- Функциональная морфология биологических мембран

Содержание
- 2. Биологическая мембрана ультратонкая пленка, состоящая из двойного слоя липидных молекул, с которым связаны белки и полисахариды.
- 3. Строение биологической мембраны Молекулы мембранных липидов – амфифильные молекулы, т.е. имеют полярную («головку») и неполярную («хвост»)
- 4. Структура мембран
- 5. Сборка липидного бислоя Находящиеся в водной фазе фосфолипиды формируют двухслойные структуры, объединяя свои гидрофобные участки. Когда
- 6. Плазматическая мембрана (внешняя клеточная мембрана, плазмолемма) основной, постоянный, универсальный для всех клеток компонент системы поверхностного аппарата.
- 7. Плазматическая мембрана Структура толщиной 7-10 нм, образованная, главным образом, липидами и белками. Молекулярное строение плазмолеммы описывается
- 8. Жирные кислоты и свойства мембраны Чем длиннее алифатический радикал, тем толще липидный бислой (орt-16-20). 2. Чем
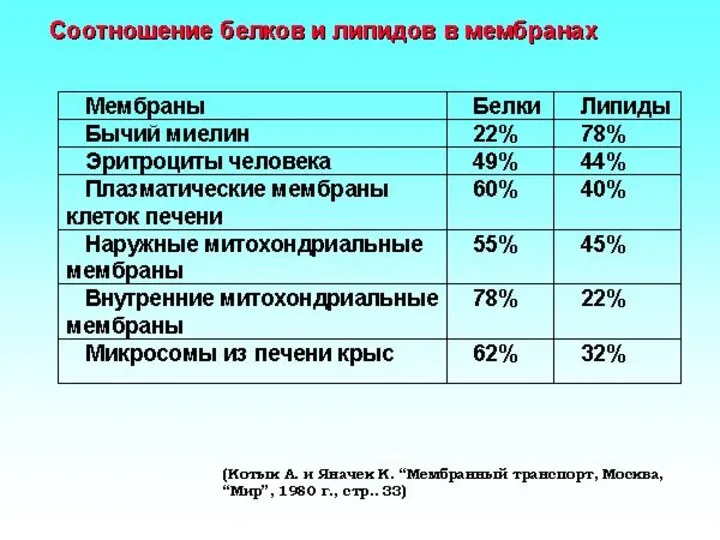
- 9. Содержание фосфолипидов в плазматической мембране
- 10. Холестерин и проницаемость биологической мембраны В мембране с преобладанием насыщенных жирных кислот («жесткая» мембрана), молекула холестерола
- 11. Мембранные белки - функционально гетерогенная группа белков (рецепторы, ферменты, переносчики, каналы и др.), обеспечивающая специфические свойства
- 13. Химическая модификация мембранных липидов и белков Молекулы олигосахаридов связываются с липидами (гликолипиды) и мембранными белками (гликопротеины).
- 14. Гликокаликс С наружной стороны плазмолеммы имеется надмембранный слой - гликокаликс (3-4 нм). Он образован углеводными фрагментами
- 15. Субмембранный комплекс Функции: Участвует в поддержании формы клетки. Участвует в формировании межклеточных контактов. Обеспечивает мембранные процессы.
- 16. Мобильность липидов При температуре тела мембрана текуча и липиды свободно перемещаются в ней. Латеральная подвижность, то
- 17. Перемещение мембранных белков Белковые молекулы мозаично распределены в липидном бислое и перемещаться в его плоскости. Перемещение
- 18. Липидные рафты домены липидного бислоя клеточной мембраны, обогащённые холестерином, и насыщенными фосфолипидами. Это участок плотно упакованного
- 19. Обновление мембран в клетке Баланс эндоцитоз/экзоцитоз для плазмолеммы. Обмен везикулами между органеллами. Синтез de novo компонентов
- 20. Функции плазмолеммы: барьерная; транспорт веществ в клетку и из клетки; взаимодействие с сигнальными молекулами; взаимодействие с
- 21. Рецепторы гликопротеины, способные высокоселективно связываться с определенными молекулами - лигандами (гормон, медиатор). Регулируют проницаемость плазмолеммы. Передают
- 22. Каталитические рецепторы - цитоплазматический домен обладает ферментативной активностью. При связывании лиганда внеклеточным доменом активируется протеинкиназная или
- 23. Рецепторы-каналы После связывания с лигандом, изменяют свою конформацию, что ведет к возникновению в мембране гидрофильного канала
- 24. Рецепторы, связанные с G-белками
- 25. Трансмембранный транспорт веществ прямой (диффузия, осмос, фильтрация); опосредованный: 1. пассивный (с участием белка-переносчика или с участием
- 26. Простая диффузия - переход вещества из области более высокой концентрации в область меньшей. Небольшие нейтральные молекулы
- 27. Осмос движение молекул растворителя через полупроницаемую мембрану из области с низкой концентрацией растворенного вещества в область
- 28. Фильтрация движение веществ через поры под действием избыточного давления.
- 29. Облегчённая диффузия транспорт веществ через мембрану с помощью специальных белков. Поры – отверстия в мембране, образованные
- 30. Эндоцитоз вид активного транспорта, с формированием эндоцитозной везикулы. Всегда рецептор-опосредованный Всегда с участием цитоскелета
- 31. Виды эндоцитоза Фагоцитоз - процесс поглощения клеткой объектов размером >10-6 м, таких как бактерии, вирусы, остатки
- 32. Экзоцитоз - процесс слияния экзоцитозного пузырька с плазматической мембраной клетки в результате которого его содержимое освобождается
- 33. Первично-активный транспорт перенос веществ через мембрану против градиента концентрации при участии транспортных АТФ-аз. Na+/K+-АТФ-аза, Са++-АТФ-аза, Н+/К+-АТФ-аза.
- 34. Вторично-активный транспорт перенос веществ против градиента концентрации сопряжен с одновременным переносом другого вещества по градиенту концентрации
- 35. Межклеточные контакты Механические – контакты, которые обеспечивают механическую связь клеток друг с другом (интердигитации, десмосомы, промежуточные
- 36. Интердигитации Наиболее просто устроенный вид взаимодействия мембран смежных клеток. Представлены выпячиваниями цитоплазмы через плазмолемму одной клетки,
- 37. Плотное соединение соединение наружных листков плазмолемм взаимодействующих клеток, опосредованное трансмембранными белками-окклюдинами. Окклюдины мембраны одной клетки взаимодействуют
- 38. Промежуточное соединение, опоясывающая десмосома Охватывают клетку по периметру в виде пояса. С цитоплазматической стороны плазмолеммы в
- 39. Десмосома (пятно сцепления) Межклеточная щель расширена (25 нм).
- 40. Нексус (щелевое соединение) Образован соединением трансмембранных гексамеров – коннексонами (10 нм) одной клетки, с а налогичными
- 41. Интегрины суперсемейство молекул адгезии к молекулам межклеточного матрикса: Коллаген. Фибронектин. Ламинин. Все интегрины гетеродимерные трансмембранные белки,
- 43. Скачать презентацию
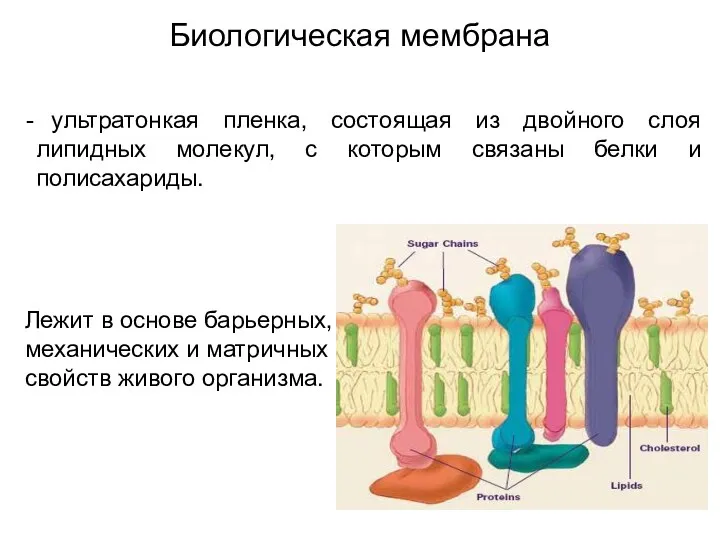
Биологическая мембрана
ультратонкая пленка, состоящая из двойного слоя липидных молекул, с
Биологическая мембрана
ультратонкая пленка, состоящая из двойного слоя липидных молекул, с
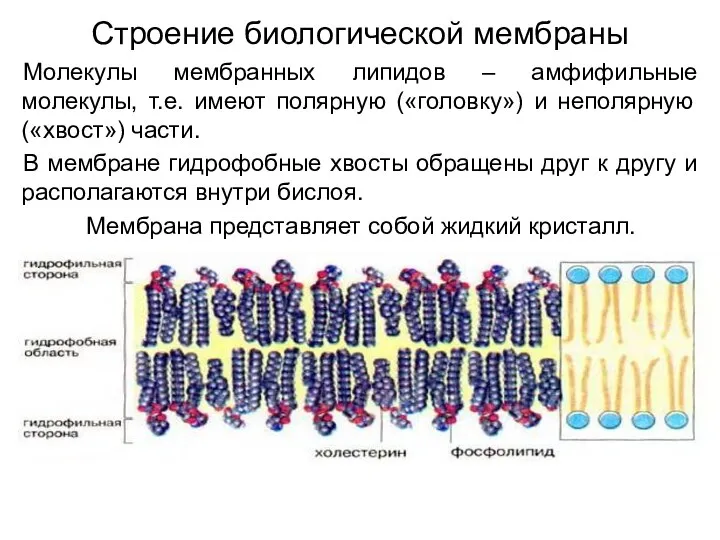
Строение биологической мембраны
Молекулы мембранных липидов – амфифильные молекулы, т.е. имеют полярную
Строение биологической мембраны
Молекулы мембранных липидов – амфифильные молекулы, т.е. имеют полярную
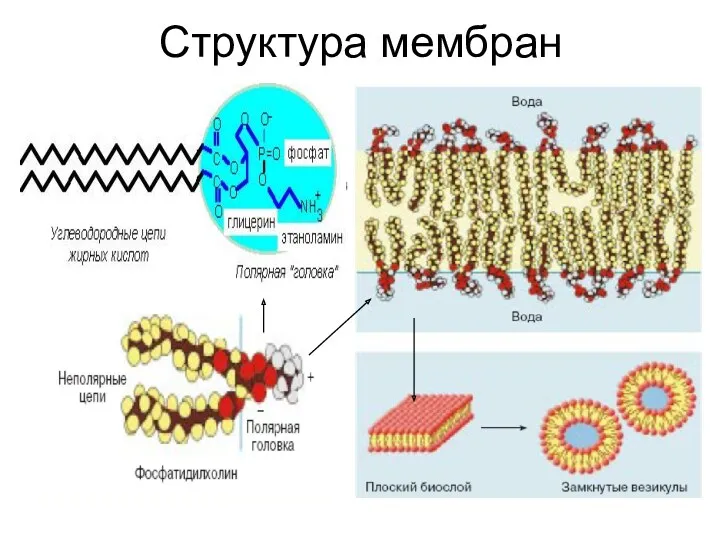
Структура мембран
Структура мембран

Сборка липидного бислоя
Находящиеся в водной фазе фосфолипиды формируют двухслойные структуры, объединяя
Сборка липидного бислоя
Находящиеся в водной фазе фосфолипиды формируют двухслойные структуры, объединяя
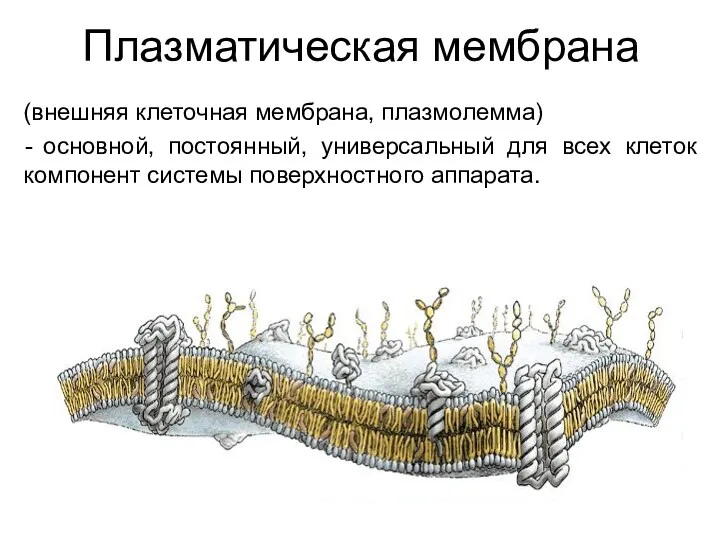
Плазматическая мембрана
(внешняя клеточная мембрана, плазмолемма)
основной, постоянный, универсальный для всех клеток
Плазматическая мембрана
(внешняя клеточная мембрана, плазмолемма)
основной, постоянный, универсальный для всех клеток
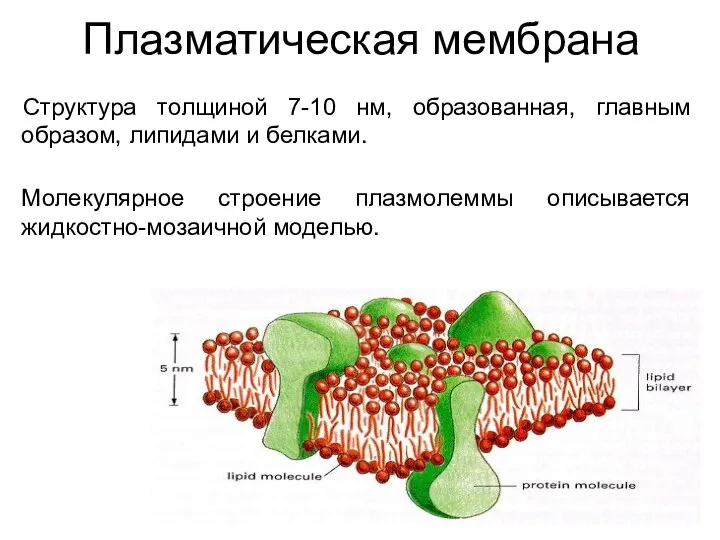
Плазматическая мембрана
Структура толщиной 7-10 нм, образованная, главным образом, липидами и белками.
Молекулярное
Плазматическая мембрана
Структура толщиной 7-10 нм, образованная, главным образом, липидами и белками.
Молекулярное
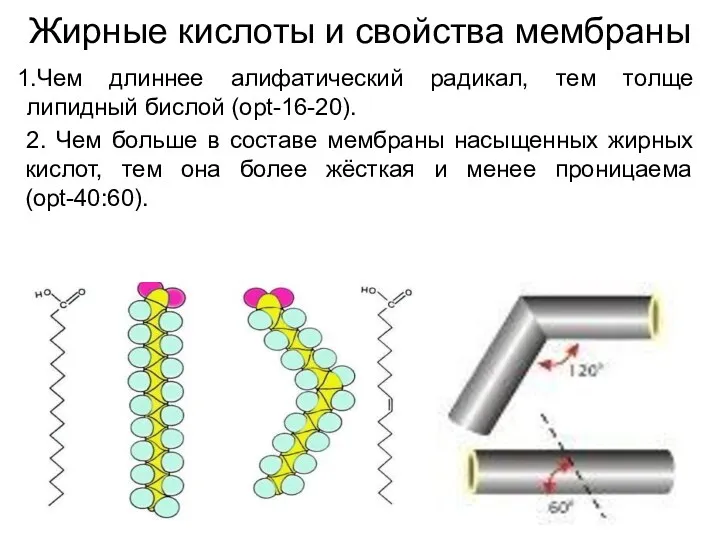
Жирные кислоты и свойства мембраны
Чем длиннее алифатический радикал, тем толще липидный
Жирные кислоты и свойства мембраны
Чем длиннее алифатический радикал, тем толще липидный
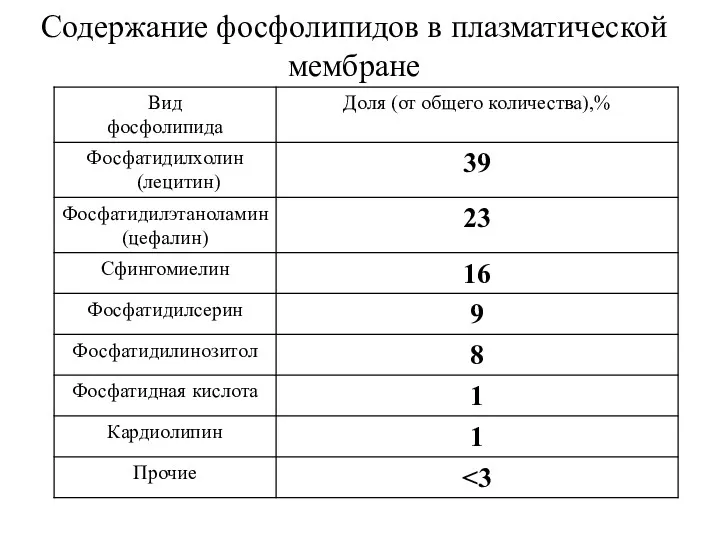
Содержание фосфолипидов в плазматической мембране
Содержание фосфолипидов в плазматической мембране
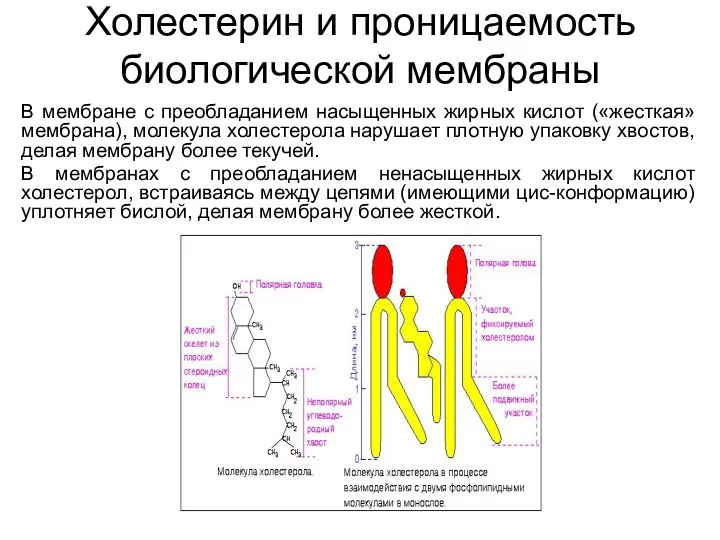
Холестерин и проницаемость биологической мембраны
В мембране с преобладанием насыщенных жирных кислот
Холестерин и проницаемость биологической мембраны
В мембране с преобладанием насыщенных жирных кислот

Мембранные белки
- функционально гетерогенная группа белков (рецепторы, ферменты, переносчики, каналы и
Мембранные белки
- функционально гетерогенная группа белков (рецепторы, ферменты, переносчики, каналы и
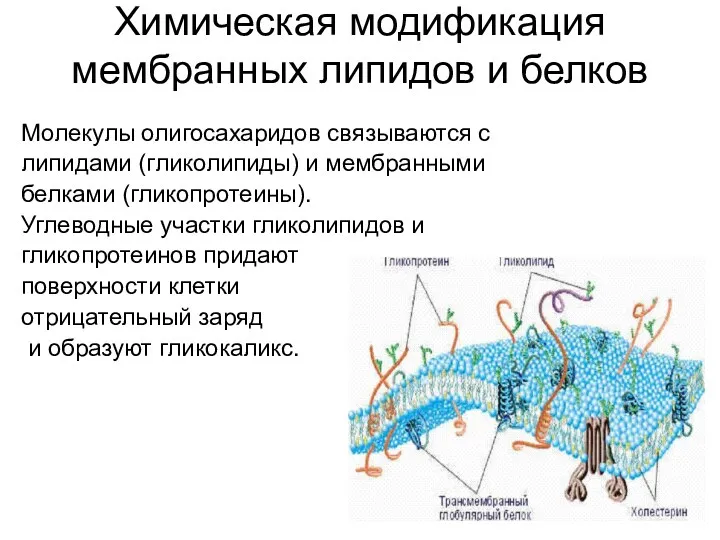
Химическая модификация мембранных липидов и белков
Молекулы олигосахаридов связываются с
липидами (гликолипиды)
Химическая модификация мембранных липидов и белков
Молекулы олигосахаридов связываются с
липидами (гликолипиды)
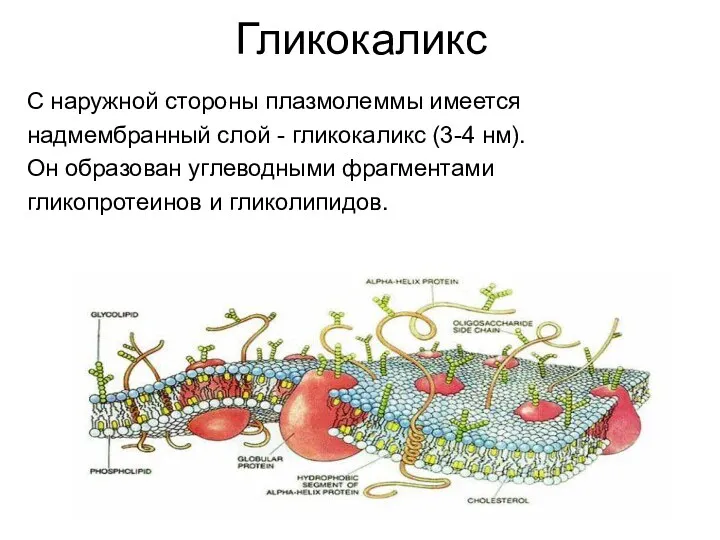
Гликокаликс
С наружной стороны плазмолеммы имеется
надмембранный слой - гликокаликс (3-4 нм).
Он
Гликокаликс
С наружной стороны плазмолеммы имеется
надмембранный слой - гликокаликс (3-4 нм).
Он
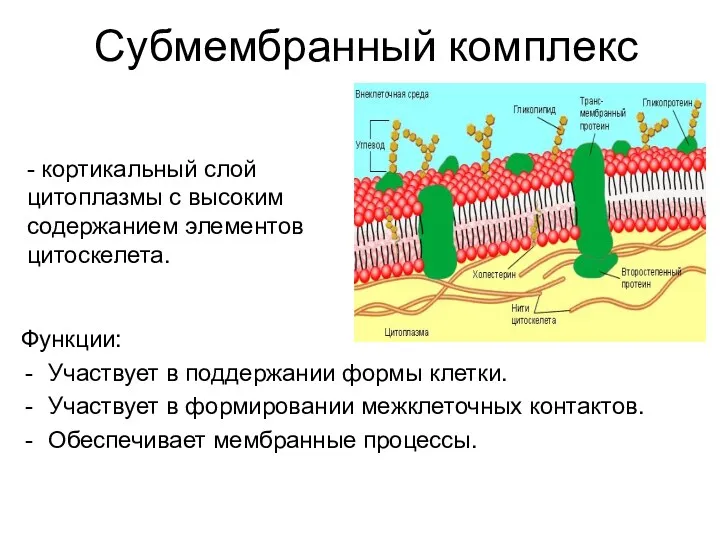
Субмембранный комплекс
Функции:
Участвует в поддержании формы клетки.
Участвует в формировании межклеточных контактов.
Обеспечивает мембранные
Субмембранный комплекс
Функции:
Участвует в поддержании формы клетки.
Участвует в формировании межклеточных контактов.
Обеспечивает мембранные

Мобильность липидов
При температуре тела мембрана текуча и
липиды свободно перемещаются
Мобильность липидов
При температуре тела мембрана текуча и
липиды свободно перемещаются

Перемещение мембранных белков
Белковые молекулы мозаично распределены в липидном бислое и перемещаться
Перемещение мембранных белков
Белковые молекулы мозаично распределены в липидном бислое и перемещаться
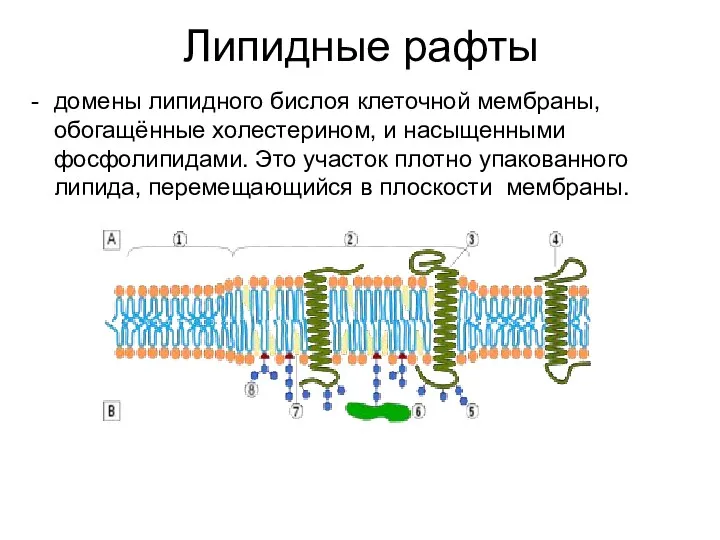
Липидные рафты
домены липидного бислоя клеточной мембраны, обогащённые холестерином, и насыщенными фосфолипидами.
Липидные рафты
домены липидного бислоя клеточной мембраны, обогащённые холестерином, и насыщенными фосфолипидами.
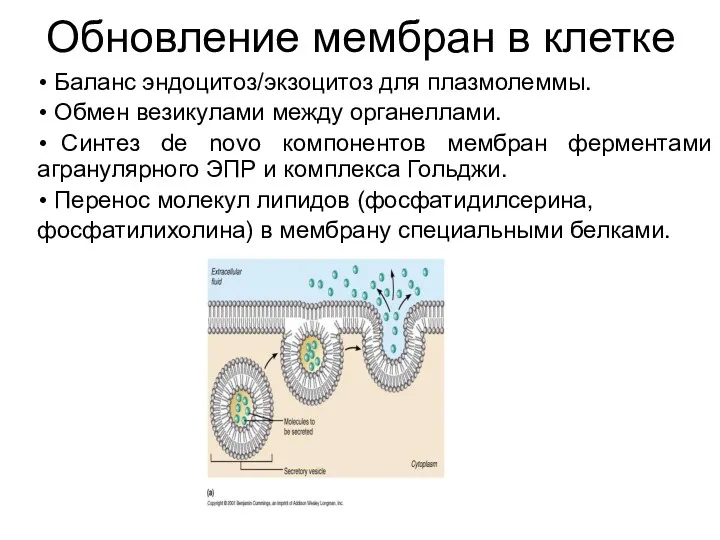
Обновление мембран в клетке
Баланс эндоцитоз/экзоцитоз для плазмолеммы.
Обмен везикулами между
Обновление мембран в клетке
Баланс эндоцитоз/экзоцитоз для плазмолеммы.
Обмен везикулами между

Функции плазмолеммы:
барьерная;
транспорт веществ в клетку и из клетки;
взаимодействие с сигнальными молекулами;
взаимодействие
Функции плазмолеммы:
барьерная;
транспорт веществ в клетку и из клетки;
взаимодействие с сигнальными молекулами;
взаимодействие

Рецепторы
гликопротеины, способные высокоселективно связываться с определенными молекулами - лигандами (гормон,
Рецепторы
гликопротеины, способные высокоселективно связываться с определенными молекулами - лигандами (гормон,
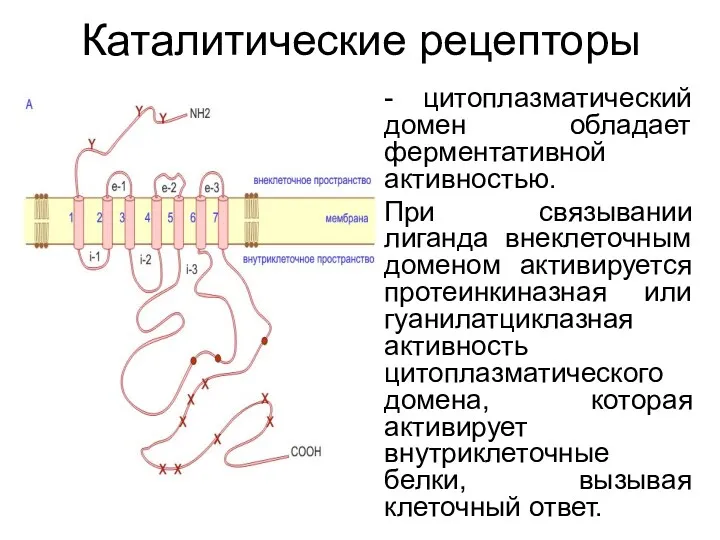
Каталитические рецепторы
- цитоплазматический домен обладает ферментативной активностью.
При связывании лиганда внеклеточным доменом
Каталитические рецепторы
- цитоплазматический домен обладает ферментативной активностью.
При связывании лиганда внеклеточным доменом
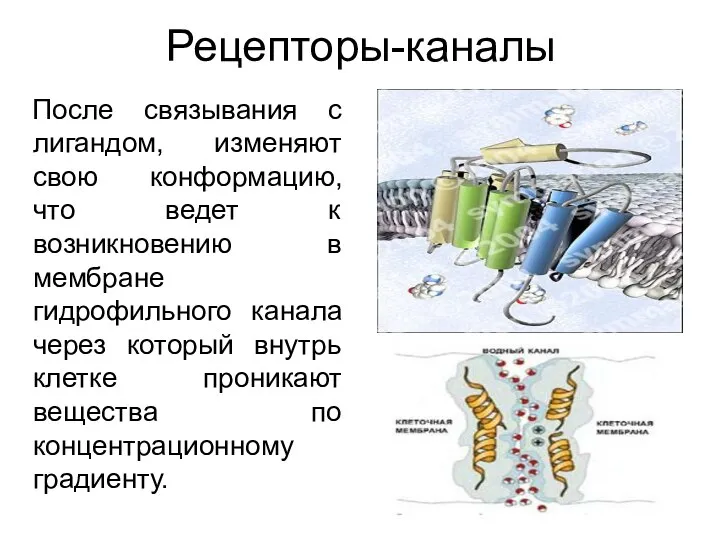
Рецепторы-каналы
После связывания с лигандом, изменяют свою конформацию, что ведет к возникновению
Рецепторы-каналы
После связывания с лигандом, изменяют свою конформацию, что ведет к возникновению
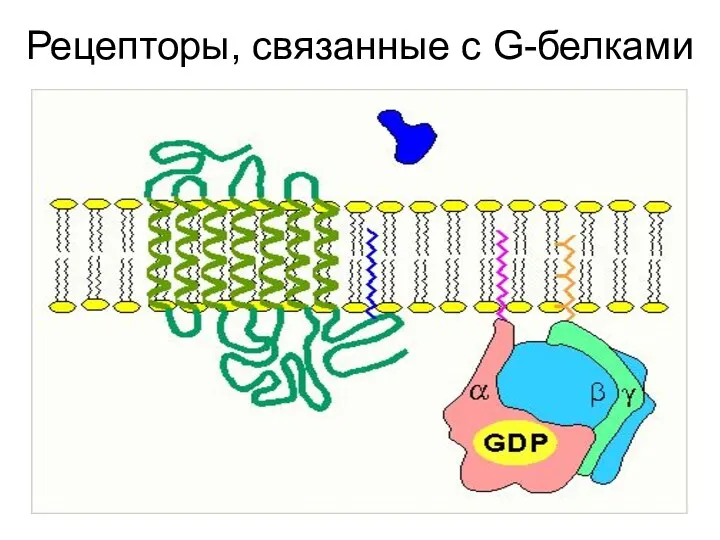
Рецепторы, связанные с G-белками
Рецепторы, связанные с G-белками

Трансмембранный транспорт веществ
прямой (диффузия, осмос, фильтрация);
опосредованный:
1. пассивный (с участием белка-переносчика
Трансмембранный транспорт веществ
прямой (диффузия, осмос, фильтрация);
опосредованный:
1. пассивный (с участием белка-переносчика

Простая диффузия
- переход вещества из области более высокой концентрации в область
Простая диффузия
- переход вещества из области более высокой концентрации в область
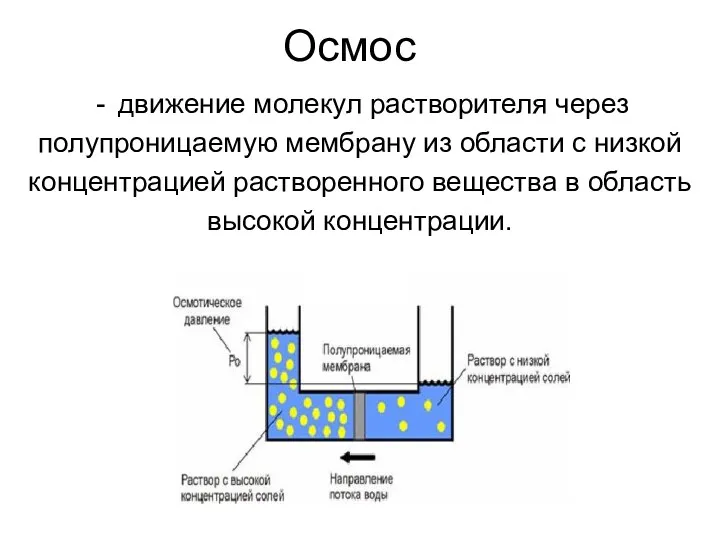
Осмос
движение молекул растворителя через
полупроницаемую мембрану из области с низкой
концентрацией
Осмос
движение молекул растворителя через
полупроницаемую мембрану из области с низкой
концентрацией
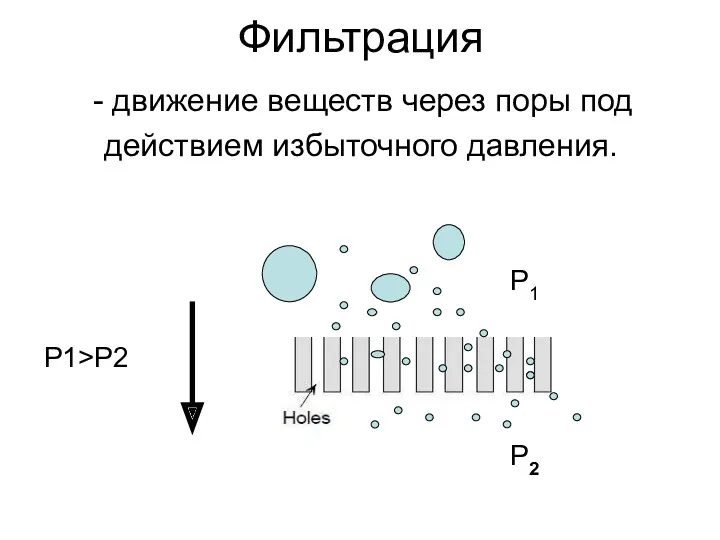
Фильтрация
движение веществ через поры под
действием избыточного давления.
Фильтрация
движение веществ через поры под
действием избыточного давления.
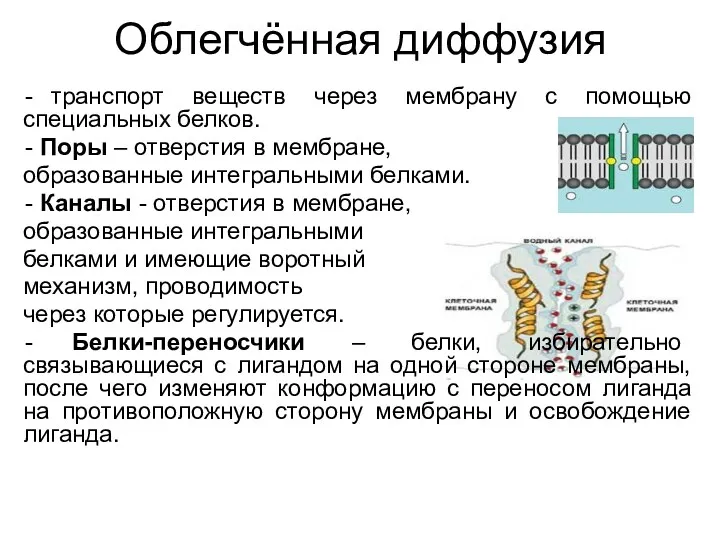
Облегчённая диффузия
транспорт веществ через мембрану с помощью специальных белков.
Облегчённая диффузия
транспорт веществ через мембрану с помощью специальных белков.
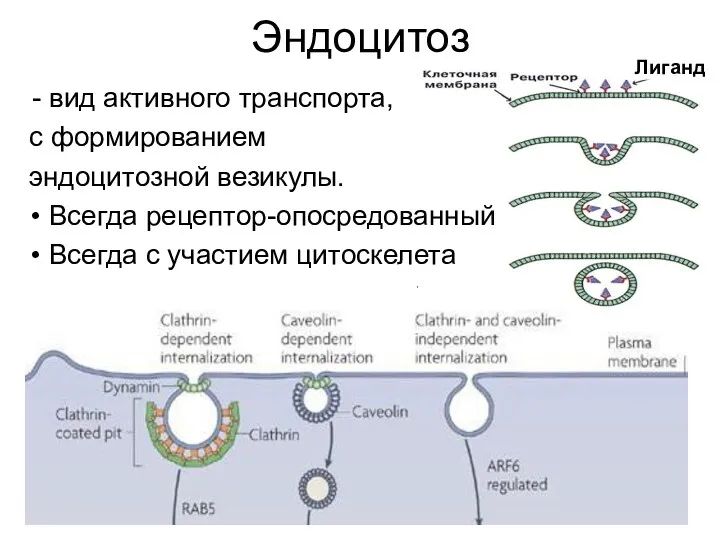
Эндоцитоз
вид активного транспорта,
с формированием
эндоцитозной везикулы.
Всегда рецептор-опосредованный
Всегда
Эндоцитоз
вид активного транспорта,
с формированием
эндоцитозной везикулы.
Всегда рецептор-опосредованный
Всегда

Виды эндоцитоза
Фагоцитоз - процесс поглощения клеткой объектов размером >10-6 м,
Виды эндоцитоза
Фагоцитоз - процесс поглощения клеткой объектов размером >10-6 м,
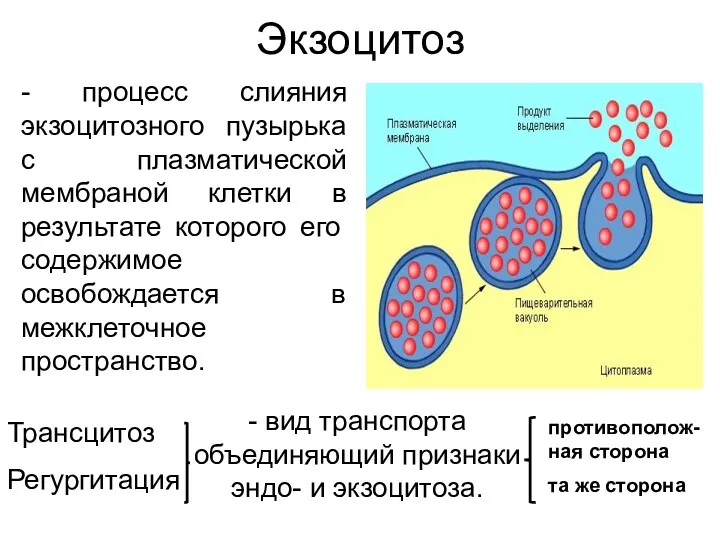
Экзоцитоз
- процесс слияния экзоцитозного пузырька с плазматической мембраной клетки в результате
Экзоцитоз
- процесс слияния экзоцитозного пузырька с плазматической мембраной клетки в результате
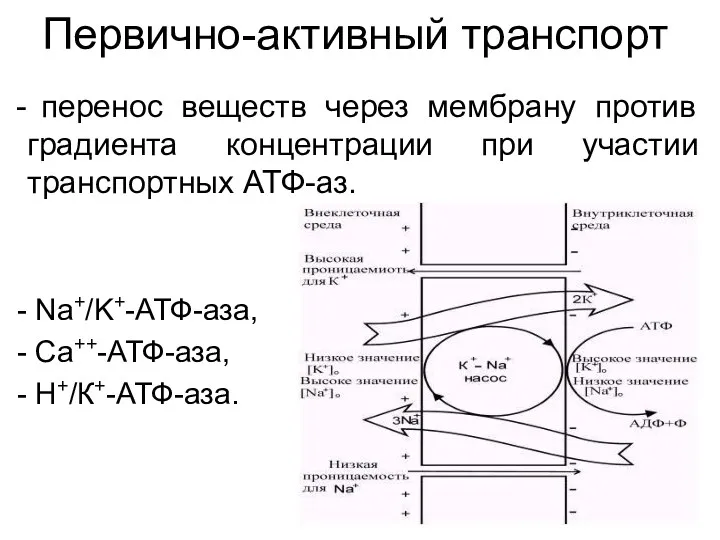
Первично-активный транспорт
перенос веществ через мембрану против градиента концентрации при участии
Первично-активный транспорт
перенос веществ через мембрану против градиента концентрации при участии
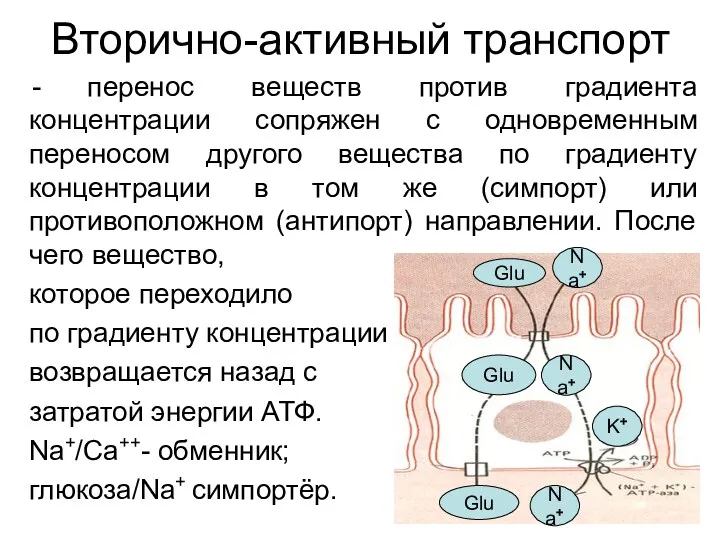
Вторично-активный транспорт
перенос веществ против градиента концентрации сопряжен с одновременным переносом
Вторично-активный транспорт
перенос веществ против градиента концентрации сопряжен с одновременным переносом

Межклеточные контакты
Механические – контакты, которые обеспечивают механическую связь клеток
Межклеточные контакты
Механические – контакты, которые обеспечивают механическую связь клеток

Интердигитации
Наиболее просто устроенный вид взаимодействия мембран смежных клеток. Представлены выпячиваниями цитоплазмы
Интердигитации
Наиболее просто устроенный вид взаимодействия мембран смежных клеток. Представлены выпячиваниями цитоплазмы
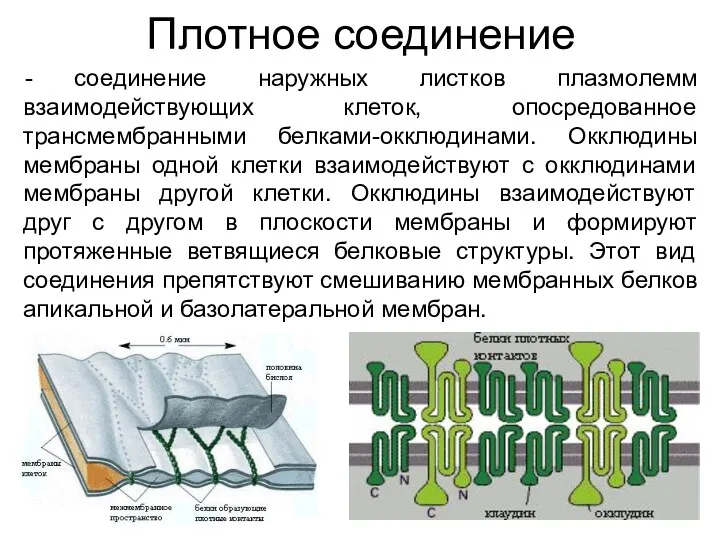
Плотное соединение
соединение наружных листков плазмолемм взаимодействующих клеток, опосредованное трансмембранными
Плотное соединение
соединение наружных листков плазмолемм взаимодействующих клеток, опосредованное трансмембранными

Промежуточное соединение, опоясывающая десмосома
Охватывают клетку по периметру в виде пояса. С
Промежуточное соединение, опоясывающая десмосома
Охватывают клетку по периметру в виде пояса. С
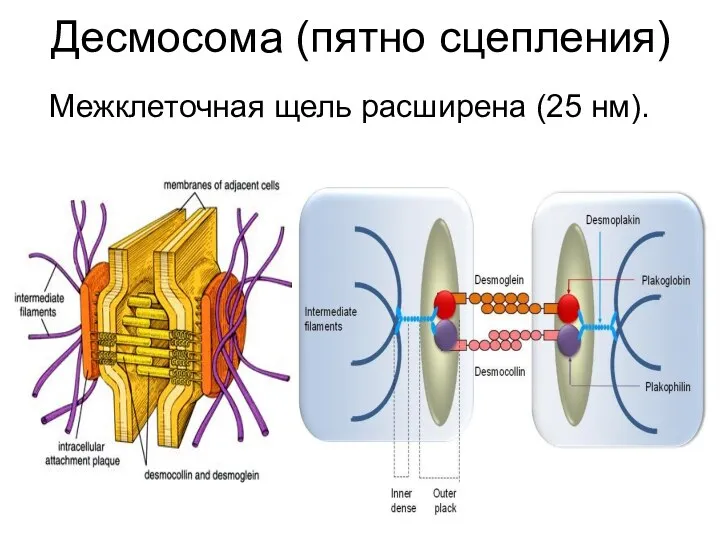
Десмосома (пятно сцепления)
Межклеточная щель расширена (25 нм).
Десмосома (пятно сцепления)
Межклеточная щель расширена (25 нм).
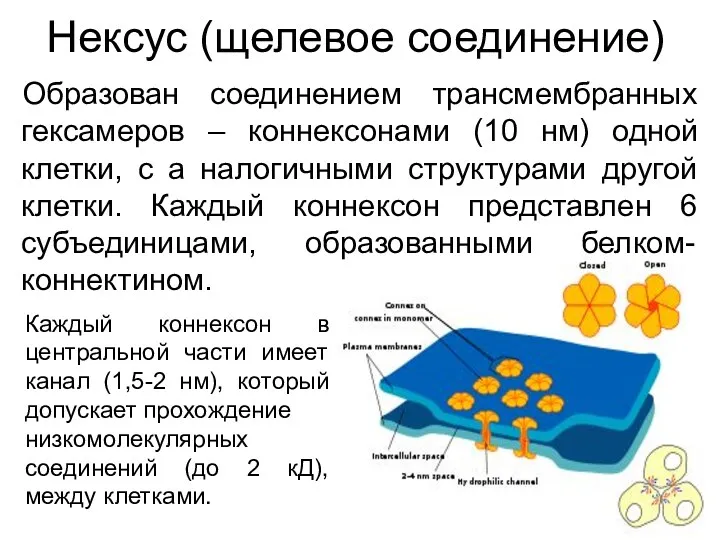
Нексус (щелевое соединение)
Образован соединением трансмембранных гексамеров – коннексонами (10 нм) одной
Нексус (щелевое соединение)
Образован соединением трансмембранных гексамеров – коннексонами (10 нм) одной
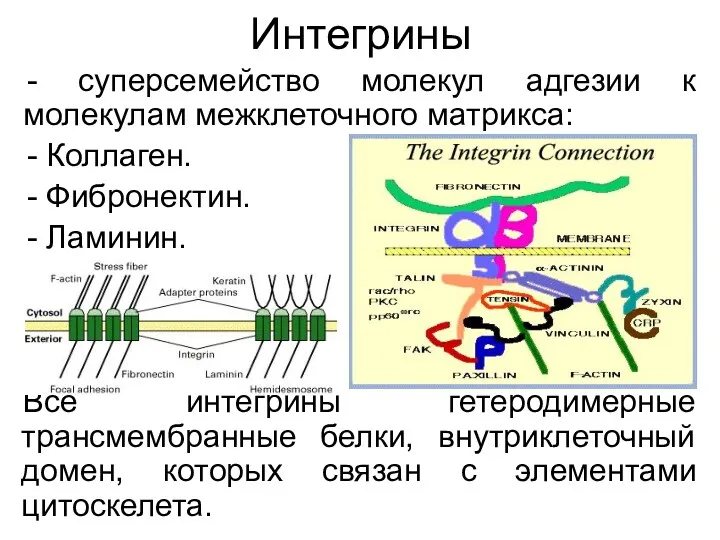
Интегрины
суперсемейство молекул адгезии к молекулам межклеточного матрикса:
Коллаген.
Фибронектин.
Ламинин.
Все
Интегрины
суперсемейство молекул адгезии к молекулам межклеточного матрикса:
Коллаген.
Фибронектин.
Ламинин.
Все
 Биохимия витаминов. (Лекция 25)
Биохимия витаминов. (Лекция 25) Мхи, папоротники, плауны, хвощи
Мхи, папоротники, плауны, хвощи Презентация Клеточная теория. Структура клетки
Презентация Клеточная теория. Структура клетки Внутренняя среда организма человека
Внутренняя среда организма человека Кровь и кровообращение. Кровеносная система человека
Кровь и кровообращение. Кровеносная система человека Проект Покормите птиц зимой
Проект Покормите птиц зимой Тесты по биологии
Тесты по биологии Биологиялық мембраналар. Биологиялық мембрана арқылы электролит емес заттардың тасымалы. Белсенді емес және белсенді тасымал
Биологиялық мембраналар. Биологиялық мембрана арқылы электролит емес заттардың тасымалы. Белсенді емес және белсенді тасымал Карантинные сорняки
Карантинные сорняки Вівця Доллі - 7
Вівця Доллі - 7 Обмін білків
Обмін білків Транскрипция. Регуляция транскрипции
Транскрипция. Регуляция транскрипции Класс Насекомые
Класс Насекомые Пищевые добавки в продуктах питания и их влияние на здоровье человека
Пищевые добавки в продуктах питания и их влияние на здоровье человека Характеристика семейства Бобовые
Характеристика семейства Бобовые Мышцы туловища и конечностей
Мышцы туловища и конечностей Онтогенез развития органов и систем
Онтогенез развития органов и систем Фотосинтез туралы мәлімет
Фотосинтез туралы мәлімет Розвиток життя у протерозоську еру
Розвиток життя у протерозоську еру Голос. Голосовой аппарат
Голос. Голосовой аппарат Обмен белков
Обмен белков органы и системы органов
органы и системы органов Белки (10 класс)
Белки (10 класс) Зона степей
Зона степей Бактерії - прокаріотичні одноклітинні мікроорганізми
Бактерії - прокаріотичні одноклітинні мікроорганізми Эон Фанерозой. Кайнозойская эра
Эон Фанерозой. Кайнозойская эра Царство грибы fungi (mycota)
Царство грибы fungi (mycota) Тип членистоногие. Классы ракообразные и паукообразные
Тип членистоногие. Классы ракообразные и паукообразные