- Физиология мышечного сокращения

Содержание
- 2. Ультраструктура скелетной мышцы. Поперечно-полосатая мышца состоит из множества функциональных единиц – мышечных волокон или мышечных клеток.
- 3. Молекула миозина состоит из длинного участка – хвоста и глобулярной части из двух головок. Молекулы миозина
- 4. Тропонин участвует в Са2+ - зависимой регуляции сокращения. Тропонин Т имеет участок для связывания тропомиозина и
- 5. Типы мышечных волокон. По способу ответа на возбуждающие нервные импульсы скелетные поперечно-полосатые мышцы подразделяют на тонические
- 6. Фазные волокна по скорости развития напряжения подразделяют на «медленные» – волокна I типа и «быстрые» –
- 7. 4-2. СОВРЕМЕННАЯ ТЕОРИЯ МЫШЕЧНОГО СОКРАЩЕНИЯ И РАССЛАБЛЕНИЯ. ЭЛЕКТРОМЕХАНИЧЕСКОЕ СОПРЯЖЕНИЕ. Механизм мышечного сокращения. В присутствии АТФ, но
- 8. Цикл после завершения может повториться или остановиться. Если бы все поперечные мостики в отдельной мышце прошли
- 9. В результате циклов поперечных мостиков саркомер укорачивается, и концы мышцы подтягиваются к ее центру. Прочная соединительнотканная
- 10. Остановка сокращения. Сигналом к остановке сокращения скелетной мышцы служит прекращение нервного импульса в двигательной концевой пластинке
- 11. 4-3. ВИДЫ И РЕЖИМЫ МЫШЕЧНОГО СОКРАЩЕНИЯ. ОДИНОЧНОЕ СОКРАЩЕНИЕ. ПОНЯТИЕ О ТЕТАНУСЕ. МЕХАНИЗМ ДЛИТЕЛЬНОГО СОКРАЩЕНИЯ В ЕСТЕСТВЕННЫХ
- 12. Одиночное сокращение и тетанус. Одиночный стимул спустя короткое время, около 50 мс, называемое латентным периодом, вызывает
- 13. Если следующий нервный импульс приходит к волокну раньше, чем за-кончилось расслабление предыдущего одиночного сокращения, происходит суперпозиция
- 14. Сила сокращения скелетных мышц. Двигательная единица. Под двигательной единицей понимают каждую группу мышечных волокон вместе с
- 15. Максимальная сила мышц. Абсолютная сила, развиваемая разными мышцами, неодинакова. Она определяется: 1) поперечным сечением мышцы, 2)
- 16. 4.4. ФИЗИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ И СВОЙСТВА гладких мышц. Классификация гладких мышц. Висцеральные гладкие мышцы находятся во всех
- 17. Иннервация гладких мышц. Висцеральная гладкая мышца имеет двойную иннервацию — симпатическую и парасимпатическую, функция кото-рой заключается
- 18. Нервно-мышечная передача. Возбуждающее влияние адренергических или холинергических нервов электрически проявляется в виде отдельных волн деполяризации. При
- 19. Автоматия. Многим гладким мышцам свойственна спонтанная, автоматическая активность. Для нее характерно медленное снижение мембранного потенциала покоя,
- 20. Механизм сокращения. В механизме сокращения гладкой мышцы имеется особенность, отличающая его от механизма сокращения скелетной мышцы.
- 21. Проведение возбуждения по гладкой мышце. В отличие от того, что имеет место в скелетных мышцах, в
- 23. Скачать презентацию

Ультраструктура скелетной мышцы. Поперечно-полосатая мышца состоит из множества функциональных единиц –
Ультраструктура скелетной мышцы. Поперечно-полосатая мышца состоит из множества функциональных единиц –

Молекула миозина состоит из длинного участка – хвоста и глобулярной части
Молекула миозина состоит из длинного участка – хвоста и глобулярной части

Тропонин участвует в Са2+ - зависимой регуляции сокращения. Тропонин Т имеет
Тропонин участвует в Са2+ - зависимой регуляции сокращения. Тропонин Т имеет

Типы мышечных волокон. По способу ответа на возбуждающие нервные импульсы скелетные
Типы мышечных волокон. По способу ответа на возбуждающие нервные импульсы скелетные

Фазные волокна по скорости развития напряжения подразделяют на «медленные» – волокна
Фазные волокна по скорости развития напряжения подразделяют на «медленные» – волокна

4-2. СОВРЕМЕННАЯ ТЕОРИЯ МЫШЕЧНОГО СОКРАЩЕНИЯ И РАССЛАБЛЕНИЯ. ЭЛЕКТРОМЕХАНИЧЕСКОЕ СОПРЯЖЕНИЕ.
Механизм мышечного
4-2. СОВРЕМЕННАЯ ТЕОРИЯ МЫШЕЧНОГО СОКРАЩЕНИЯ И РАССЛАБЛЕНИЯ. ЭЛЕКТРОМЕХАНИЧЕСКОЕ СОПРЯЖЕНИЕ.
Механизм мышечного

Цикл после завершения может повториться или остановиться. Если бы все поперечные
Цикл после завершения может повториться или остановиться. Если бы все поперечные

В результате циклов поперечных мостиков саркомер укорачивается, и концы мышцы подтягиваются
В результате циклов поперечных мостиков саркомер укорачивается, и концы мышцы подтягиваются

Остановка сокращения.
Сигналом к остановке сокращения скелетной мышцы служит прекращение нервного
Остановка сокращения.
Сигналом к остановке сокращения скелетной мышцы служит прекращение нервного

4-3. ВИДЫ И РЕЖИМЫ МЫШЕЧНОГО СОКРАЩЕНИЯ. ОДИНОЧНОЕ СОКРАЩЕНИЕ. ПОНЯТИЕ О ТЕТАНУСЕ.
4-3. ВИДЫ И РЕЖИМЫ МЫШЕЧНОГО СОКРАЩЕНИЯ. ОДИНОЧНОЕ СОКРАЩЕНИЕ. ПОНЯТИЕ О ТЕТАНУСЕ.
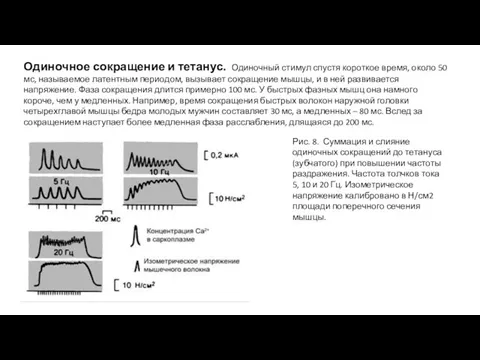
Одиночное сокращение и тетанус. Одиночный стимул спустя короткое время, около 50
Одиночное сокращение и тетанус. Одиночный стимул спустя короткое время, около 50

Если следующий нервный импульс приходит к волокну раньше, чем за-кончилось расслабление
Если следующий нервный импульс приходит к волокну раньше, чем за-кончилось расслабление

Сила сокращения скелетных мышц.
Двигательная единица. Под двигательной единицей понимают каждую
Сила сокращения скелетных мышц.
Двигательная единица. Под двигательной единицей понимают каждую

Максимальная сила мышц.
Абсолютная сила, развиваемая разными мышцами, неодинакова. Она определяется:
Максимальная сила мышц.
Абсолютная сила, развиваемая разными мышцами, неодинакова. Она определяется:

4.4. ФИЗИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ И СВОЙСТВА гладких мышц.
Классификация гладких мышц. Висцеральные
4.4. ФИЗИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ И СВОЙСТВА гладких мышц.
Классификация гладких мышц. Висцеральные

Иннервация гладких мышц.
Висцеральная гладкая мышца имеет двойную иннервацию — симпатическую
Иннервация гладких мышц.
Висцеральная гладкая мышца имеет двойную иннервацию — симпатическую

Нервно-мышечная передача. Возбуждающее влияние адренергических или холинергических нервов электрически проявляется в
Нервно-мышечная передача. Возбуждающее влияние адренергических или холинергических нервов электрически проявляется в

Автоматия. Многим гладким мышцам свойственна спонтанная, автоматическая активность. Для нее характерно
Автоматия. Многим гладким мышцам свойственна спонтанная, автоматическая активность. Для нее характерно

Механизм сокращения. В механизме сокращения гладкой мышцы имеется особенность, отличающая его
Механизм сокращения. В механизме сокращения гладкой мышцы имеется особенность, отличающая его

Проведение возбуждения по гладкой мышце. В отличие от того, что имеет
Проведение возбуждения по гладкой мышце. В отличие от того, что имеет
 Размножение хвойных растений черенкованием
Размножение хвойных растений черенкованием Происхождение человека. Семинар-дискуссия
Происхождение человека. Семинар-дискуссия Разные виды растительных клеток. 5 класс
Разные виды растительных клеток. 5 класс Царство бактерии. Особенности строения и жизнедеятельности
Царство бактерии. Особенности строения и жизнедеятельности Нервная система
Нервная система Индивидуальные параметры тела Wellness -Test
Индивидуальные параметры тела Wellness -Test Ознакомление с фитонцидными растениями и выявление возможности их использования в интерьере. Практическая работа
Ознакомление с фитонцидными растениями и выявление возможности их использования в интерьере. Практическая работа Жизнь в океане
Жизнь в океане Декоративное оформление дворянской усадьбы XIII -XIX веков
Декоративное оформление дворянской усадьбы XIII -XIX веков Класс Земноводные или Амфибии
Класс Земноводные или Амфибии Микроскопическое строение крови человека и лягушки. Лабораторная работа 3
Микроскопическое строение крови человека и лягушки. Лабораторная работа 3 Многообразие организмов. Значение работ К. Линнея и Ж.Б. Ламарка. Основные систематические категории
Многообразие организмов. Значение работ К. Линнея и Ж.Б. Ламарка. Основные систематические категории Двойное оплодотворение у цветковых растений
Двойное оплодотворение у цветковых растений Жасуша ядросы
Жасуша ядросы Животные-рекордсмены
Животные-рекордсмены Строение и жизнедеятельность бактерий
Строение и жизнедеятельность бактерий Зелёная химия
Зелёная химия Китообразные. Ластоногие. Хоботные. Хищники
Китообразные. Ластоногие. Хоботные. Хищники Мозг: как он устроен и работает
Мозг: как он устроен и работает Грибы и их многообразие
Грибы и их многообразие Морфология и ультраструктура бактерий
Морфология и ультраструктура бактерий Красота окружающего мира. Насекомые
Красота окружающего мира. Насекомые Рост и развитие растений
Рост и развитие растений Гербициды. (Лекция 8)
Гербициды. (Лекция 8) Черенкование комнатных растений
Черенкование комнатных растений Нуклеиновые кислоты. 9 класс
Нуклеиновые кислоты. 9 класс чуть больше о Воде
чуть больше о Воде Митоз и мейоз
Митоз и мейоз