Физиология мышц. Структурно-функциональная характеристика скелетной мышцы и механизм ее сокращения презентация
- Физиология мышц. Структурно-функциональная характеристика скелетной мышцы и механизм ее сокращения

Содержание
- 2. Структурной единицей скелетной мышцы является мышечное волокно - сильно вытянутая многоядерная клетка. Длина мышечного волокна зависит
- 3. Скелетные мышцы прикреплены в основном к костям, что и обусловило их название. Сокращение скелетных мышц инициируется
- 4. Мышечное волокно окружено двухслойной липопротеидной электровозбудимой мембраной - сарколеммой, которая покрыта сетью коллагеновых волокон, придающих ей
- 5. Строение мышечного волокна Сарколемма – плазматическая мембрана покрывающая мышечное волокно (соединяется с сухожилием, которое прикрепляет мышцу
- 6. Система поперечных трубочек. Это сеть Т – трубочек (поперечные), является продолжением сарколеммы; они взаимосоединяются проходя среди
- 7. Структура миофибриллы: 1. Перегородки – называемые Z - пластинками, разделяют их на саркомеры. Структура саркомера: В
- 8. Актиновые нити прикрепляются к Z – пластинкам, сами Z – пластинки ограничивают саркомер. В покоящейся мышце
- 9. Теория скользящих нитей. Укорочение саркомера: Мышца сокращается в результате укорочения множества последовательно соединенных саркомеров в миофибриллах.
- 10. Работа поперечных мостиков. Движение головок создает объединенное усилие, как бы «гребок», продвигающий актиновые нити к середине
- 11. Преобразование химической энергии в механическую. АТФ – непосредственный источник энергии для сокращения. При сокращении мышцы АТФ
- 12. Молекулярный механизм мышечного сокращения. Сокращение запускается нервным импульсом. При этом в синапсе - месте контакта нервного
- 13. Электромеханическое сопряжение Передача команды к сокращению от возбужденной клеточной мембраны к миофибриллам в глубине клетки (электромеханическое
- 14. Электромеханическое сопряжение происходит посредством распространения потенциала действия по мембранам поперечной системы внутрь клетки, потом возбуждение проходит
- 15. Функциональная роль АТФ : - в покоящейся мышце - препятствует соединению актиновых нитей с миозиновыми; -
- 16. Типы мышечных сокращений. Оптимум и пессимум мышечного сокращения В зависимости от изменения длины мышечного волокна выделяют
- 17. Мышечная работа равна произведению расстояния (укорочения мышцы) на вес груза, который поднимает мышца. При изотонической тетанической
- 18. Расслабленная мышца, сохраняющая «длину покоя» за счет фиксации обоих ее концов, не развивает силу, которая передавалась
- 19. Пиковое усилие при таких условиях называется максимумом изометрического сокращения. При сильном растяжении мышцы, сила сокращения уменьшается
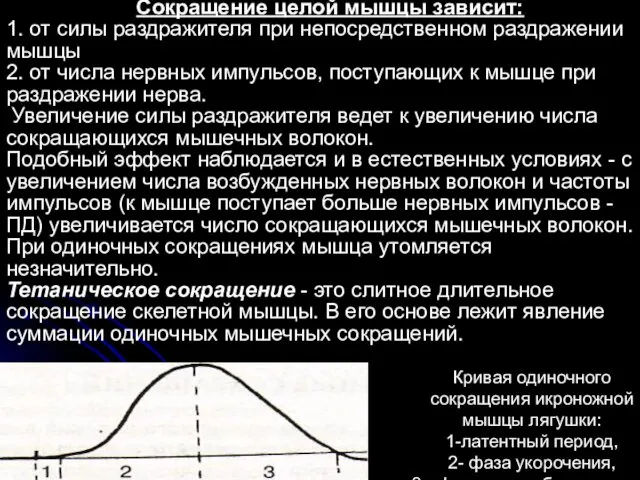
- 20. В зависимости от длительности выделяют одиночное и тетаническое сокращения мышцы. Одиночное сокращение мышцы в эксперименте вызывают
- 21. Сокращение целой мышцы зависит: 1. от силы раздражителя при непосредственном раздражении мышцы 2. от числа нервных
- 22. При нанесении на мышечное волокно или непосредственно на мышцу двух быстро следующих друг за другом раздражений
- 23. При поступлении импульсов к мышце во время ее расслабления возникает зубчатый тетанус, во время укорочения -
- 24. Рис. Сокращения икроножной мышцы лягушки при увеличении частоты раздражения седалищного нерва (ст/с — стимулов в секунду):
- 25. При чрезмерно частой стимуляции нерва (более 100 имп/с) мышца расслабляется вследствие блока проведения возбуждения в нервно-мышечных
- 26. Однако форма сокращения мышцы в целом напоминает гладкий тетанус. Причины этого - асинхронность разрядов мотонейронов и
- 27. Функциональная единица мышцы – двигательная единица Понятия. Иннервация скелетных мышечных волокон осуществляется мотонейронами спинного мозга или
- 28. При сокращении мышцы в натуральных (естественных) условиях можно зарегистрировать ее электрическую активность (электромиограмму - ЭМГ) с
- 29. Медленные двигательные единицы включают медленные мотонейроны и медленные мышечные волокна (красные). Медленные мотонейроны, как правило, низкопороговые,
- 30. Быстрые двигательные единицы состоят из быстрых мотонейронов и быстрых мышечных волокон. Быстрые высокопороговые мотонейроны включаются в
- 31. Быстрые мышечные волокна (белые мышечные волокна) более толстые, содержат больше миофибрилл, обладают большей силой, чем медленные
- 32. Скорость сокращения мышечных волокон находится в прямой зависимости от активности миозин-АТФ-азы - фермента, расщепляющего АТФ и
- 33. КПД мышцы Во время активации мышцы повышение внутриклеточной концентрации Са 2+ ведет к сокращению и к
- 34. Коэффициент полезного действия. Гидролиз одного моля АТФ дает 48 кДж энергии, 40 –50% - превращается в
- 35. Энергетический метаболизм . Во время продолжительной равномерной мышечной активности происходит аэробная регенерация АТФ за счет окислительного
- 36. Анаэробное расщепление – гликолиз: АТФ образуется в 2-3 раза быстрее, а механическая энергия мышцы в 2-3
- 37. Соотношение быстрых, и медленных волокон. Чем больше быстрых волокон содержит мышца, тем больше возможная ее сила
- 38. В различных мышцах тела соотношение между числом медленных и быстрых мышечных волокон неодинаково, поэтому и сила
- 39. Умеренное растяжение мышцы также ведет к увеличению ее сократительного эффекта. Однако при чрезмерном растяжении сила сокращения
- 40. Функциональное состояние мышцы. При утомлении мышцы величина ее сокращения снижается. Работа мышцы измеряется произведением поднятого груза
- 41. ГЛАДКАЯ МЫШЦА. Гладкая мускулатура не имеет поперечную исчерченность. Клетки в виде веретен соединены особыми межклеточными контактами
- 43. Скачать презентацию

Структурной единицей скелетной мышцы является мышечное волокно - сильно вытянутая многоядерная
Структурной единицей скелетной мышцы является мышечное волокно - сильно вытянутая многоядерная

Скелетные мышцы прикреплены в основном к костям, что и обусловило их
Скелетные мышцы прикреплены в основном к костям, что и обусловило их

Мышечное волокно окружено двухслойной липопротеидной электровозбудимой мембраной - сарколеммой, которая покрыта
Мышечное волокно окружено двухслойной липопротеидной электровозбудимой мембраной - сарколеммой, которая покрыта

Строение мышечного волокна
Сарколемма – плазматическая мембрана покрывающая мышечное волокно (соединяется с
Строение мышечного волокна
Сарколемма – плазматическая мембрана покрывающая мышечное волокно (соединяется с

Система поперечных трубочек. Это сеть Т – трубочек (поперечные), является продолжением
Система поперечных трубочек. Это сеть Т – трубочек (поперечные), является продолжением

Структура миофибриллы:
1. Перегородки – называемые Z - пластинками, разделяют их
1. Перегородки – называемые Z - пластинками, разделяют их

Актиновые нити прикрепляются к Z – пластинкам, сами Z – пластинки
Актиновые нити прикрепляются к Z – пластинкам, сами Z – пластинки

Теория скользящих нитей.
Укорочение саркомера:
Мышца сокращается в результате укорочения множества последовательно соединенных
Теория скользящих нитей.
Укорочение саркомера:
Мышца сокращается в результате укорочения множества последовательно соединенных

Работа поперечных мостиков.
Движение головок создает объединенное усилие, как бы «гребок», продвигающий
Работа поперечных мостиков.
Движение головок создает объединенное усилие, как бы «гребок», продвигающий

Преобразование химической энергии в механическую.
АТФ – непосредственный источник энергии для сокращения.
При
Преобразование химической энергии в механическую.
АТФ – непосредственный источник энергии для сокращения.
При

Молекулярный механизм мышечного сокращения.
Сокращение запускается нервным импульсом. При этом в синапсе
Молекулярный механизм мышечного сокращения.
Сокращение запускается нервным импульсом. При этом в синапсе

Электромеханическое сопряжение
Передача команды к сокращению от возбужденной клеточной мембраны к
Электромеханическое сопряжение
Передача команды к сокращению от возбужденной клеточной мембраны к

Электромеханическое сопряжение происходит посредством распространения потенциала действия по мембранам поперечной системы
Электромеханическое сопряжение происходит посредством распространения потенциала действия по мембранам поперечной системы

Функциональная роль АТФ :
- в покоящейся мышце - препятствует соединению
Функциональная роль АТФ :
- в покоящейся мышце - препятствует соединению

Типы мышечных сокращений. Оптимум и пессимум мышечного сокращения
В зависимости от
Типы мышечных сокращений. Оптимум и пессимум мышечного сокращения
В зависимости от

Мышечная работа равна произведению расстояния (укорочения мышцы) на вес груза, который
Мышечная работа равна произведению расстояния (укорочения мышцы) на вес груза, который

Расслабленная мышца, сохраняющая «длину покоя» за счет фиксации обоих ее концов,
Расслабленная мышца, сохраняющая «длину покоя» за счет фиксации обоих ее концов,

Пиковое усилие при таких условиях называется максимумом изометрического сокращения.
При сильном
Пиковое усилие при таких условиях называется максимумом изометрического сокращения.
При сильном

В зависимости от длительности выделяют одиночное и тетаническое сокращения мышцы.
Одиночное сокращение
В зависимости от длительности выделяют одиночное и тетаническое сокращения мышцы.
Одиночное сокращение
Сокращение целой мышцы зависит:
1. от силы раздражителя при непосредственном раздражении
Сокращение целой мышцы зависит:
1. от силы раздражителя при непосредственном раздражении

При нанесении на мышечное волокно или непосредственно на мышцу двух быстро
При нанесении на мышечное волокно или непосредственно на мышцу двух быстро

При поступлении импульсов к мышце во время ее расслабления возникает зубчатый
При поступлении импульсов к мышце во время ее расслабления возникает зубчатый
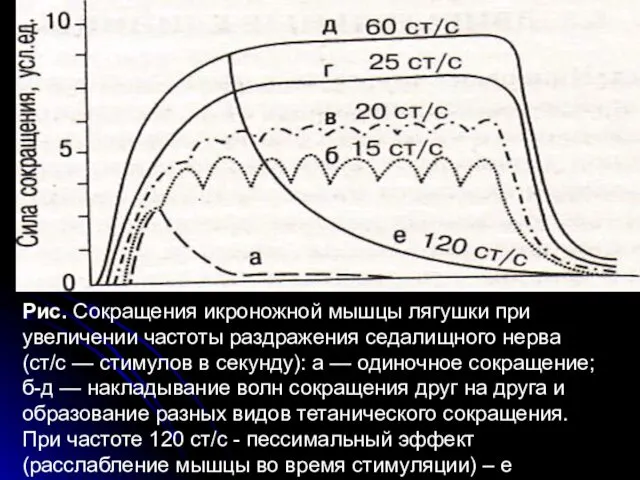
Рис. Сокращения икроножной мышцы лягушки при увеличении частоты раздражения седалищного нерва
Рис. Сокращения икроножной мышцы лягушки при увеличении частоты раздражения седалищного нерва

При чрезмерно частой стимуляции нерва (более 100 имп/с) мышца расслабляется вследствие
При чрезмерно частой стимуляции нерва (более 100 имп/с) мышца расслабляется вследствие

Однако форма сокращения мышцы в целом напоминает гладкий тетанус.
Причины этого
Однако форма сокращения мышцы в целом напоминает гладкий тетанус.
Причины этого

Функциональная единица мышцы – двигательная единица
Понятия. Иннервация скелетных мышечных волокон
Функциональная единица мышцы – двигательная единица
Понятия. Иннервация скелетных мышечных волокон

При сокращении мышцы в натуральных (естественных) условиях можно зарегистрировать ее электрическую
При сокращении мышцы в натуральных (естественных) условиях можно зарегистрировать ее электрическую

Медленные двигательные единицы включают медленные мотонейроны и медленные мышечные волокна (красные).
Медленные двигательные единицы включают медленные мотонейроны и медленные мышечные волокна (красные).

Быстрые двигательные единицы состоят из быстрых мотонейронов и быстрых мышечных волокон.
Быстрые двигательные единицы состоят из быстрых мотонейронов и быстрых мышечных волокон.

Быстрые мышечные волокна (белые мышечные волокна) более толстые, содержат больше миофибрилл,
Быстрые мышечные волокна (белые мышечные волокна) более толстые, содержат больше миофибрилл,

Скорость сокращения мышечных волокон находится в прямой зависимости от активности миозин-АТФ-азы
Скорость сокращения мышечных волокон находится в прямой зависимости от активности миозин-АТФ-азы

КПД мышцы
Во время активации мышцы повышение внутриклеточной концентрации Са 2+
КПД мышцы
Во время активации мышцы повышение внутриклеточной концентрации Са 2+

Коэффициент полезного действия.
Гидролиз одного моля АТФ дает 48 кДж энергии,
Коэффициент полезного действия.
Гидролиз одного моля АТФ дает 48 кДж энергии,

Энергетический метаболизм . Во время продолжительной равномерной мышечной активности происходит аэробная
Энергетический метаболизм . Во время продолжительной равномерной мышечной активности происходит аэробная

Анаэробное расщепление – гликолиз: АТФ образуется в 2-3 раза быстрее, а
Анаэробное расщепление – гликолиз: АТФ образуется в 2-3 раза быстрее, а

Соотношение быстрых, и медленных волокон. Чем больше быстрых волокон содержит мышца,
Соотношение быстрых, и медленных волокон. Чем больше быстрых волокон содержит мышца,

В различных мышцах тела соотношение между числом медленных и быстрых мышечных
В различных мышцах тела соотношение между числом медленных и быстрых мышечных

Умеренное растяжение мышцы также ведет к увеличению ее сократительного эффекта. Однако
Умеренное растяжение мышцы также ведет к увеличению ее сократительного эффекта. Однако

Функциональное состояние мышцы.
При утомлении мышцы величина ее сокращения снижается.
Работа
Функциональное состояние мышцы.
При утомлении мышцы величина ее сокращения снижается.
Работа

ГЛАДКАЯ МЫШЦА.
Гладкая мускулатура не имеет поперечную исчерченность. Клетки в виде веретен
ГЛАДКАЯ МЫШЦА.
Гладкая мускулатура не имеет поперечную исчерченность. Клетки в виде веретен
 Науки о человеке: анатомия, физиология, гигиена
Науки о человеке: анатомия, физиология, гигиена Биология оқыту әдістемесі ғылым ретінде. Биология оқыту әдістемесінің пәні
Биология оқыту әдістемесі ғылым ретінде. Биология оқыту әдістемесінің пәні Грегор Мендель. Закон единообразия гибридов
Грегор Мендель. Закон единообразия гибридов Микроскопические методы исследования морфологии микроорганизмов
Микроскопические методы исследования морфологии микроорганизмов Этапы эволюции человека
Этапы эволюции человека Как определить птицу
Как определить птицу Пути воздействия организмов на среду обитания
Пути воздействия организмов на среду обитания Організмовий рівень організації генетичної інформації. Взаємодія генів
Організмовий рівень організації генетичної інформації. Взаємодія генів Продуктивность кукурузы в зависимости от приемов выращивания в центральной зоне Краснодарского края
Продуктивность кукурузы в зависимости от приемов выращивания в центральной зоне Краснодарского края Сердечно-сосудистая система
Сердечно-сосудистая система Презентация по биологии Семейство крестоцветные
Презентация по биологии Семейство крестоцветные Бактериофаги. Строение бактериофагов
Бактериофаги. Строение бактериофагов Дикие и домашние животные
Дикие и домашние животные По грибы. Классный час, возраст: 7-10 лет
По грибы. Классный час, возраст: 7-10 лет Двигательные системы
Двигательные системы Индивидуальное развитие организмов (онтогенез)
Индивидуальное развитие организмов (онтогенез) Урок 8 класс. Осанка. Первая помощь при травмах скелета
Урок 8 класс. Осанка. Первая помощь при травмах скелета Характеристика класса птицы. Внешнее строение птиц
Характеристика класса птицы. Внешнее строение птиц Конспект открытого урока по теме : Опорно-двигательная система. Биология 8 класс.
Конспект открытого урока по теме : Опорно-двигательная система. Биология 8 класс. Царство животные
Царство животные Життя риб навесні. 2 клас
Життя риб навесні. 2 клас Анатомия женских половых органов (1)
Анатомия женских половых органов (1) V.A. Motsart hayoti va ijodi
V.A. Motsart hayoti va ijodi Гиповитаминозы животных
Гиповитаминозы животных Отряды насекомых
Отряды насекомых Презентация непрямой онтогенез
Презентация непрямой онтогенез В поисках витамина С. 2 класс
В поисках витамина С. 2 класс Гіпотези виникнення життя на землі
Гіпотези виникнення життя на землі