- Физиология возбудимых клеток. Мышечное сокращение

Содержание
- 2. Этапы работы нервно-мышечного комплекса 1.Возбуждение мотонейрона 2.Электромеханическое сопряжение 3.Мышечное сокращение 4.Расслабление ПП=-90 мв, КУД=-50 мв, Длит
- 3. Строение скелетной мышцы Мышечное волокно: длина до 20см диаметр 0,1 мм Работа по принципу рычагов Волокнистые
- 4. Микроструктура мышечного волокна Актин- тонкие филаменты Миозин- толстые филаменты Т-система (поперечные Т-трубки + латеральные цистерны ЭПР)
- 5. Гипотеза скользящих нитей Гипотеза Хаксли 1955 Nebulin
- 6. Микрофотография мышечного волокна
- 7. Актин ТРОПОНИНОВЫЙ КОМПЛЕКС (через каждые 7 глобул актина): Тропонин С связывание с Са++ Тропонин Т -крепление
- 8. Прикрепление актина к внеклеточному матриксу F-Актин Ламинин Дистрофин
- 9. Миозин Головка миозина Нить актина хвост шейка головка В покое связан с АТФ, имеет сайт для
- 10. Механизм сокращения 1.Са++ выходит из цистерн ЭПР (опыт с экворином, Рюгель 1977) 2.Са++ связывается с тропонином
- 11. Кинематика сокращения
- 12. Расслабление мышцы Са++ удаляется от тропонина и уходит в ЭПР Тропомиозин блокирует актин Поперечные мостики актин-миозин
- 13. Электро–механическое сопряжение 1.ПД заходит в глубину Т-трубочек 2.Активация ДГП (дигидропиридиновых) Са+ каналов 3.Открытие рианодиновых Са++ каналов
- 14. Затраты АТФ мышцы На работу Na+-К+ насоса На работу Са++ насоса На разрыв мостиков актина и
- 15. Механика сокращения одиночного волокна 4. Зависимость ответа от частоты стимуляции 1. Соотношение между возбуждением и сокращением
- 16. Механика сокращения целой мышцы Б. Типы мышечных волокон А.Моторные (двигательные) единицы Размер моторной единицы (мыш. кл.)
- 17. Виды мышц
- 18. Гладкие мышцы Мелкие (2-10 мкм диаметр, 50-500 мкм длина) Одноядерные, способны к делению Механо- и хемочувствительны
- 20. Скачать презентацию
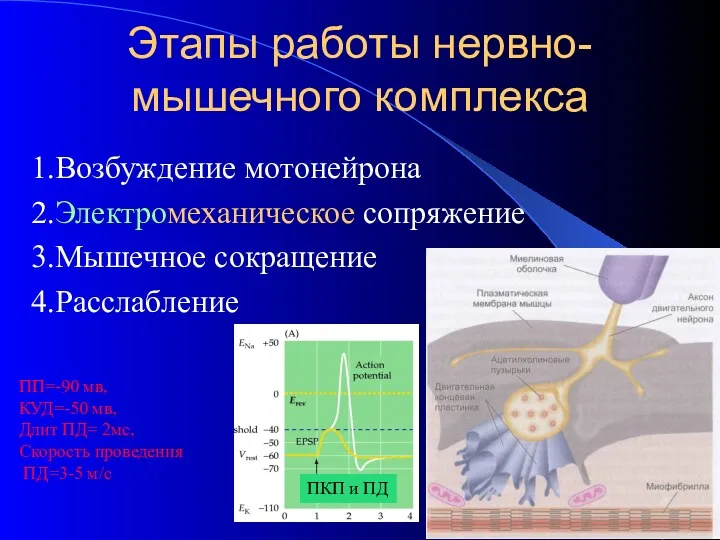
Этапы работы нервно-мышечного комплекса
1.Возбуждение мотонейрона
2.Электромеханическое сопряжение
3.Мышечное сокращение
4.Расслабление
ПП=-90 мв,
КУД=-50 мв,
Длит ПД=
Этапы работы нервно-мышечного комплекса
1.Возбуждение мотонейрона
2.Электромеханическое сопряжение
3.Мышечное сокращение
4.Расслабление
ПП=-90 мв,
КУД=-50 мв,
Длит ПД=
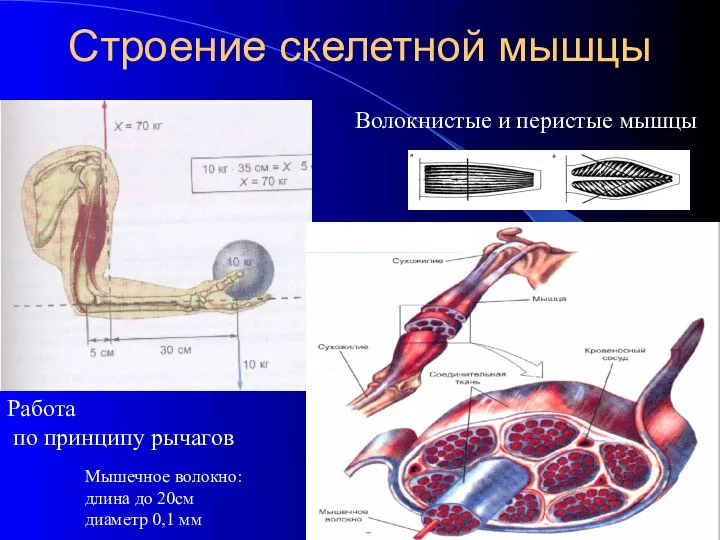
Строение скелетной мышцы
Мышечное волокно:
длина до 20см
диаметр 0,1 мм
Работа
по принципу рычагов
Волокнистые
Строение скелетной мышцы
Мышечное волокно:
длина до 20см
диаметр 0,1 мм
Работа
по принципу рычагов
Волокнистые
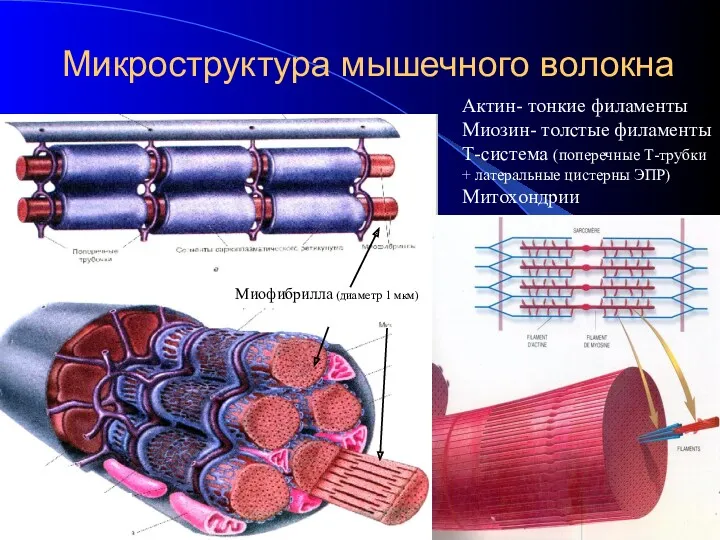
Микроструктура мышечного волокна
Актин- тонкие филаменты
Миозин- толстые филаменты
Т-система (поперечные Т-трубки + латеральные
Микроструктура мышечного волокна
Актин- тонкие филаменты
Миозин- толстые филаменты
Т-система (поперечные Т-трубки + латеральные
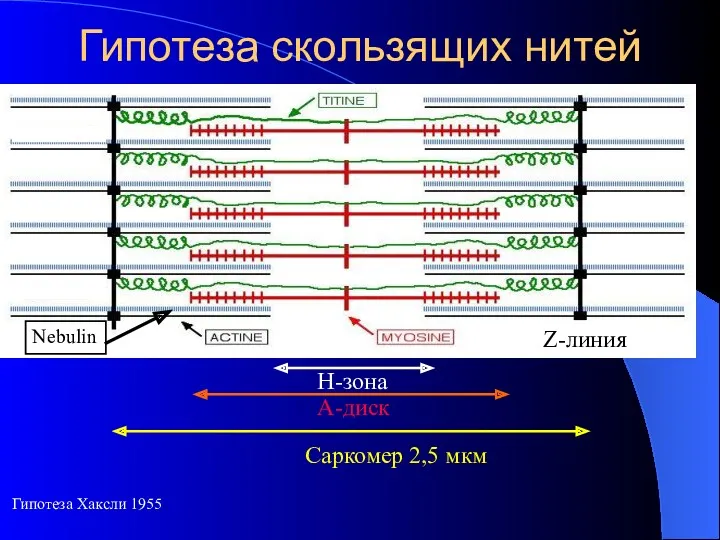
Гипотеза скользящих нитей
Гипотеза Хаксли 1955
Nebulin
Гипотеза скользящих нитей
Гипотеза Хаксли 1955
Nebulin

Микрофотография мышечного волокна
Микрофотография мышечного волокна
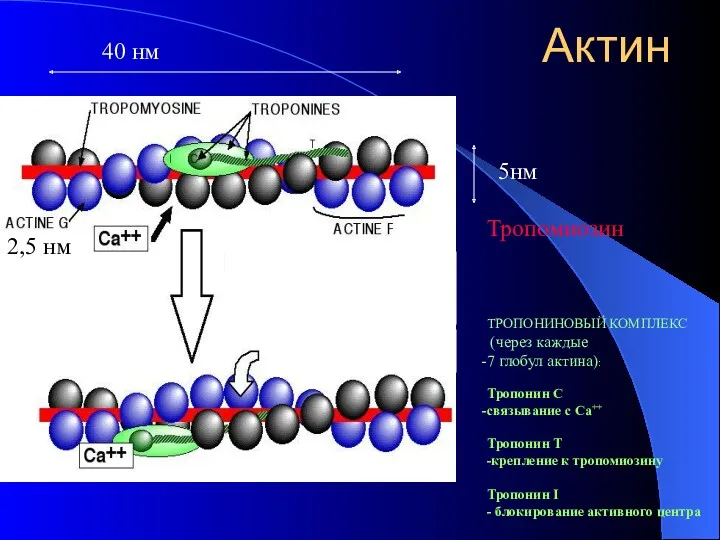
Актин
ТРОПОНИНОВЫЙ КОМПЛЕКС
(через каждые
7 глобул актина):
Тропонин С
связывание с Са++
Тропонин Т
-крепление
Актин
ТРОПОНИНОВЫЙ КОМПЛЕКС
(через каждые
7 глобул актина):
Тропонин С
связывание с Са++
Тропонин Т
-крепление
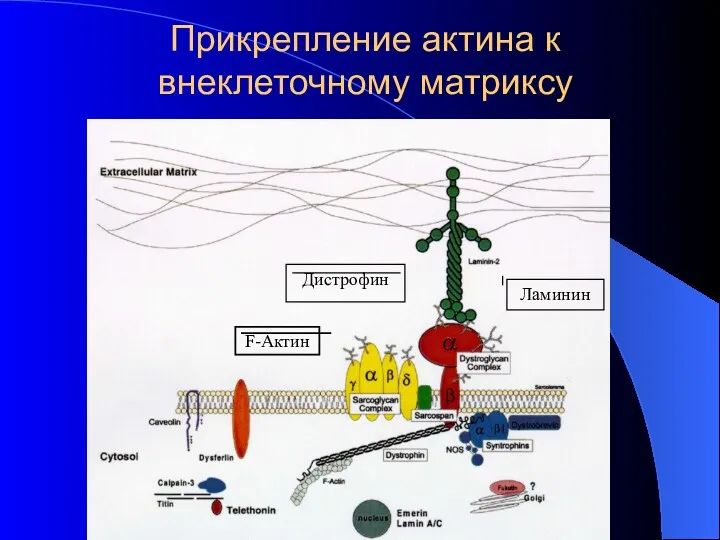
Прикрепление актина к внеклеточному матриксу
F-Актин
Ламинин
Дистрофин
Прикрепление актина к внеклеточному матриксу
F-Актин
Ламинин
Дистрофин
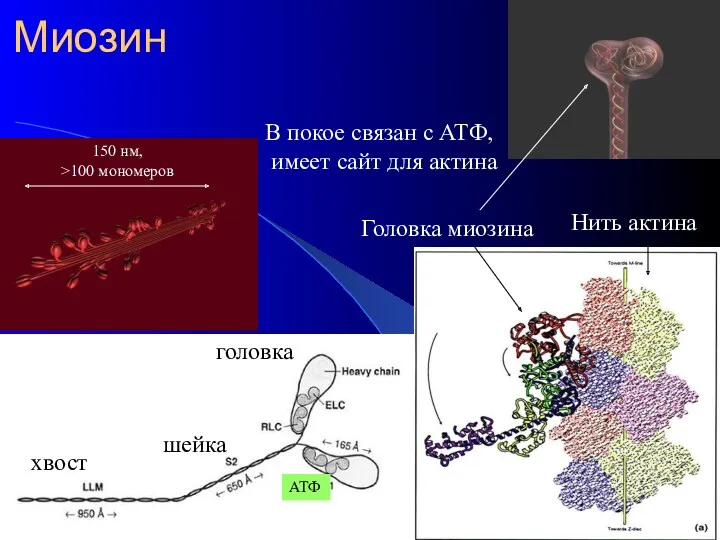
Миозин
Головка миозина
Нить актина
хвост
шейка
головка
В покое связан с АТФ,
имеет сайт для актина
АТФ
Миозин
Головка миозина
Нить актина
хвост
шейка
головка
В покое связан с АТФ,
имеет сайт для актина
АТФ
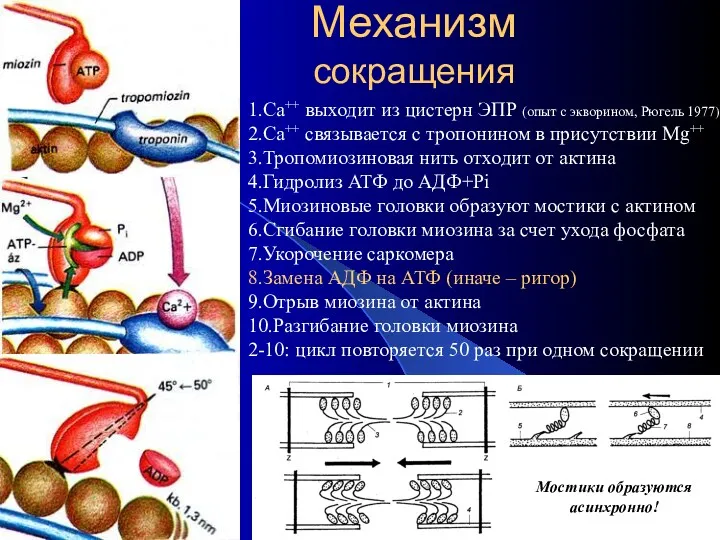
Механизм
сокращения
1.Са++ выходит из цистерн ЭПР (опыт с экворином, Рюгель 1977)
2.Са++
Механизм
сокращения
1.Са++ выходит из цистерн ЭПР (опыт с экворином, Рюгель 1977)
2.Са++
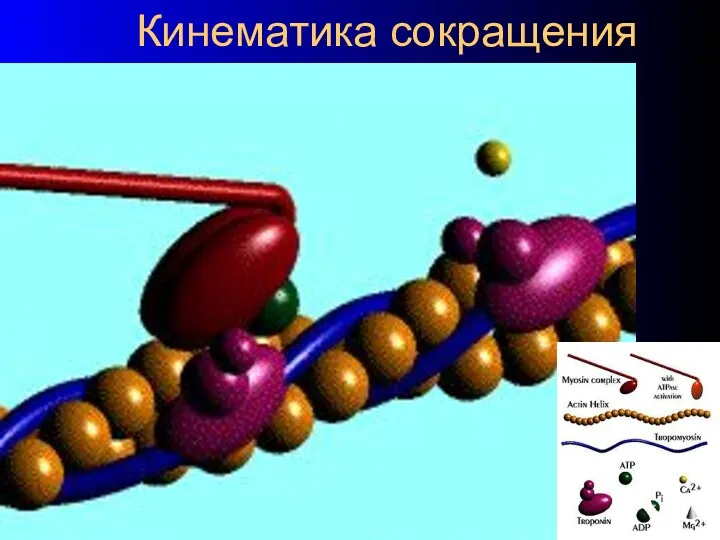
Кинематика сокращения
Кинематика сокращения
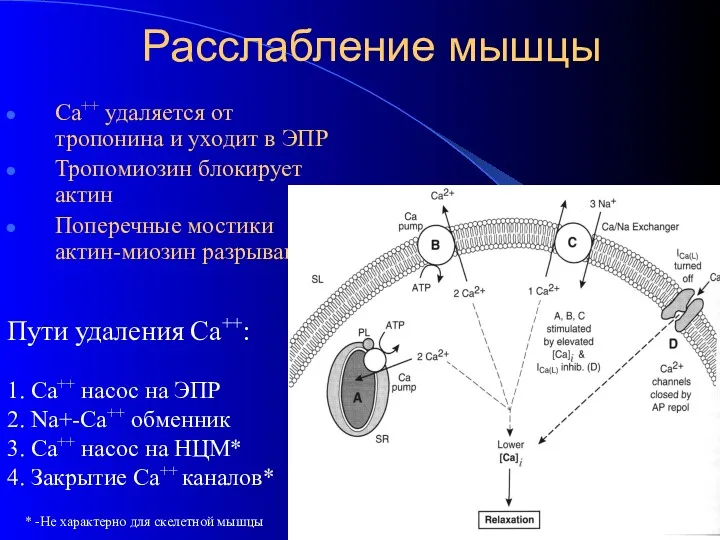
Расслабление мышцы
Са++ удаляется от тропонина и уходит в ЭПР
Тропомиозин блокирует актин
Поперечные
Расслабление мышцы
Са++ удаляется от тропонина и уходит в ЭПР
Тропомиозин блокирует актин
Поперечные
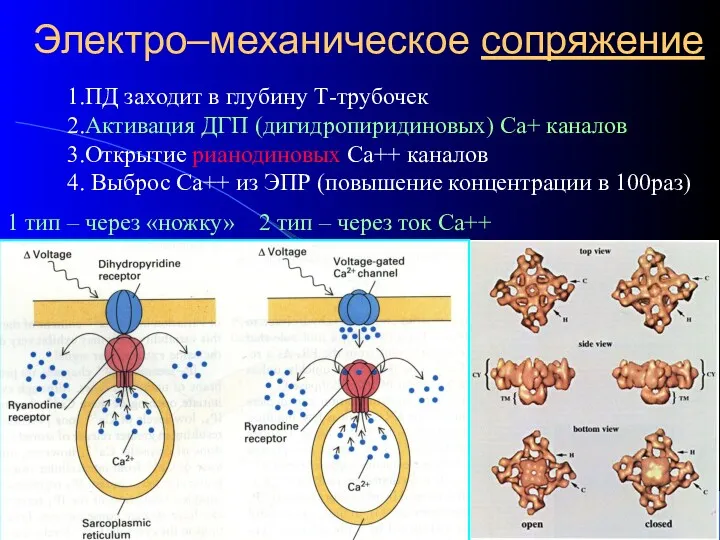
Электро–механическое сопряжение
1.ПД заходит в глубину Т-трубочек
2.Активация ДГП (дигидропиридиновых) Са+ каналов
3.Открытие рианодиновых
Электро–механическое сопряжение
1.ПД заходит в глубину Т-трубочек
2.Активация ДГП (дигидропиридиновых) Са+ каналов
3.Открытие рианодиновых

Затраты АТФ мышцы
На работу Na+-К+ насоса
На работу Са++ насоса
На разрыв мостиков
Затраты АТФ мышцы
На работу Na+-К+ насоса
На работу Са++ насоса
На разрыв мостиков
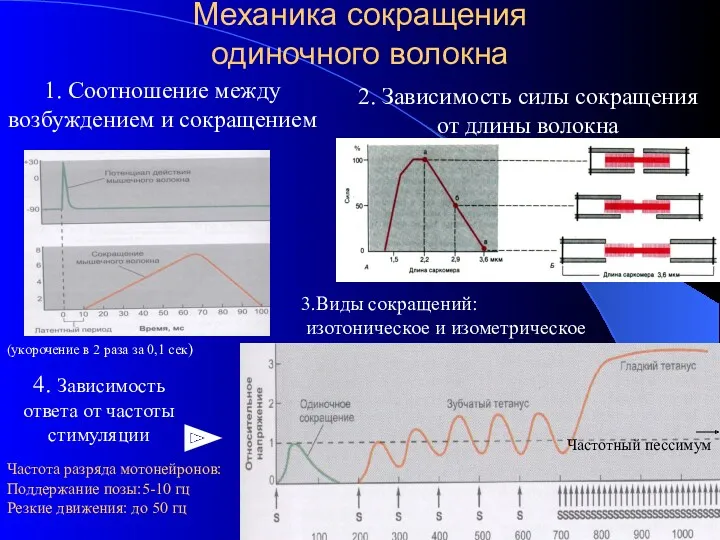
Механика сокращения
одиночного волокна
4. Зависимость ответа от частоты
стимуляции
1. Соотношение между
Механика сокращения
одиночного волокна
4. Зависимость ответа от частоты
стимуляции
1. Соотношение между

Механика сокращения
целой мышцы
Б. Типы мышечных волокон
А.Моторные (двигательные)
единицы
Размер моторной
единицы (мыш.
Механика сокращения
целой мышцы
Б. Типы мышечных волокон
А.Моторные (двигательные)
единицы
Размер моторной
единицы (мыш.
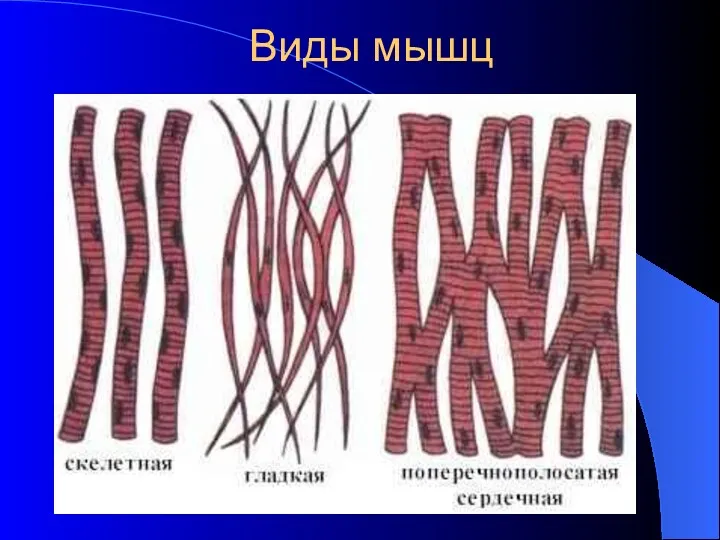
Виды мышц
Виды мышц

Гладкие мышцы
Мелкие (2-10 мкм диаметр, 50-500 мкм длина)
Одноядерные, способны к делению
Механо-
Гладкие мышцы
Мелкие (2-10 мкм диаметр, 50-500 мкм длина)
Одноядерные, способны к делению
Механо-
 Презентация к уроку на тему Видоизменение побегов в 6 классе
Презентация к уроку на тему Видоизменение побегов в 6 классе Презентация по биологии Гуморальная регуляция для 8 класса
Презентация по биологии Гуморальная регуляция для 8 класса Химический состав клетки
Химический состав клетки Статистические закономерности модификационной изменчивости
Статистические закономерности модификационной изменчивости Exotic animals
Exotic animals Виды корней и типы корневых систем
Виды корней и типы корневых систем Царство животные
Царство животные Презентация к уроку биологии Царство: грибы 6 класс Часть 1 Диск
Презентация к уроку биологии Царство: грибы 6 класс Часть 1 Диск Разнообразие растительного мира
Разнообразие растительного мира Редактирование генома
Редактирование генома Физиология дыхания. Газовый состав крови
Физиология дыхания. Газовый состав крови Органические вещества клетки. Углеводы. Часть 1. 10 класс
Органические вещества клетки. Углеводы. Часть 1. 10 класс Строение белков
Строение белков Безопасные ягоды и грибы
Безопасные ягоды и грибы Паразитические высшие растения
Паразитические высшие растения Екологічна ніша як наслідок адаптацій організмів певного виду до існування в екосистемі
Екологічна ніша як наслідок адаптацій організмів певного виду до існування в екосистемі Экологический урок: Культура обращения с отходами, раздельный сбор отходов в 2014 году
Экологический урок: Культура обращения с отходами, раздельный сбор отходов в 2014 году Кровь. Кроветворение (подготовка к контрольной работе)
Кровь. Кроветворение (подготовка к контрольной работе) Роль биологии в жизнедеятельности человека
Роль биологии в жизнедеятельности человека Всероссийский конкурс Мой лучший урок
Всероссийский конкурс Мой лучший урок Метаболизм
Метаболизм Принципы анатомического строения и функционирования анализаторов. Антиноцицептивная система. Стресс, его механизмы
Принципы анатомического строения и функционирования анализаторов. Антиноцицептивная система. Стресс, его механизмы Бесполое и половое размножение (10 класс)
Бесполое и половое размножение (10 класс) Лимфа жүйесінің құрылысы
Лимфа жүйесінің құрылысы Представители бактерий в Архангельской области
Представители бактерий в Архангельской области Прокариоты и эукариоты. Сравнение строения
Прокариоты и эукариоты. Сравнение строения Тип Кольчатые черви
Тип Кольчатые черви Многообразие грибов. Значение грибов в природе и жизни человека
Многообразие грибов. Значение грибов в природе и жизни человека