- G-белки. Понятие и классификация. G-белок сопряженные рецепторы

Содержание
- 2. G-БЕЛКИ. ПОНЯТИЕ И КЛАССИФИКАЦИЯ G-белки (англ. G proteins) — это семейство белков, относящихся к ГТФазам и
- 3. ТИПЫ G-БЕЛКОВ G-белки делятся на две основных группы — гетеротримерные («большие») и «малые». Гетеротримерные G-белки —
- 4. БЕЛКИ-ПОМОЩНИКИ G-БЕЛКОВ В работе многих G-белков участвуют вспомогательные белки. GAPs (GTPase Activating Proteins, белки-активаторы ГТФазной активности)
- 6. Цикл активации G-белка под действием G-белок-связанного рецептора.
- 7. G-БЕЛОК СВЯЗАННЫЕ РЕЦЕПТОРЫ Рецепторы, сопряженные с G белком (серпетиновые рецепторы)(англ. G-protein-coupled receptors, GPCRs), также известные как
- 8. ЛИГАНДЫ И ЛИГАНД-СВЯЗЫВАЮЩИЕ УЧАСТКИ СЕРПЕНТИНОВЫХ РЕЦЕПТОРОВ Уникальная структура лиганд-связывающих участков серпентиновых рецепторов позволяет связывать лиганды различной
- 9. КЛАССИФИКАЦИЯ G-БЕЛОК СОПРЯЖЕННЫХ РЕЦЕПТОРОВ Семейство GPCR подразделяют на 6 классов на основании гомологии их аминокислотных последовательностей
- 10. СТРУКТУРА РЕЦЕПТОРА G-БЕЛОК СОПРЯЖЕННЫХ РЕЦЕПТОРОВ Внеклеточная часть состоит из петель, в которых среди прочих остатков содержатся
- 11. СТРУКТУРА РЕЦЕПТОРА G-БЕЛОК СОПРЯЖЕННЫХ РЕЦЕПТОРОВ В 2000 году была получена структура первого GPCR млекопитающих — бычьего
- 12. АКТИЦВАЦИЯ G-БЕЛОК СОПРЯЖЕННЫХ РЕЦЕПТОРОВ С-концевой участок рецептора локализован на цитоплазматической стороне плазматической мембраны и содержит высококонсервативные
- 13. АКТИЦВАЦИЯ G-БЕЛОК СОПРЯЖЕННЫХ РЕЦЕПТОРОВ До активации все три субъединицы связаны вместе. Когда рецептор активируется присоединением лиганда,
- 14. АЦЕТИЛХОЛИН ИНДУЦИРУЕТ ОТКРЫТИЕ К+ КАНАЛОВ В МЕМБРАНЕ (КЛЕТОК СЕРДЕЧНОЙ МЫШЦЫ) Связывание АХ с мускариновым АХ-рецептором активирует
- 15. РЕГУЛЯЦИЯ G-БЕЛОК-ЗАВИСИМЫХ РЕЦЕПТОРОВ Рецепторы, связанные с G-белками теряют чувствительность после длительной экспозиции со своими лигандами. Различают
- 16. РЕГУЛЯЦИЯ G-БЕЛОК-ЗАВИСИМЫХ РЕЦЕПТОРОВ Фосфорилирование цАМФ-зависимыми протеинкиназами цАМФ-зависимые киназы (протеинкиназа А) активируются цепью сигналов с G-белка (который
- 17. РЕГУЛЯЦИЯ G-БЕЛОК-ЗАВИСИМЫХ РЕЦЕПТОРОВ Фосфорилирование GRK-киназами. Киназы рецепторов, связанных с G-белками (GRK-киназы) — это протеинкиназы, фосфорилирующие лишь
- 18. ФИЗИОЛОГИЧЕСКАЯ РОЛЬ G-БЕЛОК СОПРЯЖЕННЫХ РЕЦЕПТОРОВ Рецепторы, связанные с G-белками вовлечены в широкий круг физиологических процессов. Вот
- 19. ФИЗИОЛОГИЧЕСКАЯ РОЛЬ G-БЕЛОК СОПРЯЖЕННЫХ РЕЦЕПТОРОВ 4. регуляция активности иммунной системы и воспаления: хемокиновые рецепторы связывают лиганды,
- 20. ПРИМЕРЫ G-БЕЛОК СОПРЯЖЕННЫХ РЕЦЕПТОРОВ 1.Карта белка рецептора мускаринового Ацетилхолиновый мускариновый рецептор относится к классу серпентиновых рецепторов
- 21. ПРИМЕРЫ G-БЕЛОК СОПРЯЖЕННЫХ РЕЦЕПТОРОВ Показано также, что ингибирование M2 рецептора подавляет активность аденилатциклазы , приводя к
- 22. ПРИМЕРЫ G-БЕЛОК СОПРЯЖЕННЫХ РЕЦЕПТОРОВ 2.Карта белка рецептора адренергического бета При молекулярном клонировании гена и кДНК бета-
- 23. ПРИМЕРЫ G-БЕЛОК СОПРЯЖЕННЫХ РЕЦЕПТОРОВ 3.Рецепторы, связывающие G-белки: гликозилирование Следует отметить, что NH2 концевой участок рецепторов, связывающих
- 24. РЕЦЕПТОРЫ ГОРМОНОВ СОПРЯЖЕННЫЕ С G-БЕЛКАМИ Многие гормоны активируют рецепторы, которые регулируют активность белков клеток- мишеней (например,
- 25. РЕЦЕПТОРЫ ГОРМОНОВ СОПРЯЖЕННЫЕ С G-БЕЛКАМИ Когда лиганд (гормон) взаимодействует с внеклеточной частью рецептора, в нем происходят
- 26. Спасибо за внимание!!!
- 28. Скачать презентацию

G-БЕЛКИ. ПОНЯТИЕ И КЛАССИФИКАЦИЯ
G-белки (англ. G proteins) — это семейство белков,
G-БЕЛКИ. ПОНЯТИЕ И КЛАССИФИКАЦИЯ
G-белки (англ. G proteins) — это семейство белков,

ТИПЫ G-БЕЛКОВ
G-белки делятся на две основных группы — гетеротримерные («большие») и
ТИПЫ G-БЕЛКОВ
G-белки делятся на две основных группы — гетеротримерные («большие») и

БЕЛКИ-ПОМОЩНИКИ G-БЕЛКОВ
В работе многих G-белков участвуют вспомогательные белки.
GAPs (GTPase Activating
БЕЛКИ-ПОМОЩНИКИ G-БЕЛКОВ
В работе многих G-белков участвуют вспомогательные белки.
GAPs (GTPase Activating
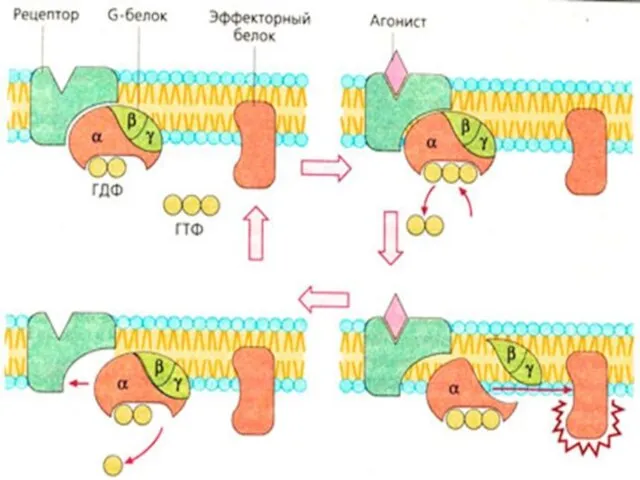
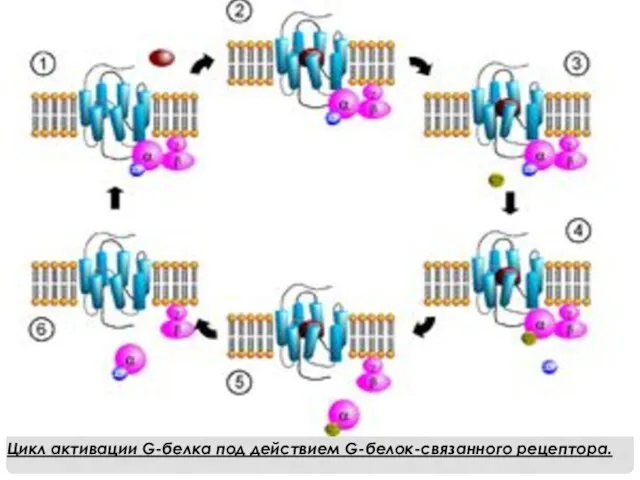
Цикл активации G-белка под действием G-белок-связанного рецептора.
Цикл активации G-белка под действием G-белок-связанного рецептора.

G-БЕЛОК СВЯЗАННЫЕ РЕЦЕПТОРЫ
Рецепторы, сопряженные с G белком (серпетиновые рецепторы)(англ. G-protein-coupled receptors,
G-БЕЛОК СВЯЗАННЫЕ РЕЦЕПТОРЫ
Рецепторы, сопряженные с G белком (серпетиновые рецепторы)(англ. G-protein-coupled receptors,
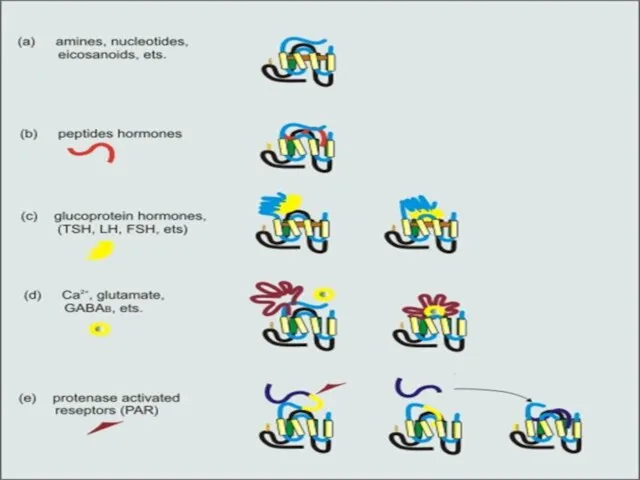
ЛИГАНДЫ И ЛИГАНД-СВЯЗЫВАЮЩИЕ УЧАСТКИ СЕРПЕНТИНОВЫХ РЕЦЕПТОРОВ
Уникальная структура лиганд-связывающих участков серпентиновых рецепторов
ЛИГАНДЫ И ЛИГАНД-СВЯЗЫВАЮЩИЕ УЧАСТКИ СЕРПЕНТИНОВЫХ РЕЦЕПТОРОВ
Уникальная структура лиганд-связывающих участков серпентиновых рецепторов

КЛАССИФИКАЦИЯ G-БЕЛОК СОПРЯЖЕННЫХ РЕЦЕПТОРОВ
Семейство GPCR подразделяют на 6 классов на основании
КЛАССИФИКАЦИЯ G-БЕЛОК СОПРЯЖЕННЫХ РЕЦЕПТОРОВ
Семейство GPCR подразделяют на 6 классов на основании

СТРУКТУРА РЕЦЕПТОРА G-БЕЛОК СОПРЯЖЕННЫХ РЕЦЕПТОРОВ
Внеклеточная часть состоит из петель, в которых
СТРУКТУРА РЕЦЕПТОРА G-БЕЛОК СОПРЯЖЕННЫХ РЕЦЕПТОРОВ
Внеклеточная часть состоит из петель, в которых

СТРУКТУРА РЕЦЕПТОРА G-БЕЛОК СОПРЯЖЕННЫХ РЕЦЕПТОРОВ
В 2000 году была получена структура первого
СТРУКТУРА РЕЦЕПТОРА G-БЕЛОК СОПРЯЖЕННЫХ РЕЦЕПТОРОВ
В 2000 году была получена структура первого

АКТИЦВАЦИЯ G-БЕЛОК СОПРЯЖЕННЫХ РЕЦЕПТОРОВ
С-концевой участок рецептора локализован на цитоплазматической стороне плазматической
АКТИЦВАЦИЯ G-БЕЛОК СОПРЯЖЕННЫХ РЕЦЕПТОРОВ
С-концевой участок рецептора локализован на цитоплазматической стороне плазматической

АКТИЦВАЦИЯ G-БЕЛОК СОПРЯЖЕННЫХ РЕЦЕПТОРОВ
До активации все три субъединицы связаны вместе. Когда
АКТИЦВАЦИЯ G-БЕЛОК СОПРЯЖЕННЫХ РЕЦЕПТОРОВ
До активации все три субъединицы связаны вместе. Когда
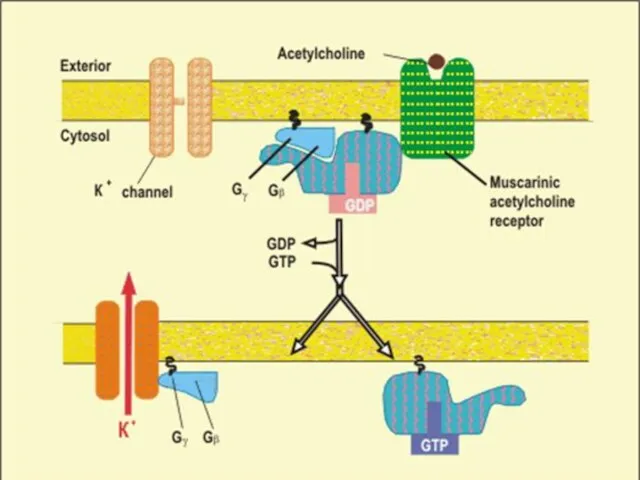
АЦЕТИЛХОЛИН ИНДУЦИРУЕТ ОТКРЫТИЕ К+ КАНАЛОВ В МЕМБРАНЕ (КЛЕТОК СЕРДЕЧНОЙ МЫШЦЫ)
Связывание АХ
АЦЕТИЛХОЛИН ИНДУЦИРУЕТ ОТКРЫТИЕ К+ КАНАЛОВ В МЕМБРАНЕ (КЛЕТОК СЕРДЕЧНОЙ МЫШЦЫ)
Связывание АХ

РЕГУЛЯЦИЯ G-БЕЛОК-ЗАВИСИМЫХ РЕЦЕПТОРОВ
Рецепторы, связанные с G-белками теряют чувствительность после длительной экспозиции
РЕГУЛЯЦИЯ G-БЕЛОК-ЗАВИСИМЫХ РЕЦЕПТОРОВ
Рецепторы, связанные с G-белками теряют чувствительность после длительной экспозиции

РЕГУЛЯЦИЯ G-БЕЛОК-ЗАВИСИМЫХ РЕЦЕПТОРОВ
Фосфорилирование цАМФ-зависимыми протеинкиназами
цАМФ-зависимые киназы (протеинкиназа А) активируются цепью сигналов
РЕГУЛЯЦИЯ G-БЕЛОК-ЗАВИСИМЫХ РЕЦЕПТОРОВ
Фосфорилирование цАМФ-зависимыми протеинкиназами
цАМФ-зависимые киназы (протеинкиназа А) активируются цепью сигналов

РЕГУЛЯЦИЯ G-БЕЛОК-ЗАВИСИМЫХ РЕЦЕПТОРОВ
Фосфорилирование GRK-киназами.
Киназы рецепторов, связанных с G-белками (GRK-киназы) — это
РЕГУЛЯЦИЯ G-БЕЛОК-ЗАВИСИМЫХ РЕЦЕПТОРОВ
Фосфорилирование GRK-киназами.
Киназы рецепторов, связанных с G-белками (GRK-киназы) — это

ФИЗИОЛОГИЧЕСКАЯ РОЛЬ G-БЕЛОК СОПРЯЖЕННЫХ РЕЦЕПТОРОВ
Рецепторы, связанные с G-белками вовлечены в широкий
ФИЗИОЛОГИЧЕСКАЯ РОЛЬ G-БЕЛОК СОПРЯЖЕННЫХ РЕЦЕПТОРОВ
Рецепторы, связанные с G-белками вовлечены в широкий

ФИЗИОЛОГИЧЕСКАЯ РОЛЬ G-БЕЛОК СОПРЯЖЕННЫХ РЕЦЕПТОРОВ
4. регуляция активности иммунной системы и воспаления: хемокиновые
ФИЗИОЛОГИЧЕСКАЯ РОЛЬ G-БЕЛОК СОПРЯЖЕННЫХ РЕЦЕПТОРОВ
4. регуляция активности иммунной системы и воспаления: хемокиновые

ПРИМЕРЫ G-БЕЛОК СОПРЯЖЕННЫХ РЕЦЕПТОРОВ
1.Карта белка рецептора мускаринового
Ацетилхолиновый мускариновый рецептор относится к
ПРИМЕРЫ G-БЕЛОК СОПРЯЖЕННЫХ РЕЦЕПТОРОВ
1.Карта белка рецептора мускаринового
Ацетилхолиновый мускариновый рецептор относится к

ПРИМЕРЫ G-БЕЛОК СОПРЯЖЕННЫХ РЕЦЕПТОРОВ
Показано также, что ингибирование M2 рецептора подавляет активность
ПРИМЕРЫ G-БЕЛОК СОПРЯЖЕННЫХ РЕЦЕПТОРОВ
Показано также, что ингибирование M2 рецептора подавляет активность

ПРИМЕРЫ G-БЕЛОК СОПРЯЖЕННЫХ РЕЦЕПТОРОВ
2.Карта белка рецептора адренергического бета
При молекулярном клонировании гена
ПРИМЕРЫ G-БЕЛОК СОПРЯЖЕННЫХ РЕЦЕПТОРОВ
2.Карта белка рецептора адренергического бета
При молекулярном клонировании гена

ПРИМЕРЫ G-БЕЛОК СОПРЯЖЕННЫХ РЕЦЕПТОРОВ
3.Рецепторы, связывающие G-белки: гликозилирование
Следует отметить, что NH2 концевой
ПРИМЕРЫ G-БЕЛОК СОПРЯЖЕННЫХ РЕЦЕПТОРОВ
3.Рецепторы, связывающие G-белки: гликозилирование
Следует отметить, что NH2 концевой

РЕЦЕПТОРЫ ГОРМОНОВ СОПРЯЖЕННЫЕ С G-БЕЛКАМИ
Многие гормоны активируют рецепторы, которые регулируют активность
РЕЦЕПТОРЫ ГОРМОНОВ СОПРЯЖЕННЫЕ С G-БЕЛКАМИ
Многие гормоны активируют рецепторы, которые регулируют активность

РЕЦЕПТОРЫ ГОРМОНОВ СОПРЯЖЕННЫЕ С G-БЕЛКАМИ
Когда лиганд (гормон) взаимодействует с внеклеточной частью
РЕЦЕПТОРЫ ГОРМОНОВ СОПРЯЖЕННЫЕ С G-БЕЛКАМИ
Когда лиганд (гормон) взаимодействует с внеклеточной частью

Спасибо за внимание!!!
Спасибо за внимание!!!
 Анатомия спинного мозга (medulla spinalis)
Анатомия спинного мозга (medulla spinalis) Устройство увеличительных приборов. 5 класс
Устройство увеличительных приборов. 5 класс Видоизменение побегов. Клубень
Видоизменение побегов. Клубень Редко встречающиеся виды краснокнижных растений
Редко встречающиеся виды краснокнижных растений Зимующие птицы
Зимующие птицы Растительный мир Алтайского края
Растительный мир Алтайского края Муравьи: семейство формицидов
Муравьи: семейство формицидов Птицы Рязанской области. Знакомство с уникальными видами птиц
Птицы Рязанской области. Знакомство с уникальными видами птиц Почему нужно есть много овощей и фруктов
Почему нужно есть много овощей и фруктов Единый государственный экзамен по биологии. Содержательные блоки курса биология
Единый государственный экзамен по биологии. Содержательные блоки курса биология Нейрон. Рефлекс. Рефлекторна дуга. Будова нервової системи. Нервова тканина
Нейрон. Рефлекс. Рефлекторна дуга. Будова нервової системи. Нервова тканина Особливості годівлі хутрових звірів
Особливості годівлі хутрових звірів Генетика микроорганизмов
Генетика микроорганизмов История развития анатомии, физиологии и медицины
История развития анатомии, физиологии и медицины Презентация, Обобщающий урок по теме скелет.
Презентация, Обобщающий урок по теме скелет. Презентация к КВН Зеленые друзья 6 класс
Презентация к КВН Зеленые друзья 6 класс Культивирование клеток. Лекция 3
Культивирование клеток. Лекция 3 Wild and domestic animals
Wild and domestic animals Проверочная работа по теме Органоиды клетки.
Проверочная работа по теме Органоиды клетки. Отряды насекомых
Отряды насекомых Факторы, лимитирующие первичную продукцию в наземных и водных сообществах
Факторы, лимитирующие первичную продукцию в наземных и водных сообществах Самоочищение водоема
Самоочищение водоема Жизнедеятельность растений
Жизнедеятельность растений Органи рослин
Органи рослин Ботаникалық сипаттамасы
Ботаникалық сипаттамасы Тип Хордовые. Подтип Бесчерепные. Подтип Черепные (Позвоночные)
Тип Хордовые. Подтип Бесчерепные. Подтип Черепные (Позвоночные) Первые современные люди - неоантропы
Первые современные люди - неоантропы Виявлення пристосувань до способу життя птахів. Практична робота 3
Виявлення пристосувань до способу життя птахів. Практична робота 3