1822 – 1822) (1848 – 1910)
- 4. Выдающийся русский микробиолог, основатель экологической микробиологии и почвенной микробиологии. Открыл хемосинтезирующие микроорганизмы, выполняющие важную роль в
- 5. Ма́ртин Ви́ллем Бе́йеринк (Martinus Willem Beijerinck; 1851—1931) Первооткрыватель симбиотических азотфиксаторов (1888 год), свободноживущих аэробных азотфиксаторов рода
- 9. ПРОТОТРОФЫ Микроорганизмы, способные синтезировать все необходимые им органические соединения (первичные и вторичные метаболиты) из одного источника
- 10. ФАКТОРЫ РОСТА К факторам роста относят аминокислоты, пуриновые и пиримидиновые основания, липиды, витамины, железопорфирины (гем) и
- 12. транспорт веществ через мембрану
- 14. Активный транспорт неизмененных молекул Большинство метаболитов, ионов и других веществ транспортируются в клетку активным путем с
- 15. Транслокация групп
- 16. Выход веществ из клетки осуществляется за счет диффузии и при участии транспортных систем. Бактерии синтезируют и
- 17. Секреция белков бактериями осуществляется с помощью различных систем и механизмов. При этом следует различать секрецию белков
- 18. механизмы секреции белка-предшественника лидерными пептидами через ЦМ Модель прямого транспорта предполагает прямое вхождение лидерного пептида в
- 19. Бактериоцины, кодируемые различными плазмидами, не содержат в своей структуре сигнальных пептидов. Для их секреции через ЦМ
- 20. Ферменты бактерий. (греч. fermentum - закваска) Открытию различных ферментов и изучению путей биохимических реакций, катализируемых ими,
- 21. 1) оксидоредуктазы - катализируют окислительно-восстановительные реакции - дегидрогеназы, оксидазы и др.; 2) трансферазы - катализируют реакции
- 22. Ферменты бактерий. Эндоферменты - катализируют метаболизм, проходящий внутри клетки. Это, например, окислительно-восстановительные ферменты цитоплазматической мембраны, участвующие
- 23. Ферменты бактерий. В соответствии с особенностями генетического контроля Конститутивные, синтез которых происходит в течение всего клеточного
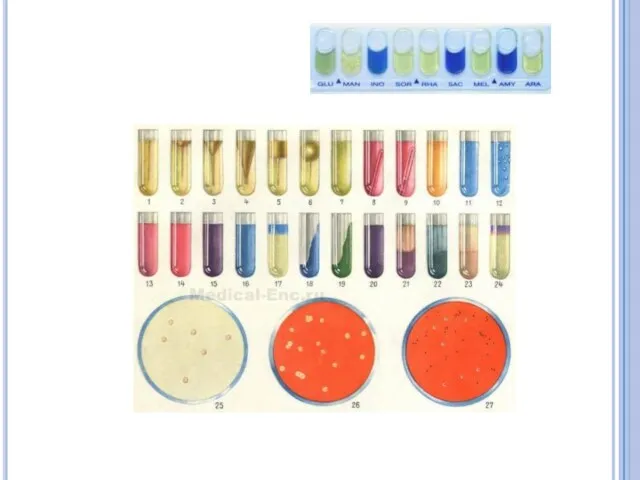
- 24. Ферменты бактерий. Сахаролитические или гликолитические (расщепление сахаров) Протеолитические (разложение белков) Липолитические (расщепление жиров) и другие, Ферментный
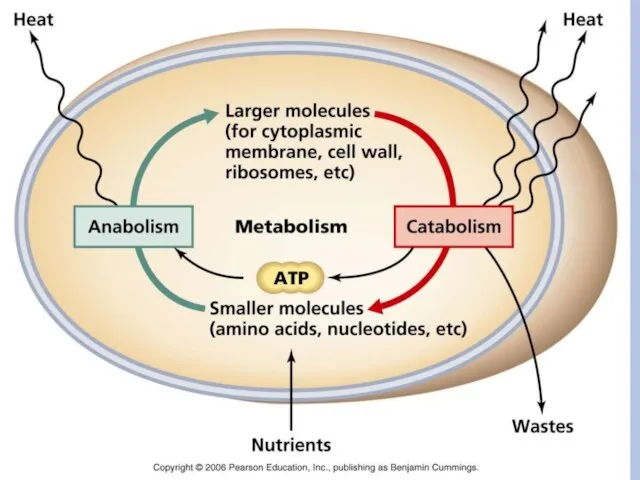
- 29. Пластический (конструктивный) метаболизм В конструктивном обмене можно выделить две группы биосинтетических процессов: биосинтез мономеров (аминокислот, нуклеотидов,
- 31. Биосинтез углеводов. Микроорганизмы синтезируют моно-, олиго-, полисахариды и другие соединения, в состав которых входят углеводы. Автотрофы
- 32. Биосинтез аминокислот. Только несколько аминокислот (пируват → аланин, α-кетоглутарат → аспартат и фумарат → глутамат) образуются
- 33. Биосинтез аминокислот. Многие прокариоты, так же как эукариоты, могут получать аминокислоты из молекул белка, которые предварительно
- 34. Биосинтез белков. Основу конструктивного обмена составляет биосинтез белков, который находится под контролем генетической системы организма и
- 35. Биосинтез липидов. Липиды микроорганизмов представлены жирными кислотами, фосфолипидами, воском, терпенами, каротиноидами, которые содержат длинноцепочечные насыщенные и
- 36. Биосинтез липидов. Последовательное удлинение этих фрагментов приводит в конечном счете к образованию высших жирных кислот, содержащих
- 37. Энергетический метаболизм Дыхание, или биологическое окисление, основано на окислительно-восстановительных реакциях, идущих с образованием АТФ - универсального
- 38. Акцептором водорода или электронов может быть молекулярный кислород (такое дыхание называется аэробным) или нитрат, сульфат, фумарат
- 39. Облигатные аэробы могут расти только при наличии кислорода. При наличии кислорода бактерии образуют перекисные радикалы кислорода,
- 40. Получение энергии путем субстратного фосфорилирования. Брожение. У микроорганизмов выделяют три пути превращения сахаров в основной энергетический
- 41. Гликолиз (путь Эмдена - Мейергофа). В результате расщепления глюкозы расходуется 2 и синтезируется 4 молекулы АТФ.
- 43. Брожение Для облигатных и факультативных анаэробов характерно брожение - способом получения энергии в результате окислительно-восстановительных реакций,
- 45. Молочнокислое брожение. Бактерии родов Lactobacillus, Streptococcus, Bifidobacterium1 способны образовывать из пирувата молочную кислоту. При этом в
- 46. Маслянокислое брожение. Одним из основных продуктов брожения является масляная кислота. При этом типе брожения образуются также
- 47. Муравьинокислое брожение. Этот тип брожения характерен для представителей семейства энтеробактерий. Одним из конечных продуктов данного типа
- 48. Пропионовокислое брожение характерно для пропионобактерий, которые из пирувата образуют пропионовую кислоту. Многие бактерии при сбраживании углеводов
- 49. Пентозофосфатный, или гексозомонофосфатный (ГМФ), путь. Этот путь расщепления глюкозы характерен для многих микроорганизмов и не приводит
- 50. Кетодезоксифосфоглюконатный (КДФГ) путь (Энтнера - Дудорова). У высших организмов КДФГ-путь отсутствует. Пировиноградная кислота образуется у этих
- 51. Получение энергии путем окислительного фосфорилирования. При субстратном фосфорилировании из глюкозы или других источников углерода получают лишь
- 52. . Пируват, образующийся в процессе гликолиза, окисляется до ацетил-КоА , который при взаимодействии с уксусной кислотой
- 53. У всех аэробных и факультативно-анаэробных бактерий дыхательная цепь локализована на цитоплазматической мембране. Перенос электронов на молекулярный
- 55. У бактерий распространены системы окисления субстрата, связанные не с цитохромами, а с флавинзависимыми оксидазами, которые опосредуют
- 56. Поскольку при окислительном фосфорилировании высвобождается гораздо больше энергии, чем при брожении, некоторые бактерии осуществляют такой тип
- 57. Рост и размножение бактерий. Жизнедеятельность бактерий характеризуется ростом - формированием структурно-функциональных компонентов клетки и увеличением самой
- 58. Бактерии размножаются путем бинарного деления пополам, реже путем почкования. Актиномицеты, как и грибы, могут размножаться спорами.
- 59. Грамположительные бактерии делятся путем врастания синтезирующихся перегородок деления внутрь клетки, а грамотрицательные - путем перетяжки, в
- 60. Размножение бактерий в жидкой питательной среде. Бактерии, засеянные в определенный, не изменяющийся объем питательной среды, размножаясь,
- 62. При выращивании бактерий на жидкой питательной среде наблюдается придонный, диффузный или поверхностный (в виде пленки) рост
- 64. Размножение бактерий на плотной питательной среде. Бактерии, растущие на плотных питательных средах, образуют изолированные колонии различной
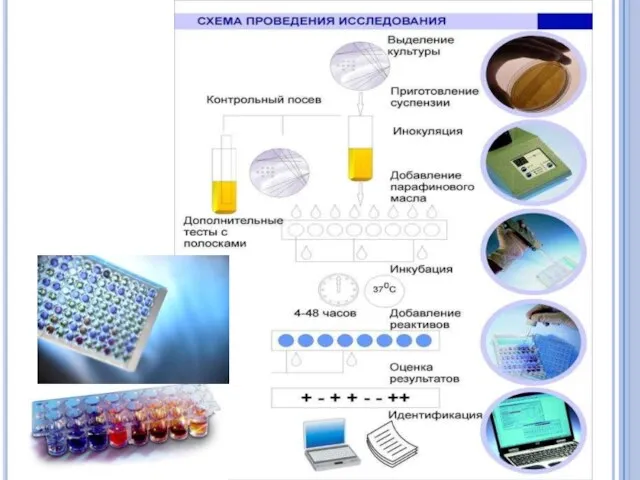
- 66. Питательные среды Питательные среды должны обязательно отвечать основным требованиям: они должны содержать в достаточном количестве все
- 67. Питательные среды Существует много различных вариантов питательных сред, сконструированных с учетом потребностей соответствующих видов бактерий и
- 68. По назначению питательные среды подразделяют на следующие основные категории. Универсальные Селективные (синонимы: избирательные, элективные, обогатительные) Дифференциально-селективные
- 69. Универсальные — среды, на которых хорошо растут многие виды патогенных и непатогенных бактерий. К ним относятся:
- 70. Дифференциально-диагностические — среды, позволяющие отличать одни виды бактерий от других по их ферментативной активности или культуральным
- 71. Селективные (синонимы: избирательные, элективные, обогатительные) - среды, содержащие вещества, используемые микроорганизмами определенных видов и не благоприятствующие
- 72. Дифференциально-селективные — среды, сочетающие в себе свойства дифференциально-диагностических и селективных сред. Они используются, в частности, для
- 73. Синтетические — среды строго определенного химического состава, представляющие собой растворы неорганических солей с добавлением химических соединений,
- 74. Специальные — среды, специально приготовленные для получения роста тех бактерий, которые не растут или очень плохо
- 75. Синтетические — среды строго определенного химического состава, представляющие собой растворы неорганических солей с добавлением химических соединений,
- 77. Скачать презентацию

Определение понятий
Питание бактерий
Пластический обмен
Энергетический обмен. Дыхание бактерий.
Рост и размножение бактерий
Культивирование бактерий
Определение понятий
Питание бактерий
Пластический обмен
Энергетический обмен. Дыхание бактерий.
Рост и размножение бактерий
Культивирование бактерий

Физиологический
период
Луи́ Пасте́р Роберт Кох
(1822 – 1822) (1848 – 1910)
Физиологический
период
Луи́ Пасте́р Роберт Кох
(1822 – 1822) (1848 – 1910)

Выдающийся русский микробиолог, основатель экологической микробиологии и почвенной микробиологии. Открыл
Выдающийся русский микробиолог, основатель экологической микробиологии и почвенной микробиологии. Открыл

Ма́ртин Ви́ллем Бе́йеринк (Martinus Willem Beijerinck; 1851—1931)
Первооткрыватель симбиотических азотфиксаторов
Ма́ртин Ви́ллем Бе́йеринк (Martinus Willem Beijerinck; 1851—1931)
Первооткрыватель симбиотических азотфиксаторов
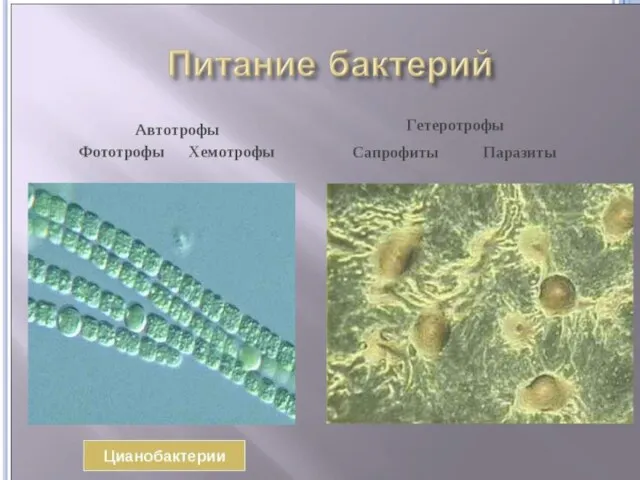
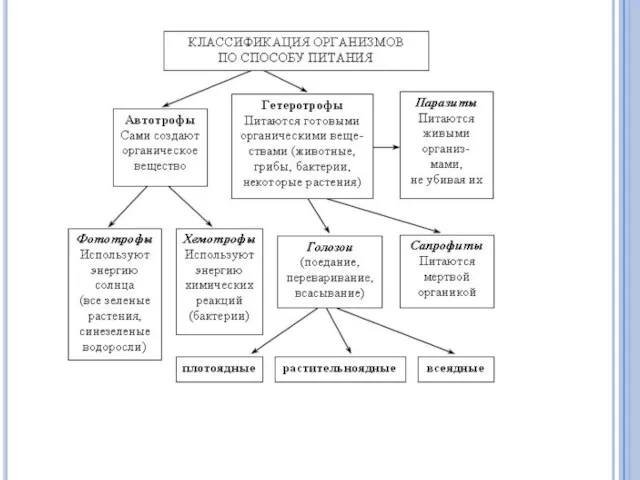

ПРОТОТРОФЫ
Микроорганизмы, способные синтезировать все необходимые им органические соединения (первичные и
ПРОТОТРОФЫ
Микроорганизмы, способные синтезировать все необходимые им органические соединения (первичные и

ФАКТОРЫ РОСТА
К факторам роста относят аминокислоты, пуриновые и пиримидиновые основания, липиды,
ФАКТОРЫ РОСТА
К факторам роста относят аминокислоты, пуриновые и пиримидиновые основания, липиды,
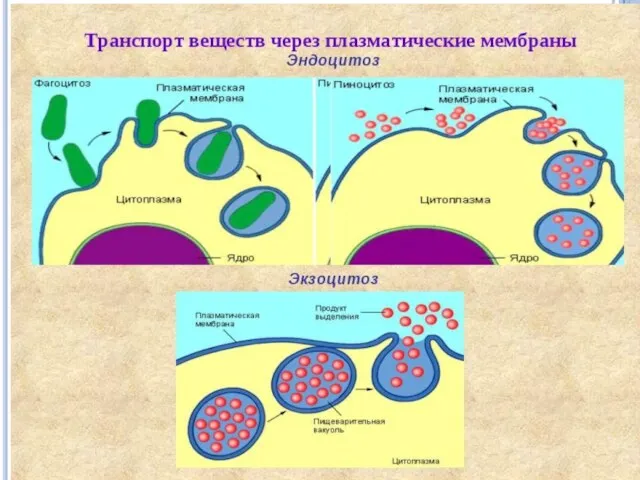
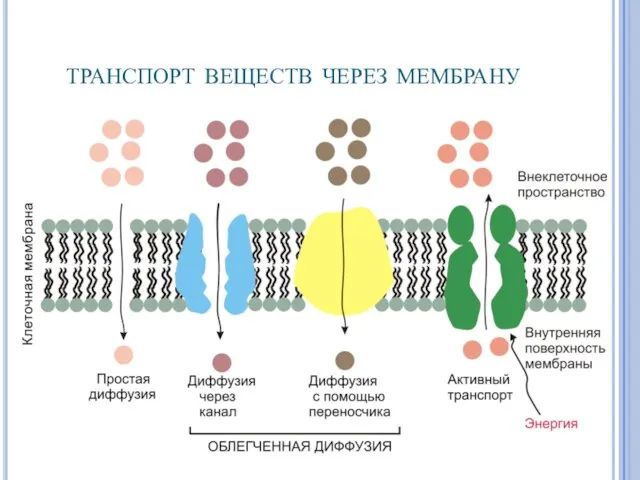
транспорт веществ через мембрану
транспорт веществ через мембрану
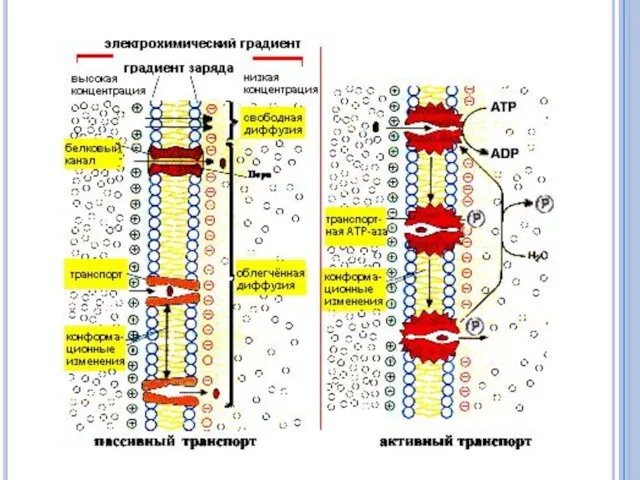

Активный транспорт неизмененных молекул
Большинство метаболитов, ионов и других веществ транспортируются в
Активный транспорт неизмененных молекул
Большинство метаболитов, ионов и других веществ транспортируются в
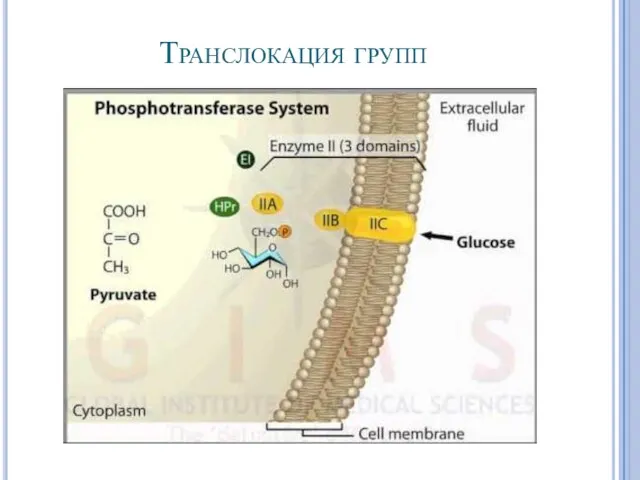
Транслокация групп
Транслокация групп

Выход веществ из клетки осуществляется за счет диффузии и при участии
Выход веществ из клетки осуществляется за счет диффузии и при участии

Секреция белков бактериями осуществляется с помощью различных систем и механизмов. При
Секреция белков бактериями осуществляется с помощью различных систем и механизмов. При

механизмы секреции белка-предшественника лидерными пептидами через ЦМ
Модель прямого транспорта предполагает
механизмы секреции белка-предшественника лидерными пептидами через ЦМ
Модель прямого транспорта предполагает

Бактериоцины, кодируемые различными плазмидами, не содержат в своей структуре сигнальных пептидов.
Бактериоцины, кодируемые различными плазмидами, не содержат в своей структуре сигнальных пептидов.
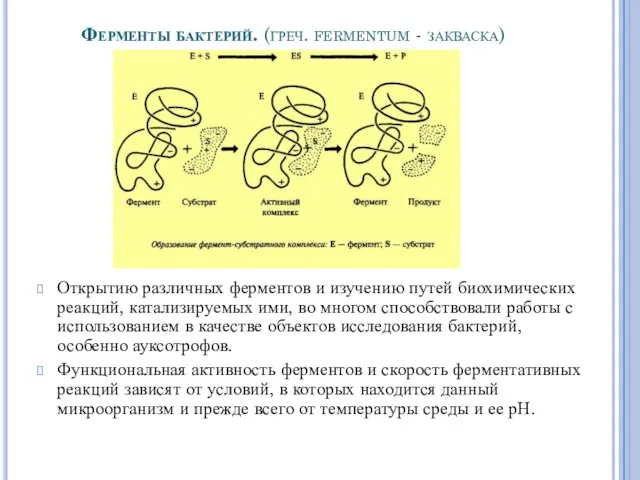
Ферменты бактерий. (греч. fermentum - закваска)
Открытию различных ферментов и изучению
Ферменты бактерий. (греч. fermentum - закваска)
Открытию различных ферментов и изучению

1) оксидоредуктазы - катализируют окислительно-восстановительные реакции - дегидрогеназы, оксидазы и др.;
2)
1) оксидоредуктазы - катализируют окислительно-восстановительные реакции - дегидрогеназы, оксидазы и др.;
2)

Ферменты бактерий.
Эндоферменты - катализируют метаболизм, проходящий внутри клетки. Это, например, окислительно-восстановительные
Ферменты бактерий.
Эндоферменты - катализируют метаболизм, проходящий внутри клетки. Это, например, окислительно-восстановительные

Ферменты бактерий.
В соответствии с особенностями
генетического контроля
Конститутивные, синтез которых происходит
Ферменты бактерий.
В соответствии с особенностями
генетического контроля
Конститутивные, синтез которых происходит

Ферменты бактерий.
Сахаролитические или гликолитические (расщепление сахаров)
Протеолитические (разложение белков)
Липолитические (расщепление жиров)
и
Ферменты бактерий.
Сахаролитические или гликолитические (расщепление сахаров)
Протеолитические (разложение белков)
Липолитические (расщепление жиров)
и



Пластический (конструктивный) метаболизм
В конструктивном обмене можно выделить две группы биосинтетических процессов:
Пластический (конструктивный) метаболизм
В конструктивном обмене можно выделить две группы биосинтетических процессов:

Биосинтез углеводов.
Микроорганизмы синтезируют моно-, олиго-, полисахариды и другие соединения, в состав
Биосинтез углеводов.
Микроорганизмы синтезируют моно-, олиго-, полисахариды и другие соединения, в состав
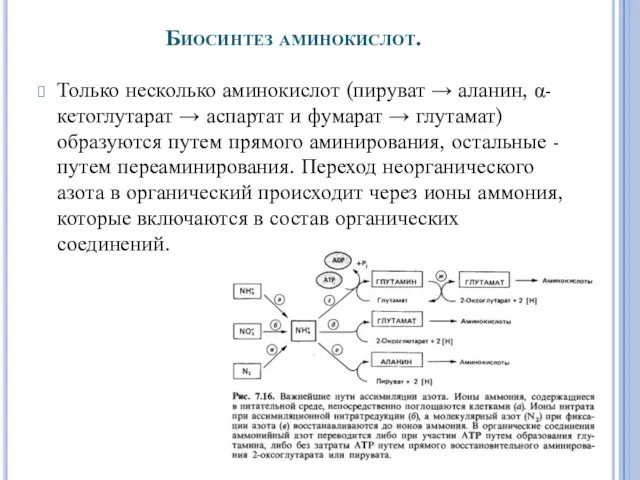
Биосинтез аминокислот.
Только несколько аминокислот (пируват → аланин, α-кетоглутарат → аспартат и
Биосинтез аминокислот.
Только несколько аминокислот (пируват → аланин, α-кетоглутарат → аспартат и

Биосинтез аминокислот.
Многие прокариоты, так же как эукариоты, могут получать аминокислоты из
Биосинтез аминокислот.
Многие прокариоты, так же как эукариоты, могут получать аминокислоты из

Биосинтез белков.
Основу конструктивного обмена составляет биосинтез белков, который находится под контролем
Биосинтез белков.
Основу конструктивного обмена составляет биосинтез белков, который находится под контролем

Биосинтез липидов.
Липиды микроорганизмов представлены жирными кислотами, фосфолипидами, воском, терпенами, каротиноидами, которые
Биосинтез липидов.
Липиды микроорганизмов представлены жирными кислотами, фосфолипидами, воском, терпенами, каротиноидами, которые

Биосинтез липидов.
Последовательное удлинение этих фрагментов приводит в конечном счете к образованию
Биосинтез липидов.
Последовательное удлинение этих фрагментов приводит в конечном счете к образованию
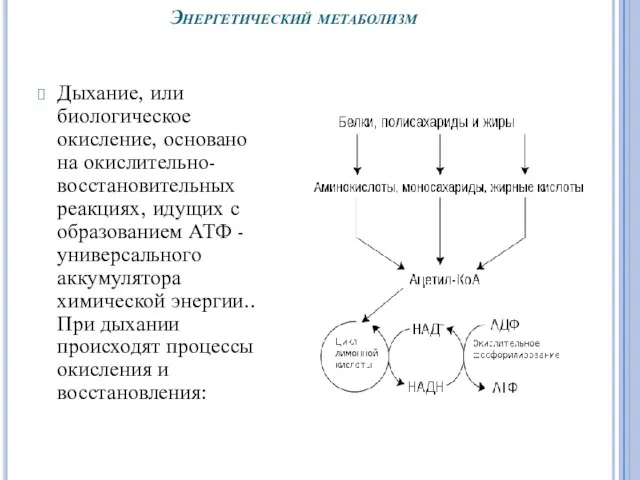
Энергетический метаболизм
Дыхание, или биологическое окисление, основано на окислительно-восстановительных реакциях, идущих с
Энергетический метаболизм
Дыхание, или биологическое окисление, основано на окислительно-восстановительных реакциях, идущих с

Акцептором водорода или электронов может быть молекулярный кислород (такое дыхание называется
Акцептором водорода или электронов может быть молекулярный кислород (такое дыхание называется

Облигатные аэробы могут расти только при наличии кислорода. При наличии кислорода
Облигатные аэробы могут расти только при наличии кислорода. При наличии кислорода

Получение энергии путем субстратного фосфорилирования.
Брожение.
У микроорганизмов выделяют три пути превращения
Получение энергии путем субстратного фосфорилирования.
Брожение.
У микроорганизмов выделяют три пути превращения

Гликолиз (путь Эмдена - Мейергофа).
В результате расщепления глюкозы расходуется 2
Гликолиз (путь Эмдена - Мейергофа).
В результате расщепления глюкозы расходуется 2
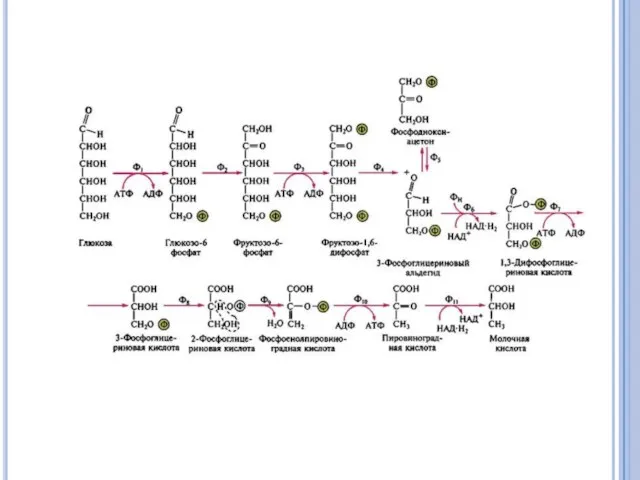

Брожение
Для облигатных и факультативных анаэробов характерно брожение - способом получения
Брожение
Для облигатных и факультативных анаэробов характерно брожение - способом получения
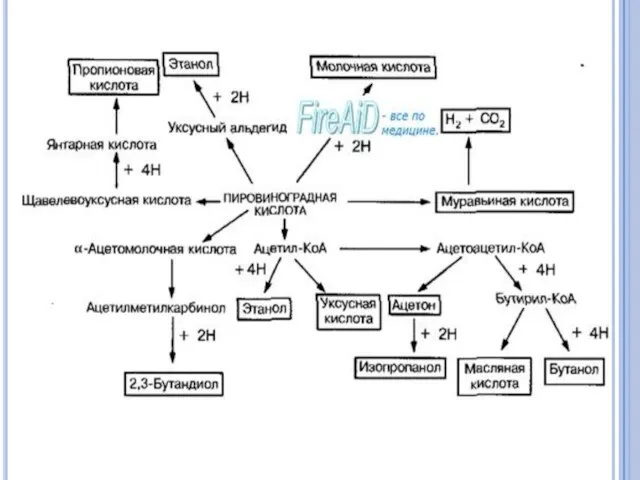

Молочнокислое брожение.
Бактерии родов Lactobacillus, Streptococcus, Bifidobacterium1 способны образовывать из пирувата молочную
Молочнокислое брожение.
Бактерии родов Lactobacillus, Streptococcus, Bifidobacterium1 способны образовывать из пирувата молочную

Маслянокислое брожение.
Одним из основных продуктов брожения является масляная кислота.
При этом
Маслянокислое брожение.
Одним из основных продуктов брожения является масляная кислота.
При этом

Муравьинокислое брожение.
Этот тип брожения характерен для представителей семейства энтеробактерий.
Одним из
Муравьинокислое брожение.
Этот тип брожения характерен для представителей семейства энтеробактерий.
Одним из

Пропионовокислое брожение
характерно для пропионобактерий, которые из пирувата образуют пропионовую кислоту.
Многие бактерии
Пропионовокислое брожение
характерно для пропионобактерий, которые из пирувата образуют пропионовую кислоту.
Многие бактерии

Пентозофосфатный,
или гексозомонофосфатный (ГМФ), путь.
Этот путь расщепления глюкозы характерен для многих
Пентозофосфатный,
или гексозомонофосфатный (ГМФ), путь.
Этот путь расщепления глюкозы характерен для многих

Кетодезоксифосфоглюконатный (КДФГ) путь (Энтнера - Дудорова).
У высших организмов КДФГ-путь отсутствует. Пировиноградная
Кетодезоксифосфоглюконатный (КДФГ) путь (Энтнера - Дудорова).
У высших организмов КДФГ-путь отсутствует. Пировиноградная
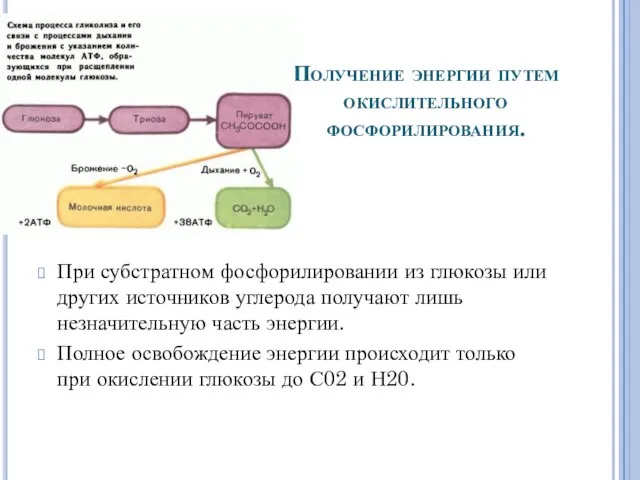
Получение энергии путем
окислительного фосфорилирования.
При субстратном фосфорилировании из глюкозы или других
Получение энергии путем
окислительного фосфорилирования.
При субстратном фосфорилировании из глюкозы или других

.
Пируват, образующийся в процессе гликолиза, окисляется до ацетил-КоА , который
.
Пируват, образующийся в процессе гликолиза, окисляется до ацетил-КоА , который

У всех аэробных и факультативно-анаэробных бактерий дыхательная цепь локализована на цитоплазматической
У всех аэробных и факультативно-анаэробных бактерий дыхательная цепь локализована на цитоплазматической
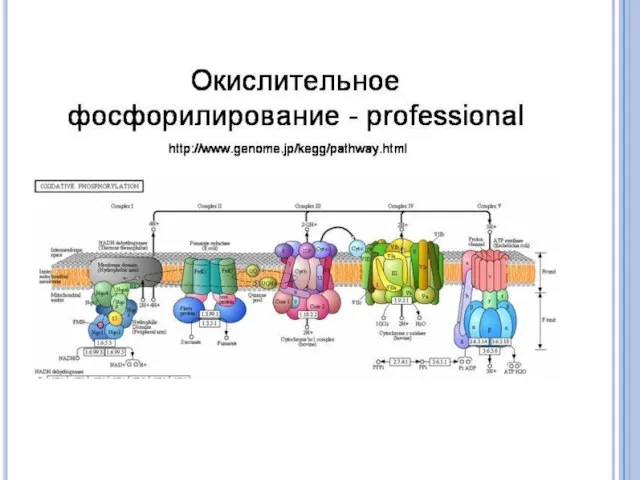

У бактерий распространены системы окисления субстрата, связанные не с цитохромами, а
У бактерий распространены системы окисления субстрата, связанные не с цитохромами, а

Поскольку при окислительном фосфорилировании высвобождается гораздо больше энергии, чем при брожении,
Поскольку при окислительном фосфорилировании высвобождается гораздо больше энергии, чем при брожении,

Рост и размножение бактерий.
Жизнедеятельность бактерий характеризуется
ростом - формированием структурно-функциональных
Рост и размножение бактерий.
Жизнедеятельность бактерий характеризуется
ростом - формированием структурно-функциональных

Бактерии размножаются путем бинарного деления пополам, реже путем почкования. Актиномицеты, как
Бактерии размножаются путем бинарного деления пополам, реже путем почкования. Актиномицеты, как
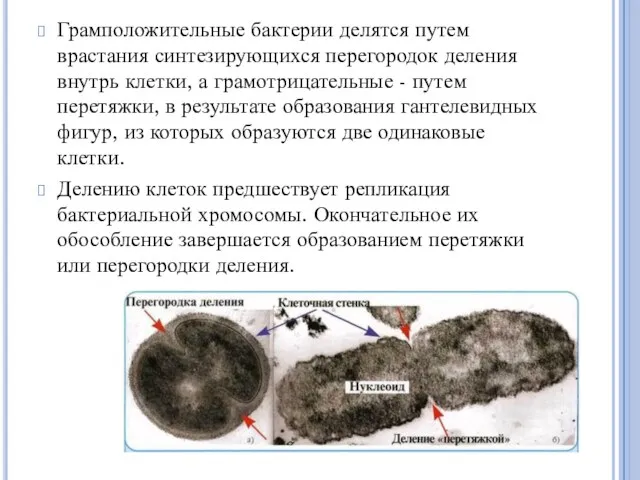
Грамположительные бактерии делятся путем врастания синтезирующихся перегородок деления внутрь клетки, а
Грамположительные бактерии делятся путем врастания синтезирующихся перегородок деления внутрь клетки, а

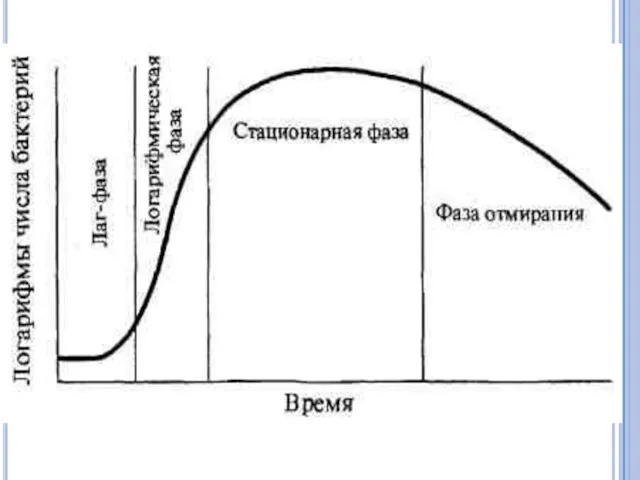
Размножение бактерий в жидкой питательной среде. Бактерии, засеянные в определенный, не
Размножение бактерий в жидкой питательной среде. Бактерии, засеянные в определенный, не


При выращивании бактерий на жидкой питательной среде наблюдается придонный, диффузный или
При выращивании бактерий на жидкой питательной среде наблюдается придонный, диффузный или

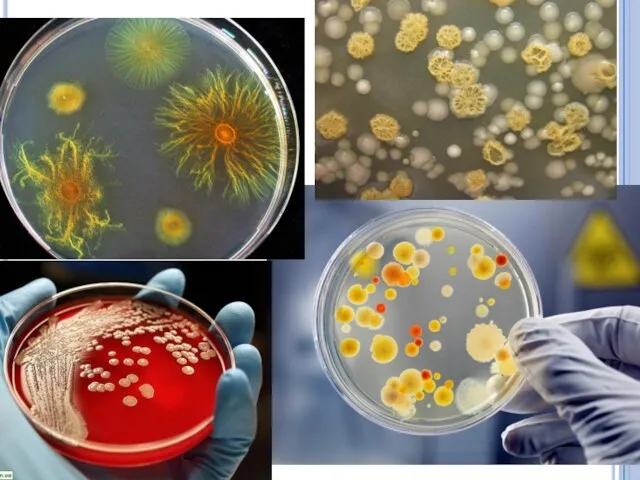
Размножение бактерий
на плотной питательной среде.
Бактерии, растущие на плотных питательных средах,
Размножение бактерий
на плотной питательной среде.
Бактерии, растущие на плотных питательных средах,

Питательные среды
Питательные среды должны обязательно отвечать основным требованиям:
они должны
Питательные среды
Питательные среды должны обязательно отвечать основным требованиям:
они должны

Питательные среды
Существует много различных вариантов питательных сред, сконструированных с учетом
Питательные среды
Существует много различных вариантов питательных сред, сконструированных с учетом

По назначению питательные среды подразделяют на следующие основные категории.
Универсальные
Селективные
По назначению питательные среды подразделяют на следующие основные категории.
Универсальные
Селективные

Универсальные — среды, на которых хорошо растут многие виды патогенных и
Универсальные — среды, на которых хорошо растут многие виды патогенных и

Дифференциально-диагностические — среды, позволяющие отличать одни виды бактерий от других по
Дифференциально-диагностические — среды, позволяющие отличать одни виды бактерий от других по

Селективные (синонимы: избирательные, элективные, обогатительные) - среды, содержащие вещества, используемые микроорганизмами
Селективные (синонимы: избирательные, элективные, обогатительные) - среды, содержащие вещества, используемые микроорганизмами

Дифференциально-селективные — среды, сочетающие в себе свойства дифференциально-диагностических и селективных сред.
Дифференциально-селективные — среды, сочетающие в себе свойства дифференциально-диагностических и селективных сред.

Синтетические — среды строго определенного химического состава, представляющие собой растворы неорганических
Синтетические — среды строго определенного химического состава, представляющие собой растворы неорганических

Специальные — среды, специально приготовленные для получения роста тех бактерий, которые
Специальные — среды, специально приготовленные для получения роста тех бактерий, которые

Синтетические — среды строго определенного химического состава, представляющие собой растворы неорганических
Синтетические — среды строго определенного химического состава, представляющие собой растворы неорганических
 Генетика: основные термины и понятия
Генетика: основные термины и понятия Хомяк джунгарский
Хомяк джунгарский Промышленное культивирование микроорганизмов
Промышленное культивирование микроорганизмов Фізіологія ендокринної системи
Фізіологія ендокринної системи ПРИСПОСОБИТЕЛЬНЫЕ ОСОБЕННОСТИ СТРОЕНИЯ,ОКРАСКИ ТЕЛА И ПОВЕДЕНИЯ ЖИВОТНЫХ.
ПРИСПОСОБИТЕЛЬНЫЕ ОСОБЕННОСТИ СТРОЕНИЯ,ОКРАСКИ ТЕЛА И ПОВЕДЕНИЯ ЖИВОТНЫХ. Meristem and cover tissues. Constant tissues: transport, mechanic
Meristem and cover tissues. Constant tissues: transport, mechanic Учение о тканях
Учение о тканях Плауны. Хвощи. Папоротники
Плауны. Хвощи. Папоротники Тип хордовые. Подтип черепные. Класс земноводные (амфибии
Тип хордовые. Подтип черепные. Класс земноводные (амфибии Биогенные элементы. Классификация. Топография биогенных элементов в организме человека
Биогенные элементы. Классификация. Топография биогенных элементов в организме человека Энергетический и пластический обмен
Энергетический и пластический обмен Спинной мозг
Спинной мозг Мочеполовой аппарат
Мочеполовой аппарат Урок-игротека Животные
Урок-игротека Животные Тұқымқуалаудың хромосомадан тыс факторлары: плазмидалар, транспозондар, isтіркестер. Бактериялар мен вирустардың генетикасы
Тұқымқуалаудың хромосомадан тыс факторлары: плазмидалар, транспозондар, isтіркестер. Бактериялар мен вирустардың генетикасы Посевные качества семян
Посевные качества семян Обобщение по теме Царство Грибы
Обобщение по теме Царство Грибы Что нужно знать, чтобы вырастить различные виды растений?
Что нужно знать, чтобы вырастить различные виды растений? Задачи по генетике: дигибридное скрещивание
Задачи по генетике: дигибридное скрещивание Рефлексы
Рефлексы Иммуноферментный анализ (ИФА). Иммуноблоттинг
Иммуноферментный анализ (ИФА). Иммуноблоттинг Половое размножение животных
Половое размножение животных Белки. Переваривание и всасывание
Белки. Переваривание и всасывание Основы цитологии. Понятие о клетке
Основы цитологии. Понятие о клетке Общие свойства сенсорных систем. Болевая сенсорная система
Общие свойства сенсорных систем. Болевая сенсорная система Игра Что? Где? Когда?
Игра Что? Где? Когда? Дидактическая игра Самые умные
Дидактическая игра Самые умные Большой год 2019 областной фотоконкурс
Большой год 2019 областной фотоконкурс