- Физиология нервной системы (часть 3)

Содержание
- 2. Соматические функции — восприятие раздражения и осуществление двигательных реакций с участием скелетных мышц. Вегетативные функции —
- 3. Особенности рефлекторной дуги вегетативного рефлекса, функции вегетативных ганглиев Эфферентная часть рефлекторной дуги двухнейронная. Первый нейрон расположен
- 5. Общая схема регуляции вегетативных функций Общая схема обеспечения эфферентных вегетативных реакций предполагает возникновение побуждения, замысла действия,
- 6. Структурное обеспечение регуляции вегетативных функций А. НИЗШИЕ ВЕГЕТАТИВНЫЕ ЦЕНТРЫ Спинной мозг. Вегетативные нейроны заложены в боковых
- 7. В.ГИПОТАЛАМУС как высший подкорковый центр программного обеспечения вегетативных функций Афферентные входы связывают гипоталамус с лимбической системой,
- 8. С. ЛИМБИЧЕСКАЯ СИСТЕМА И НОВАЯ КОРА В РЕГУЛЯЦИИ ВЕГЕТАТИВНЫХ ФУНКЦИЙ Лимбическая система -структуры коры (гиппокамп, парагиппокампова
- 9. Общий план строения афферентной системы, специфическая и неспецифическая системы В ЦНС афферентная импульсация от рецепторов перерабатывается
- 11. Афферентная система :лемнисковая (специфическая) и экстралемнисковая (неспецифическая). Специфическая афферентная система включает в себя задний столб спинного
- 12. Афферентные системы спинного мозга Афференты спинного мозга входят через задние корешки. В спинном мозге они располагаются
- 13. ВОСХОДЯЩИЕ ПУТИ СПИННОГО МОЗГА Задние столбы и ядра задних столбов. Задний столб состоит из прямых коллатералей
- 14. Соматосенсорные функции ствола мозга От ствола мозга отходит большая часть черепномозговых нервов. Он содержит множество отдельных
- 15. Соматосенсорные функции ретикулярной формации Представляет собой важную структуру неспецифической афферентной системы. К ней поступает информация от
- 17. Таламус как коллектор афферентных путей Является важнейшим подкорковым центром обработки информации. На его уровне происходит ее
- 18. Соматосенсорная кора Получает информацию от специфических ядер таламуса. Зрительная и слуховая кора получает информацию от медиального
- 20. Скачать презентацию

Соматические функции — восприятие раздражения и осуществление двигательных реакций с участием
Соматические функции — восприятие раздражения и осуществление двигательных реакций с участием

Особенности рефлекторной дуги вегетативного рефлекса, функции вегетативных ганглиев
Эфферентная часть рефлекторной
Особенности рефлекторной дуги вегетативного рефлекса, функции вегетативных ганглиев
Эфферентная часть рефлекторной
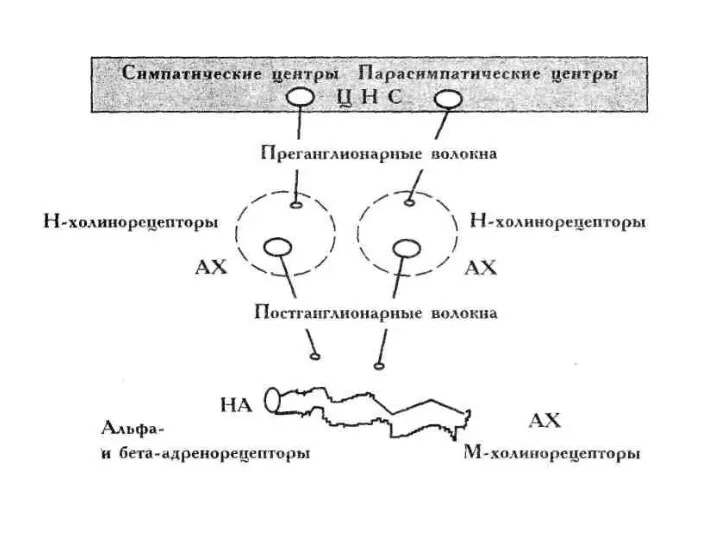

Общая схема регуляции вегетативных функций
Общая схема обеспечения эфферентных вегетативных реакций
Общая схема регуляции вегетативных функций
Общая схема обеспечения эфферентных вегетативных реакций

Структурное обеспечение регуляции вегетативных функций
А. НИЗШИЕ ВЕГЕТАТИВНЫЕ ЦЕНТРЫ
Спинной мозг. Вегетативные
Структурное обеспечение регуляции вегетативных функций
А. НИЗШИЕ ВЕГЕТАТИВНЫЕ ЦЕНТРЫ
Спинной мозг. Вегетативные

В.ГИПОТАЛАМУС как высший подкорковый центр программного обеспечения вегетативных функций
Афферентные входы связывают
В.ГИПОТАЛАМУС как высший подкорковый центр программного обеспечения вегетативных функций
Афферентные входы связывают

С. ЛИМБИЧЕСКАЯ СИСТЕМА И НОВАЯ КОРА В РЕГУЛЯЦИИ
ВЕГЕТАТИВНЫХ ФУНКЦИЙ
Лимбическая система
С. ЛИМБИЧЕСКАЯ СИСТЕМА И НОВАЯ КОРА В РЕГУЛЯЦИИ
ВЕГЕТАТИВНЫХ ФУНКЦИЙ
Лимбическая система

Общий план строения афферентной системы, специфическая и неспецифическая системы
В ЦНС афферентная
Общий план строения афферентной системы, специфическая и неспецифическая системы
В ЦНС афферентная
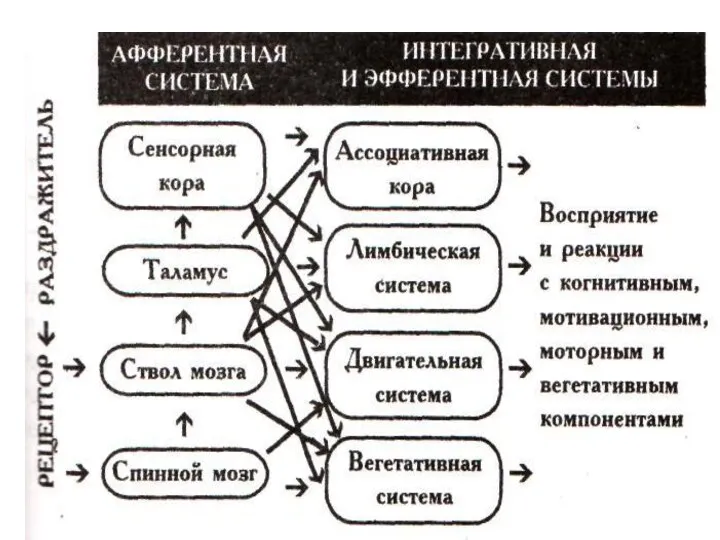

Афферентная система :лемнисковая (специфическая) и экстралемнисковая (неспецифическая).
Специфическая афферентная система включает в
Афферентная система :лемнисковая (специфическая) и экстралемнисковая (неспецифическая).
Специфическая афферентная система включает в
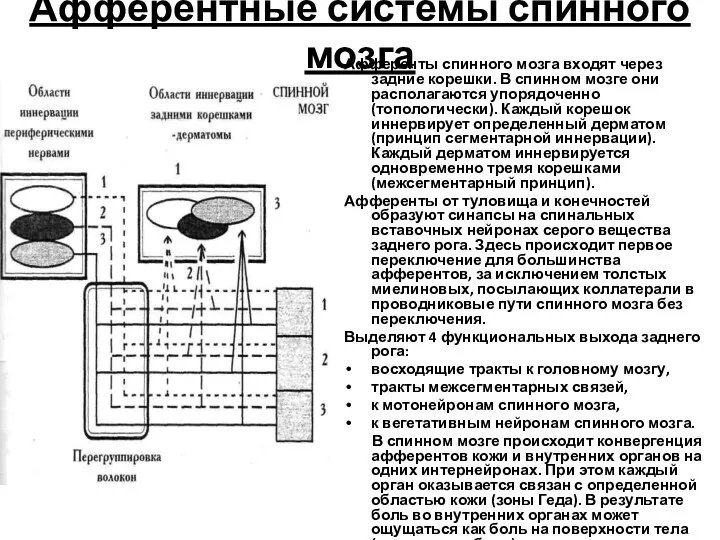
Афферентные системы спинного мозга
Афференты спинного мозга входят через задние корешки. В
Афферентные системы спинного мозга
Афференты спинного мозга входят через задние корешки. В

ВОСХОДЯЩИЕ ПУТИ СПИННОГО МОЗГА
Задние столбы и ядра задних столбов.
Задний столб состоит
ВОСХОДЯЩИЕ ПУТИ СПИННОГО МОЗГА
Задние столбы и ядра задних столбов.
Задний столб состоит

Соматосенсорные функции ствола мозга
От ствола мозга отходит большая часть черепномозговых
Соматосенсорные функции ствола мозга
От ствола мозга отходит большая часть черепномозговых

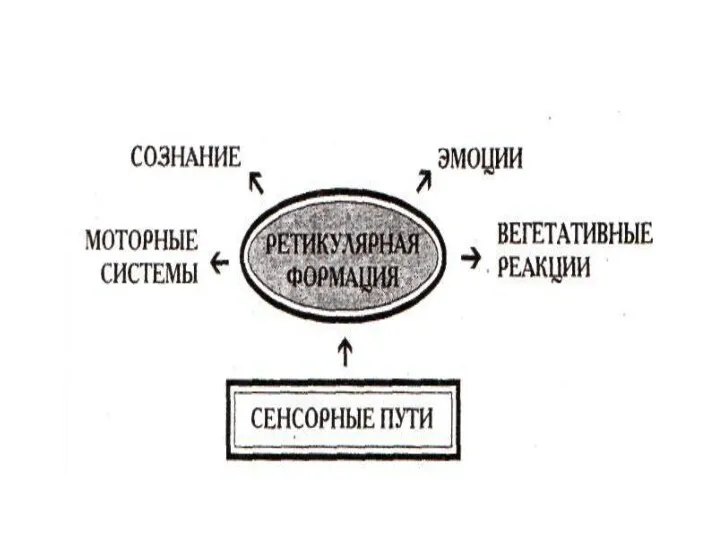
Соматосенсорные функции ретикулярной формации
Представляет собой важную структуру неспецифической афферентной системы. К
Соматосенсорные функции ретикулярной формации
Представляет собой важную структуру неспецифической афферентной системы. К

Таламус как коллектор афферентных путей
Является важнейшим подкорковым центром обработки информации. На
Таламус как коллектор афферентных путей
Является важнейшим подкорковым центром обработки информации. На

Соматосенсорная кора
Получает информацию от специфических ядер таламуса. Зрительная и слуховая кора
Соматосенсорная кора
Получает информацию от специфических ядер таламуса. Зрительная и слуховая кора
 Исследовательская работа Проблемы заболеваний и повреждений листьев осенью
Исследовательская работа Проблемы заболеваний и повреждений листьев осенью Животный мир Пермского края
Животный мир Пермского края Постэмбриональный период
Постэмбриональный период Віроїди, мікоплазми, рикетсії, нематоди, квіткові рослини-паразити як збудники хвороб рослин
Віроїди, мікоплазми, рикетсії, нематоди, квіткові рослини-паразити як збудники хвороб рослин Внутренняя среда организма. Плазма крови
Внутренняя среда организма. Плазма крови Млекопитающие Башкортостана
Млекопитающие Башкортостана Невидимые нити в зимнем лесу
Невидимые нити в зимнем лесу Строение клетки. Цитология
Строение клетки. Цитология Жирафы. Интересные факты
Жирафы. Интересные факты Концепция эволюции. Биология и человек. (Часть 2)
Концепция эволюции. Биология и человек. (Часть 2) Механизмы эволюционного процесса. Формы адаптаций организмов
Механизмы эволюционного процесса. Формы адаптаций организмов Ассимиляция и диссимиляция. Метаболизм. Схема обмена веществ
Ассимиляция и диссимиляция. Метаболизм. Схема обмена веществ Імунні зрушення
Імунні зрушення Мотивация обучения
Мотивация обучения Презентация Среды жизни и места обитания животных7 кл.
Презентация Среды жизни и места обитания животных7 кл. Антропогенез. Гипотезы возникновения человека. Сходство и различия человека и животных. (Часть 1)
Антропогенез. Гипотезы возникновения человека. Сходство и различия человека и животных. (Часть 1) Структура нуклеиновых кислот. ДНК и РНК
Структура нуклеиновых кислот. ДНК и РНК Биология - наука о жизни
Биология - наука о жизни Опорно-рухова система
Опорно-рухова система Питание и пищеварение
Питание и пищеварение Влияние фитонцидов различных растений на жизнедеятельность колорадского жука
Влияние фитонцидов различных растений на жизнедеятельность колорадского жука Врожденные и приобретенные формы поведения животных и человека
Врожденные и приобретенные формы поведения животных и человека Действие фитонцидов комнатных растений на простые организмы
Действие фитонцидов комнатных растений на простые организмы Слуховой анализатор
Слуховой анализатор Оценка жизнеспособности деревьев и правила их отбора и назначения к вырубке и пересадке. (Лекция 13)
Оценка жизнеспособности деревьев и правила их отбора и назначения к вырубке и пересадке. (Лекция 13) Основні відомості про будову нуклеїнових кислот, їх властивості, процеси транскрипції та реплікації, трансляції
Основні відомості про будову нуклеїнових кислот, їх властивості, процеси транскрипції та реплікації, трансляції Строение клеток живых организмов. Лабораторная работа
Строение клеток живых организмов. Лабораторная работа Мозок людини
Мозок людини