- Філогенія прокаріот

Содержание
- 2. * План лекції 1. Архебактерії найдавніші організми Землі 2. Огляд філем еубактерій 3. Гіпотези походження клітини
- 3. * Вууз, 1977 з співробітниками побудував філогенетичне дерево, яке базується на сиквенсі 16 S та 18
- 4. * Домініон ARCHEBACTERIA 1. Плазматична мембрана одношарова без жирних кислот. 2. Вона складається з простих етерів
- 5. *
- 6. * Царство CRENARCHEOTA Woese, 1990 Неметаногенні, сіркозалежні, термофільні (70-105°С) архебактерії, яфкі мають специфічну послідовність рРНК. Тип
- 7. * Царство ARCAETENERICUTOBIONTES Drozdov, 1997 Термофільні аеробні архебактерії, які не мають клітинних оболонок, містять гістоноподібні білки,
- 8. * Царство HALOBACTERIOBIONTES (Mohn, 1984) Галофільні (14-30% р-р хлориду натрія) термофільні (40-50°С) аеробні архебактерії, які містять
- 9. * Царство METANOBACTERIOBIONTES (Mohn, 1984) Облігатні анаероби, енергію отримують за рахунок окислення водню (він виділяється при
- 10. * Euryarchaeota Філогенія Архебактерій Crenarchaeota
- 11. * 2. ОГЛЯД ФІЛЕМ ЕУБАКТЕРІЙ Доминион EUBACTERIA Woese et Fox, 1977 1. Одноклітинні або багатоклітинні прокаріоти
- 12. * Надцарство GRACILICUTI (Gibbons, Murray, 1978) Мають специфічний тип будови клітинної оболонки. Царство CYANOBIONTES (Tachtadjan, 1974)
- 13. * Відділ СYANOPHYTA 2000 видів Основні ознаки відділу 1. Відсутність справжнього ядра. 2. Відсутність органел, що
- 14. * Філогенетичні зв’язки 1. Виникли в архейську еру (2,7-3,2 млрд. років тому). 2. Синьозелені водорості та
- 15. * Царство ANOXYPHOTOBACTERIOBIONTES Drozdov, 1977 Містять бактеріохлорофіли. Тип Rhodospirillophyles (Pfennig et Truper, 1971) Пурпурні бактерії Бактеріохлорофіли
- 16. * Царство SCOTOBACTERIOBIONTES (Gibbons, Murray, 1974) Хемоавтотрофи та хемоорганотрофи. Тип Thiobacillophyles Mohn, 1984 – Хемоавтотрофні бактерії,
- 17. * Тип AZOTOBACTERIOPHYLES (Becking, 1974) Mohn, 1984 Азотфіксуючі, нефотосинтезуючі (Azotobacter, Rhizobium). Тип ENTEROBACTERIOPHYLES (Rahn, 1937) Mohn,
- 18. * Тип CYTOPHAGOPHYLES (Stainer, 1940) Kusakin, Drozdov, 1997 Утворюють трихоми, можуть рухатися на твердих субстратах. На
- 19. * Царство SPIROCHAETOBACTERIOBIONTES Kusakin, Drozdov, 1997 Видовжені, тонкі, спірально закручені клітини, які мають у периплазматичному просторі
- 20. * Надцарство FIRMICUTOBIONTOI (Gibbons, Murray, 1978) Kusakin, Drozdov, 1997 Клітинні оболонки грампозитивні. Царство ACTINOBACTERIOBIONTES (Krasilnikov, 1949)
- 21. * Тип MYCOBACTERIOPHYLES (Chister, 1897) Kusakin, Drozdov, 1997 Здатні утворювати короткі нитковидні структури, без ендоспор та
- 22. * Царство EUFIRMICUTOBIONTES Kusakin, Drozdov, 1997 Справжні грампозитивні бактеріобіонти (кокі, бацили). Тип CLOSTRIDIOPHYLES Mohn, 1984 Бацили
- 23. * Царство TENERICUTOBIONTES (Murrey, 1984) Kusakin, Drozdov, 1997 Найдрібніші прокаріоти (0,2-0,3 мкм), які не здатні до
- 24. * Походження клітини доктор біологічних наук, професор О.Є. Ходосовцев Херсон - 2016
- 25. * План 1. Гіпотеза теплих водойм 2. Коацерватна гіпотеза 3. Хемоавтотрофна гіпотеза
- 26. 1. Гіпотеза теплих водойм Ч. Дарвін у 1871 р. писав в одному з листів: "Поширена думка,
- 27. * Життя виникло абіогенним шляхом. 2) Біологічній еволюції передувала довга хімічна еволюція. 3) Виникнення життя –
- 28. * Теорія коацерватів: Коацерватні системи – багатомолекулярні системи, які відокремилися від навколишнього середовища ліпідними мембранами 2)
- 29. *
- 30. * 7) Нуклеіновій кислоті належить вагома роль в організації живих систем, однак вона не є самодостатнім
- 31. * Міхаел Рассел (Michael Russel)– британський геофізик 3. Хемоавтотрофна гіпотеза
- 32. *
- 33. *
- 34. * Гіпотеза хемоавтотрофного походження клітини Рассела, Хейла 1997 життя виникло на великих глибинах (близько 4 км),
- 35. * a) відклади моносульфіда заліза, які утворилися в гідротермальних джерелах “черних курильників” 360 млн. років тому
- 36. * Модель походження життя в redox, pH та температур-ному градієнті, в підводних гідротер-мальних порожнинах.
- 37. * Етапи: Ера пребіотичної хімії Ера РНК Ера ДНК Ера ДНК та протеїнів 5) Ера універсального
- 38. * Модель утворення вільно-існуючих клітин 1) Ускладнення біохімічних процесів Незалежне виникнення біохімічних процесів, що привело до
- 39. * Архейне дерево життя Еоцитне дерево життя
- 40. * Гіпотезу молекулярного годинника запропонували 1962 р. Е. Цукеркандл та Л. Полінг Гіпотеза виходить з того,
- 42. Скачать презентацию

*
План лекції
1. Архебактерії найдавніші організми Землі
2. Огляд філем еубактерій
3. Гіпотези походження
*
План лекції
1. Архебактерії найдавніші організми Землі
2. Огляд філем еубактерій
3. Гіпотези походження
*
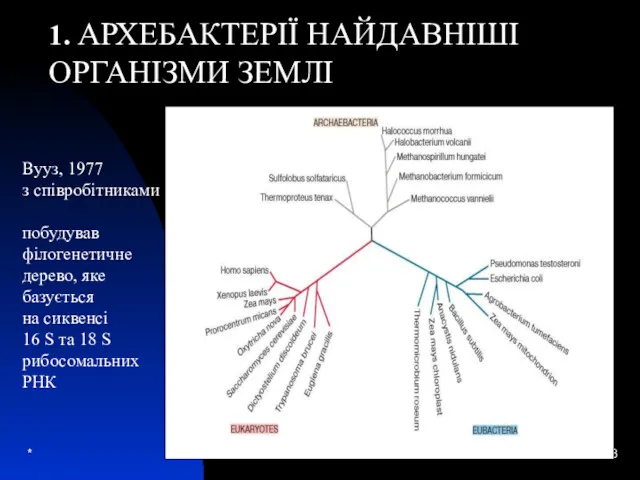
Вууз, 1977
з співробітниками
побудував
філогенетичне
дерево, яке
базується
на сиквенсі
16 S та 18 S
рибосомальних
РНК
1.
*
Вууз, 1977
з співробітниками
побудував
філогенетичне
дерево, яке
базується
на сиквенсі
16 S та 18 S
рибосомальних
РНК
1.

*
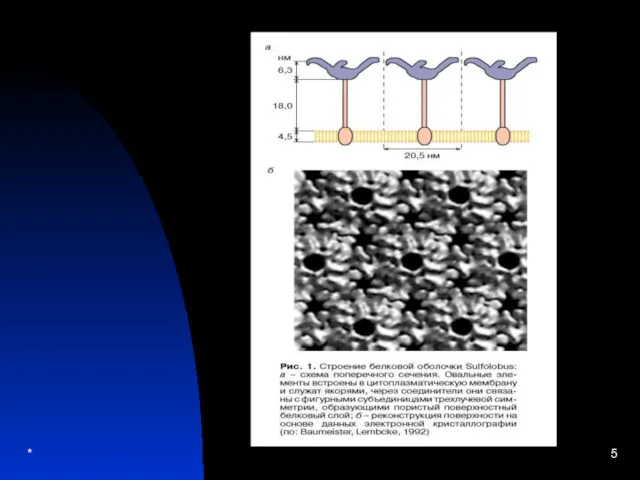
Домініон ARCHEBACTERIA
1. Плазматична мембрана одношарова без жирних кислот.
2. Вона складається з
*
Домініон ARCHEBACTERIA
1. Плазматична мембрана одношарова без жирних кислот.
2. Вона складається з
*
*

*
Царство CRENARCHEOTA Woese, 1990
Неметаногенні, сіркозалежні, термофільні (70-105°С) архебактерії, яфкі мають
*
Царство CRENARCHEOTA Woese, 1990
Неметаногенні, сіркозалежні, термофільні (70-105°С) архебактерії, яфкі мають

*
Царство ARCAETENERICUTOBIONTES Drozdov, 1997
Термофільні аеробні архебактерії, які не мають клітинних оболонок,
*
Царство ARCAETENERICUTOBIONTES Drozdov, 1997
Термофільні аеробні архебактерії, які не мають клітинних оболонок,

*
Царство HALOBACTERIOBIONTES
(Mohn, 1984)
Галофільні (14-30% р-р хлориду натрія) термофільні (40-50°С) аеробні
*
Царство HALOBACTERIOBIONTES
(Mohn, 1984)
Галофільні (14-30% р-р хлориду натрія) термофільні (40-50°С) аеробні

*
Царство METANOBACTERIOBIONTES (Mohn, 1984)
Облігатні анаероби, енергію отримують за рахунок окислення
*
Царство METANOBACTERIOBIONTES (Mohn, 1984)
Облігатні анаероби, енергію отримують за рахунок окислення
*
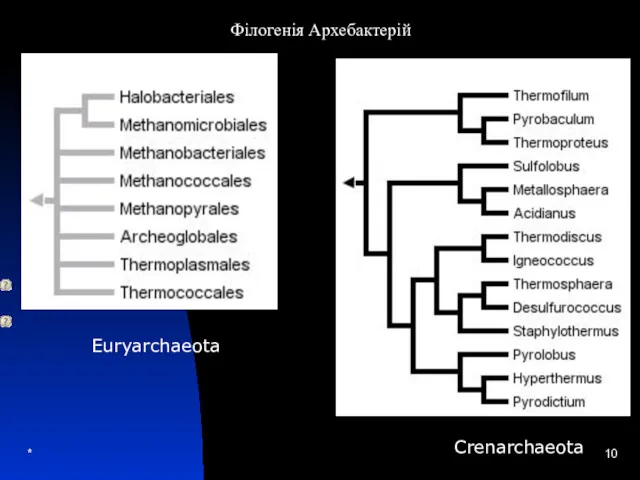
Euryarchaeota
Філогенія Архебактерій
Crenarchaeota
*
Euryarchaeota
Філогенія Архебактерій
Crenarchaeota

*
2. ОГЛЯД ФІЛЕМ ЕУБАКТЕРІЙ
Доминион EUBACTERIA Woese et Fox, 1977
1. Одноклітинні або
*
2. ОГЛЯД ФІЛЕМ ЕУБАКТЕРІЙ
Доминион EUBACTERIA Woese et Fox, 1977
1. Одноклітинні або

*
Надцарство GRACILICUTI
(Gibbons, Murray, 1978)
Мають специфічний тип будови клітинної оболонки.
Царство
*
Надцарство GRACILICUTI
(Gibbons, Murray, 1978)
Мають специфічний тип будови клітинної оболонки.
Царство

*
Відділ СYANOPHYTA
2000 видів
Основні ознаки відділу
1. Відсутність справжнього ядра.
2. Відсутність органел, що
*
Відділ СYANOPHYTA
2000 видів
Основні ознаки відділу
1. Відсутність справжнього ядра.
2. Відсутність органел, що

*
Філогенетичні зв’язки
1. Виникли в архейську еру (2,7-3,2 млрд. років тому).
2. Синьозелені
*
Філогенетичні зв’язки
1. Виникли в архейську еру (2,7-3,2 млрд. років тому).
2. Синьозелені

*
Царство ANOXYPHOTOBACTERIOBIONTES Drozdov, 1977
Містять бактеріохлорофіли.
Тип Rhodospirillophyles (Pfennig et Truper, 1971) Пурпурні
*
Царство ANOXYPHOTOBACTERIOBIONTES Drozdov, 1977
Містять бактеріохлорофіли.
Тип Rhodospirillophyles (Pfennig et Truper, 1971) Пурпурні

*
Царство
SCOTOBACTERIOBIONTES (Gibbons, Murray, 1974)
Хемоавтотрофи та хемоорганотрофи.
Тип Thiobacillophyles Mohn, 1984 – Хемоавтотрофні
*
Царство
SCOTOBACTERIOBIONTES (Gibbons, Murray, 1974)
Хемоавтотрофи та хемоорганотрофи.
Тип Thiobacillophyles Mohn, 1984 – Хемоавтотрофні

*
Тип AZOTOBACTERIOPHYLES (Becking, 1974) Mohn, 1984
Азотфіксуючі, нефотосинтезуючі (Azotobacter, Rhizobium).
Тип ENTEROBACTERIOPHYLES (Rahn,
*
Тип AZOTOBACTERIOPHYLES (Becking, 1974) Mohn, 1984
Азотфіксуючі, нефотосинтезуючі (Azotobacter, Rhizobium).
Тип ENTEROBACTERIOPHYLES (Rahn,

*
Тип CYTOPHAGOPHYLES (Stainer, 1940)
Kusakin, Drozdov, 1997
Утворюють трихоми, можуть рухатися
*
Тип CYTOPHAGOPHYLES (Stainer, 1940)
Kusakin, Drozdov, 1997
Утворюють трихоми, можуть рухатися

*
Царство SPIROCHAETOBACTERIOBIONTES Kusakin, Drozdov, 1997
Видовжені, тонкі, спірально закручені клітини, які мають
*
Царство SPIROCHAETOBACTERIOBIONTES Kusakin, Drozdov, 1997
Видовжені, тонкі, спірально закручені клітини, які мають

*
Надцарство FIRMICUTOBIONTOI
(Gibbons, Murray, 1978) Kusakin, Drozdov, 1997
Клітинні оболонки грампозитивні.
Царство ACTINOBACTERIOBIONTES (Krasilnikov,
*
Надцарство FIRMICUTOBIONTOI
(Gibbons, Murray, 1978) Kusakin, Drozdov, 1997
Клітинні оболонки грампозитивні.
Царство ACTINOBACTERIOBIONTES (Krasilnikov,

*
Тип MYCOBACTERIOPHYLES (Chister, 1897)
Kusakin, Drozdov, 1997
Здатні утворювати короткі нитковидні структури,
*
Тип MYCOBACTERIOPHYLES (Chister, 1897)
Kusakin, Drozdov, 1997
Здатні утворювати короткі нитковидні структури,

*
Царство EUFIRMICUTOBIONTES Kusakin, Drozdov, 1997
Справжні грампозитивні бактеріобіонти (кокі, бацили).
Тип CLOSTRIDIOPHYLES
*
Царство EUFIRMICUTOBIONTES Kusakin, Drozdov, 1997
Справжні грампозитивні бактеріобіонти (кокі, бацили).
Тип CLOSTRIDIOPHYLES

*
Царство TENERICUTOBIONTES
(Murrey, 1984) Kusakin, Drozdov, 1997
Найдрібніші прокаріоти (0,2-0,3 мкм), які не
*
Царство TENERICUTOBIONTES
(Murrey, 1984) Kusakin, Drozdov, 1997
Найдрібніші прокаріоти (0,2-0,3 мкм), які не

*
Походження клітини
доктор біологічних наук, професор
О.Є. Ходосовцев
Херсон - 2016
*
Походження клітини
доктор біологічних наук, професор
О.Є. Ходосовцев
Херсон - 2016

*
План
1. Гіпотеза теплих водойм
2. Коацерватна гіпотеза
3. Хемоавтотрофна гіпотеза
*
План
1. Гіпотеза теплих водойм
2. Коацерватна гіпотеза
3. Хемоавтотрофна гіпотеза

1. Гіпотеза теплих водойм
Ч. Дарвін у 1871 р. писав в одному
1. Гіпотеза теплих водойм
Ч. Дарвін у 1871 р. писав в одному

*
Життя виникло абіогенним шляхом.
2) Біологічній еволюції передувала довга
хімічна еволюція.
3) Виникнення
*
Життя виникло абіогенним шляхом.
2) Біологічній еволюції передувала довга
хімічна еволюція.
3) Виникнення

*
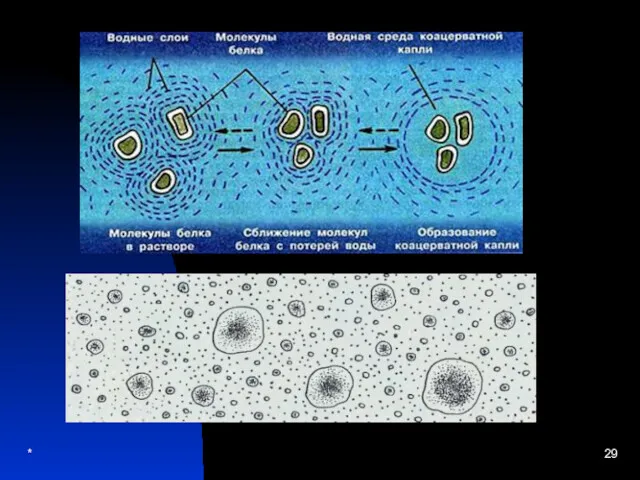
Теорія коацерватів:
Коацерватні системи – багатомолекулярні системи, які
відокремилися від навколишнього середовища
*
Теорія коацерватів:
Коацерватні системи – багатомолекулярні системи, які
відокремилися від навколишнього середовища
*
*

*
7) Нуклеіновій кислоті належить вагома роль в організації живих систем,
однак вона
*
7) Нуклеіновій кислоті належить вагома роль в організації живих систем,
однак вона

*
Міхаел Рассел
(Michael Russel)–
британський геофізик
3. Хемоавтотрофна гіпотеза
*
Міхаел Рассел
(Michael Russel)–
британський геофізик
3. Хемоавтотрофна гіпотеза

*
*

*
*

*
Гіпотеза хемоавтотрофного походження клітини
Рассела, Хейла 1997
життя виникло на великих
*
Гіпотеза хемоавтотрофного походження клітини
Рассела, Хейла 1997
життя виникло на великих
*
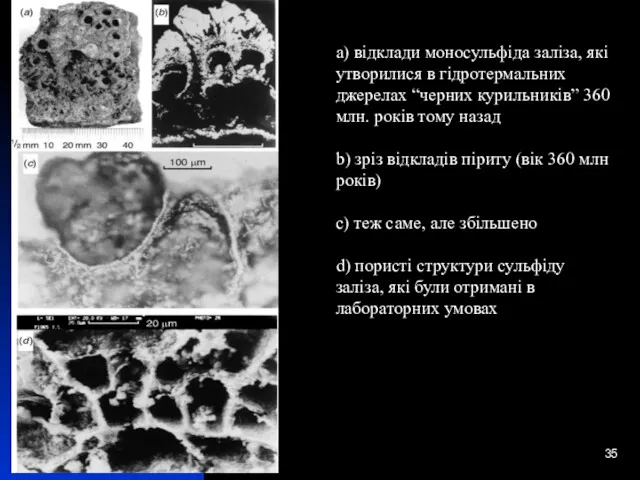
a) відклади моносульфіда заліза, які утворилися в гідротермальних джерелах “черних курильників”
*
a) відклади моносульфіда заліза, які утворилися в гідротермальних джерелах “черних курильників”
*
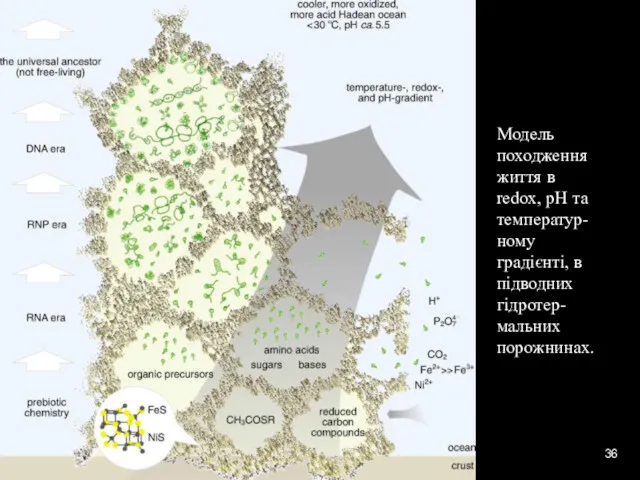
Модель походження життя в redox, pH та температур-ному градієнті, в підводних
*
Модель походження життя в redox, pH та температур-ному градієнті, в підводних

*
Етапи:
Ера пребіотичної хімії
Ера РНК
Ера ДНК
Ера ДНК та протеїнів
5) Ера універсального не
*
Етапи:
Ера пребіотичної хімії
Ера РНК
Ера ДНК
Ера ДНК та протеїнів
5) Ера універсального не
*
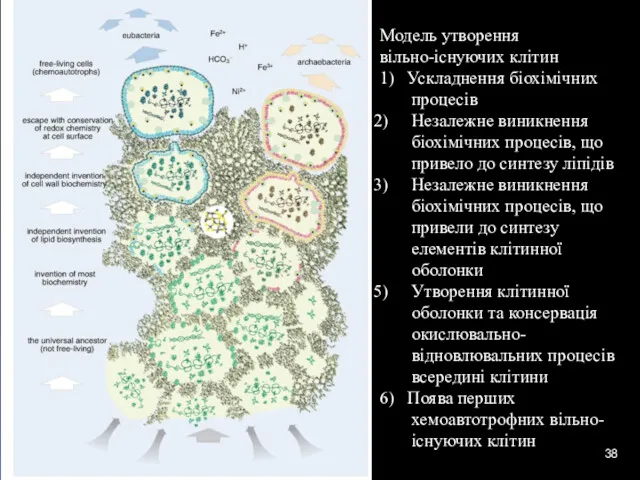
Модель утворення
вільно-існуючих клітин
1) Ускладнення біохімічних процесів
Незалежне виникнення біохімічних процесів, що
*
Модель утворення
вільно-існуючих клітин
1) Ускладнення біохімічних процесів
Незалежне виникнення біохімічних процесів, що
*
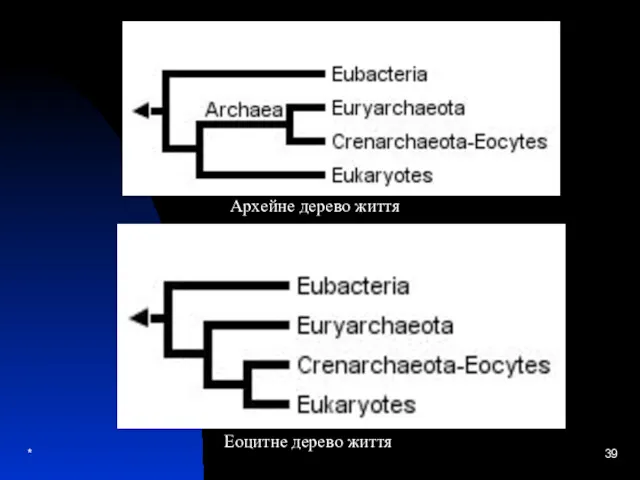
Архейне дерево життя
Еоцитне дерево життя
*
Архейне дерево життя
Еоцитне дерево життя

*
Гіпотезу молекулярного годинника запропонували
1962 р. Е. Цукеркандл та Л. Полінг
Гіпотеза виходить
*
Гіпотезу молекулярного годинника запропонували
1962 р. Е. Цукеркандл та Л. Полінг
Гіпотеза виходить
 Презентация по биологии
Презентация по биологии Общая характеристика отдела Цветковые (Покрытосеменные) растения
Общая характеристика отдела Цветковые (Покрытосеменные) растения Онтогенез. Строение половых клеток
Онтогенез. Строение половых клеток Презентация к уроку Наука биология
Презентация к уроку Наука биология Жизненные формы высших растений
Жизненные формы высших растений Ріст рослин в умовах невагомості
Ріст рослин в умовах невагомості Приспособленность организмов и её относительность
Приспособленность организмов и её относительность Сүт. Сүттің құрамы
Сүт. Сүттің құрамы Науково-технічні засоби в експертній практиці: актуальні питання та концептуальні засади
Науково-технічні засоби в експертній практиці: актуальні питання та концептуальні засади Ученые- биологи
Ученые- биологи Биологические исследования
Биологические исследования Почки растений
Почки растений Морские свинки
Морские свинки Теории происхождения человека
Теории происхождения человека Грибы, бактерии, царства животные, растения
Грибы, бактерии, царства животные, растения Бактериальный фотосинтез
Бактериальный фотосинтез Про белку. Среда обитания:
Про белку. Среда обитания: Самоочищение водоема
Самоочищение водоема Генетика пола и наследование, сцепленное с полом
Генетика пола и наследование, сцепленное с полом Уровни организации жизни на земле
Уровни организации жизни на земле Урок по биологии по теме Простейшие. Голые амёбы.
Урок по биологии по теме Простейшие. Голые амёбы. Использование материалов аэро - и космических съемок для целей сельского хозяйства. Исследования почвенного покрова земли
Использование материалов аэро - и космических съемок для целей сельского хозяйства. Исследования почвенного покрова земли Клеточная мембрана
Клеточная мембрана Высшая нервная деятельность
Высшая нервная деятельность Тип хордовые. Общая характеристика. Подтип бесчерепные. Ланцетник
Тип хордовые. Общая характеристика. Подтип бесчерепные. Ланцетник Теория внешнего вмешательства
Теория внешнего вмешательства Закономерности развития живой природы
Закономерности развития живой природы Растения культурные и дикорастущие
Растения культурные и дикорастущие