- Генетическая информация. Репликация ДНК

Содержание
- 2. Молекулярная биология наука об информационных процессах в клетке, протекающих на молекулярном уровне
- 3. Потенциальное разнообразие белков огромно – 20n, где n – длина цепочки (в среднем 300 а.к.) Каждый
- 4. вся информация о строении клеток и организма в целом, записанная в молекулах ДНК Необходима для синтеза
- 5. Матричный Усиления в результате многократного копирования Принципы передачи информации
- 6. Вспомним сначала, что нужно для синтеза регулярного полимера – например, полисахарида: А А А А +
- 7. 1. Матричный принцип Что нужно для синтеза НЕрегулярного полимера? А Т Т Ц + Все то
- 8. Молекула-матрица Молекула, у которой фермент «спрашивает» какой мономер ставить на следующее место в цепи Ответ –
- 9. Николай Константинович Кольцов 1872-1940 1927 Идея о молекулах-матрицах
- 10. Генонема Рисунок Н.К. Кольцова Кольцов считал, что матрицами могут быть белки. О ДНК тогда знали мало
- 11. 1952 Окончательно доказано, что носитель наследственной информации – ДНК (Херши, Чейз) ДНК – матрица Но она
- 12. Центральная догма ДНК РНК белок Матрицами могут быть только нуклеиновые кислоты
- 13. ДНК РНК белок Матричные синтезы, разрешенные по центральной догме Не обнаружен
- 14. Центральная догма ДНК РНК белок Репликация Транскрипция Трансляция Обратная транскрипция Репликация РНК Только РНК-вирусы Ретро-РНК-вирусы Другие
- 15. Запрещенные матричные синтезы Белки никогда не бывают матрицами
- 16. По матричному принципу синтезируются все нерегулярные полимеры: ДНК, РНК, белки. Но матрицами могут быть только нуклеиновые
- 17. Второй принцип матричных синтезов – принцип усиления В ходе копирования информации становится больше Зигота Развитие многокле-
- 18. Из-за принципа усиления изменения в молекуле ДНК реализуются на макроуровне Зигота Развитие многокле- точного организма Все
- 19. Репликация ДНК
- 20. Универсальный биологический процесс передачи генетической информации в поколениях клеток и организмов, благодаря созданию точных копий ДНК.
- 21. Место репликации в клеточном цикле Репликация ДНК всегда предшествует делению клетки. Репликация S-период (Synthesis) Интерфаза Деление
- 22. Принципы репликации 1. Комплементарность 2. Антипараллельность 3. Полуконсервативность 4. Униполярность 5. Прерывистость 6. Потребность в затравке
- 23. Полуконсервативность Полуконсервативный Консервативный Дисперсионный
- 24. Репликон – расстояние между двумя сайтами начала репликации ori ~ 100 тыс. н.п. У прокариот вся
- 25. Прерывистость репликации ДНК одной хромосомы ori ori Репликативные вилки
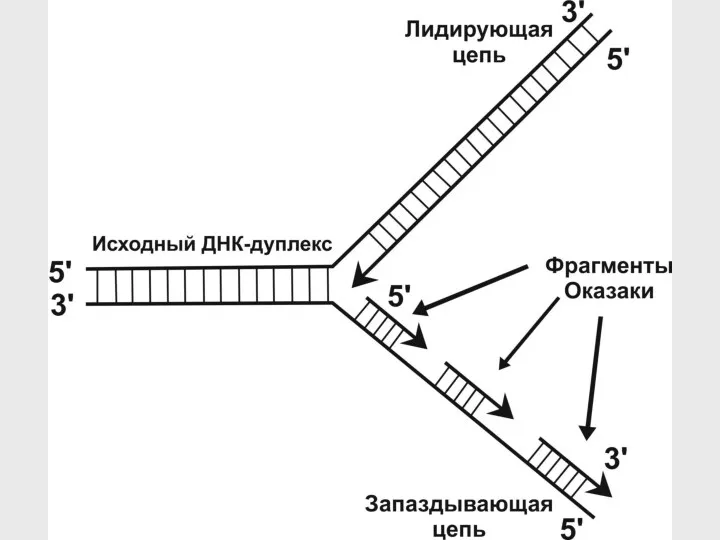
- 26. Прерывистость репликации
- 27. Прерывистость репликации ДНК одной хромосомы ori 3' 5' 3' 5' 5' 5' 3' 3' Противоречие с
- 28. Репликативная вилка Униполярность: Растущий конец новой цепочки – всегда 3' 3' 5' 3' 3' Запаздывающая цепь
- 29. Молекулярные машины Комплекс белков и ферментов, действующих согласованно Реплисома Рибосома Сплайсосома Протеасома Примеры молекулярных машин
- 30. Структура протеасомы Протеасома — это белковый комплекс, который занимается в клетке избирательной деградацией белков. Протеасома не
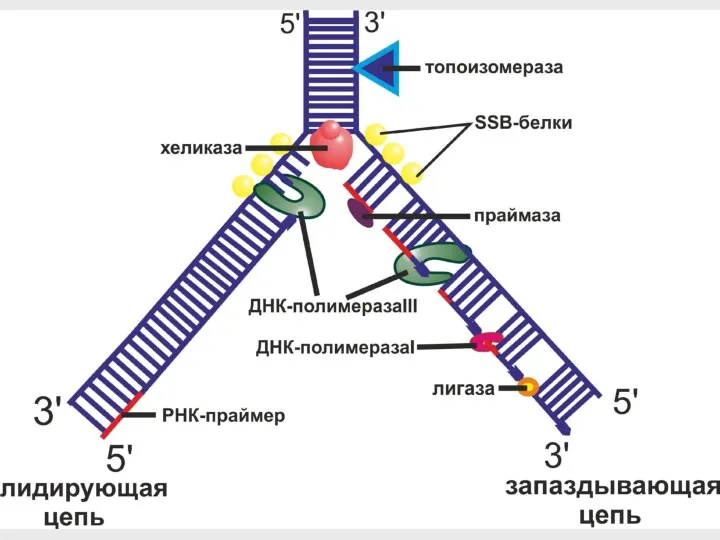
- 31. Молекулярная машина репликации
- 32. 1. Геликазы раскручивают двойную спираль
- 33. ДНК- полимераза праймаза Праймер РНК 2. Праймаза синтезирует РНК-затравку (праймер)
- 34. Удаление праймера 3. ДНК-полимераза III синтезирует новую цепь ДНК 4. ДНК-полимераза I удаляет праймер и заделывает
- 35. ДНК-полимераза использует нуклеотиды в виде 5' трифосфатов Растущий 3‘ конец цепочки Дезокси-нуклеотид трифосфат 5' 3' 5'
- 36. Свойства ДНК-полимеразы 1. Присоединяет по одному нуклеотиду с 3‘ конца растущей цепочки. 2. Требует для начала
- 37. ДНК-полимераза исправляет ошибки Если новый нуклеотид не спарен – фермент не может двигаться дальше. Тогда он
- 40. Скорость репликации ДНК У прокариот – 1000 нуклеотидов /сек У эукариот – 100 нуклеотидов /сек (медленнее,
- 41. Выводы по репликации ДНК В результате репликации каждая дочерняя клетка получает точную копию всей ДНК содержавшейся
- 42. Мутации и системы репарации
- 43. Частота ошибок ДНК-полимеразы ~1٠10– 9 – 1 нуклеотид на миллиард. Мутации – это случайные наследуемые изменения
- 44. Спонтанный уровень (внутренние причины мутаций): Ошибки репликации Ошибки деления клеток Перемещение мобильных элементов Мутагены – факторы
- 45. Системы репарации Белки, которые исправляют ошибки и повреждения в ДНК. Дефекты этих систем ведут к тяжелым
- 46. Мутации и репарация ДНК Генетическая информация кодируется последовательностью оснований ДНК и поэтому изменения в структуре или
- 47. Репарация ДНК Выделяют виды репараций: фотореактивация, темновая и SOS-реактивация. Фотореактивация защищает клетку от негативного действия ультрафиолетовой
- 48. Репарация ДНК Феномен темновой репарации сложнее предыдущего. Его сущность заключается в том, что особенные ферменты находят
- 49. Основные понятия по теме «Репликация» Место репликации в клеточном цикле Принципы репликации Лидирующая и запаздывающая цепи
- 50. Теломеры и теломераза Дополнение к теме
- 51. В зависимости от места положения центромеры различают: Равноплечие хромосомы; Неравноплечие хромосомы; Резко неравноплечие хромосомы; Одноплечие; Спутничные.
- 52. В хромосоме различают: 5 – первичную перетяжку; 6 – вторичную перетяжку (ядрышковый организатор); 7 – спутники
- 53. Проблема укорочения концов у линейных ДНК Сформулирована – А.М. Оловников, 1971 При каждой репликации новые цепи
- 54. Адрес рисунка http://www.uic.edu/classes/bios/bios100/f06pm/lect10.htm
- 55. Гипотеза Оловникова Укорочение концов – это внутренние часы, отмеряющие время жизни многоклеточного организма – число отпущенных
- 56. Но почему тогда клетки зародышевой линии делятся бесконечно? Оловников: должен существовать механизм удлинения концов хромосом. Теломераза
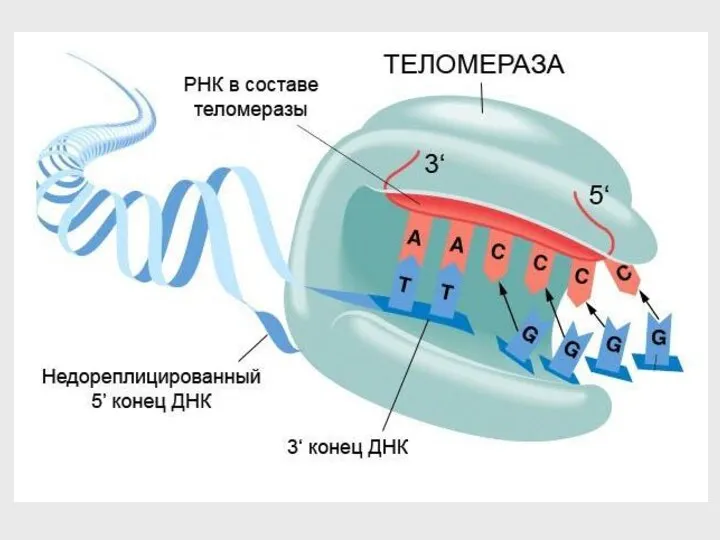
- 57. Теломераза фермент, надстраивающий концы хромосом, содержит РНК. удлинение происходит путем обратной транскрипции: РНК → ДНК На
- 61. Скачать презентацию

Молекулярная биология
наука об информационных процессах в клетке, протекающих на молекулярном
Молекулярная биология
наука об информационных процессах в клетке, протекающих на молекулярном

Потенциальное разнообразие белков огромно – 20n,
где n – длина цепочки
Потенциальное разнообразие белков огромно – 20n,
где n – длина цепочки

вся информация о строении клеток и организма в целом, записанная в
вся информация о строении клеток и организма в целом, записанная в

Матричный
Усиления в результате многократного копирования
Принципы передачи информации
Матричный
Усиления в результате многократного копирования
Принципы передачи информации
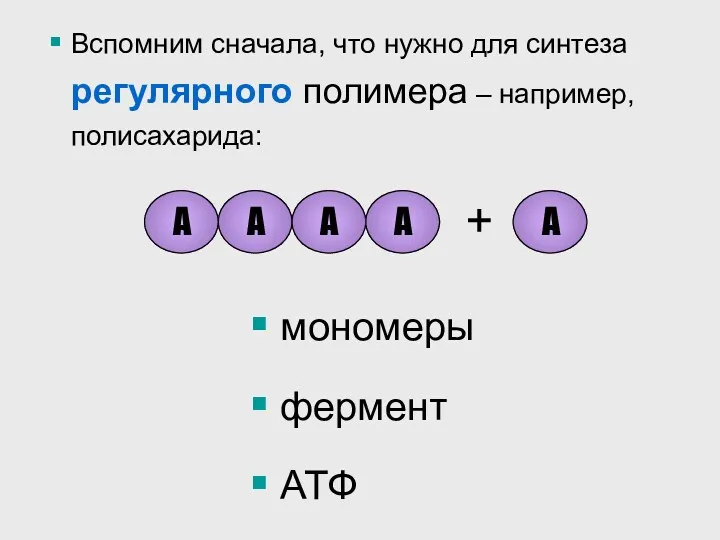
Вспомним сначала, что нужно для синтеза регулярного полимера – например, полисахарида:
А
А
А
А
+
Вспомним сначала, что нужно для синтеза регулярного полимера – например, полисахарида:
А
А
А
А
+
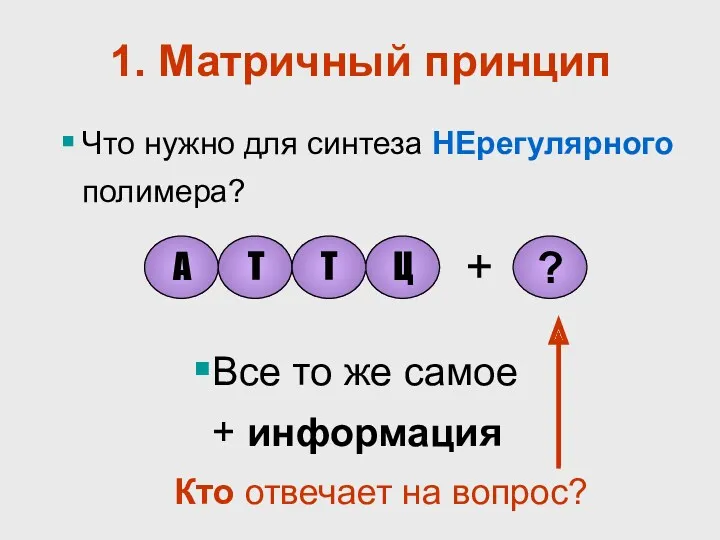
1. Матричный принцип
Что нужно для синтеза НЕрегулярного полимера?
А
Т
Т
Ц
+
Все то же самое
1. Матричный принцип
Что нужно для синтеза НЕрегулярного полимера?
А
Т
Т
Ц
+
Все то же самое

Молекула-матрица
Молекула, у которой фермент «спрашивает» какой мономер ставить на следующее место
Молекула-матрица
Молекула, у которой фермент «спрашивает» какой мономер ставить на следующее место

Николай Константинович Кольцов
1872-1940
1927
Идея о молекулах-матрицах
Николай Константинович Кольцов
1872-1940
1927
Идея о молекулах-матрицах
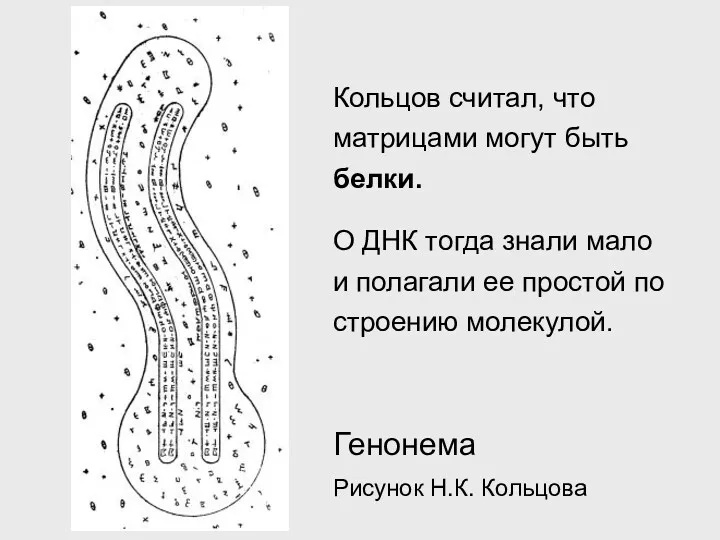
Генонема
Рисунок Н.К. Кольцова
Кольцов считал, что матрицами могут быть белки.
О ДНК
Генонема
Рисунок Н.К. Кольцова
Кольцов считал, что матрицами могут быть белки.
О ДНК

1952
Окончательно доказано, что носитель наследственной информации – ДНК (Херши, Чейз)
ДНК
1952
Окончательно доказано, что носитель наследственной информации – ДНК (Херши, Чейз)
ДНК

Центральная догма
ДНК
РНК
белок
Матрицами могут быть только нуклеиновые кислоты
Центральная догма
ДНК
РНК
белок
Матрицами могут быть только нуклеиновые кислоты
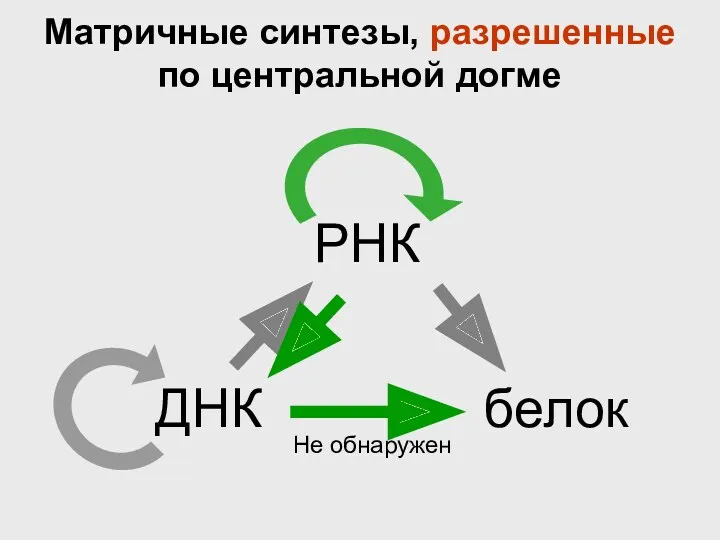
ДНК
РНК
белок
Матричные синтезы, разрешенные по центральной догме
Не обнаружен
ДНК
РНК
белок
Матричные синтезы, разрешенные по центральной догме
Не обнаружен
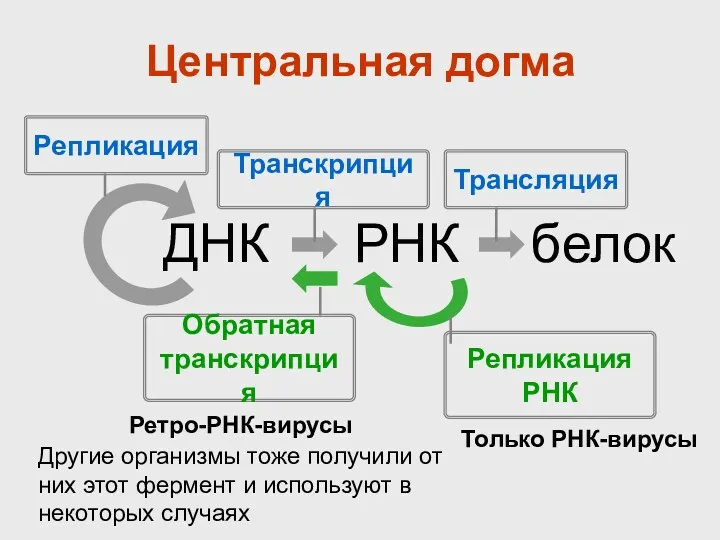
Центральная догма
ДНК
РНК
белок
Репликация
Транскрипция
Трансляция
Обратная
транскрипция
Репликация
РНК
Только РНК-вирусы
Ретро-РНК-вирусы
Другие организмы тоже получили от них этот фермент и
Центральная догма
ДНК
РНК
белок
Репликация
Транскрипция
Трансляция
Обратная
транскрипция
Репликация
РНК
Только РНК-вирусы
Ретро-РНК-вирусы
Другие организмы тоже получили от них этот фермент и
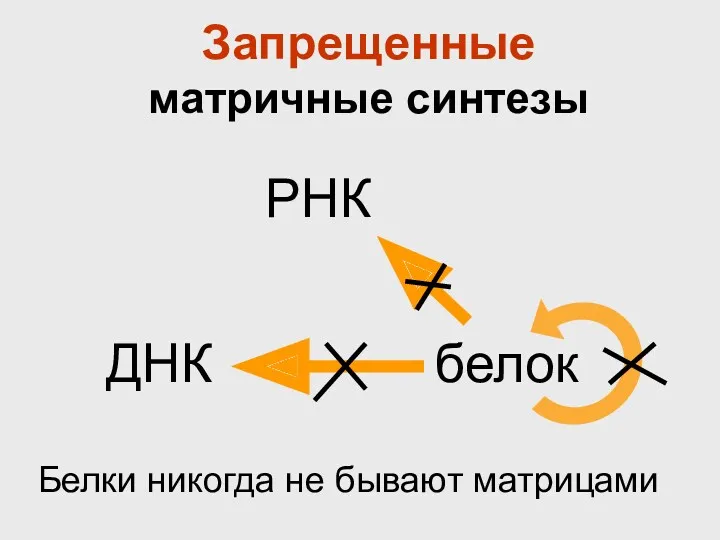
Запрещенные
матричные синтезы
Белки никогда не бывают матрицами
Запрещенные
матричные синтезы
Белки никогда не бывают матрицами

По матричному принципу синтезируются все нерегулярные полимеры: ДНК, РНК, белки.
Но матрицами
По матричному принципу синтезируются все нерегулярные полимеры: ДНК, РНК, белки.
Но матрицами
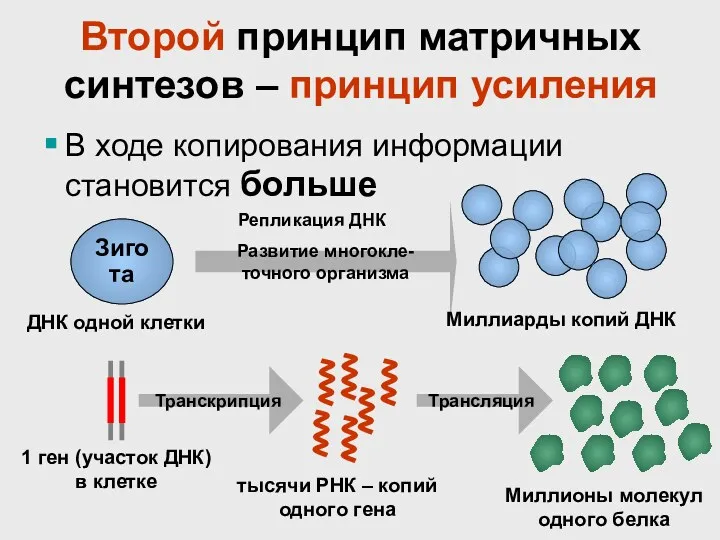
Второй принцип матричных синтезов – принцип усиления
В ходе копирования информации становится
Второй принцип матричных синтезов – принцип усиления
В ходе копирования информации становится
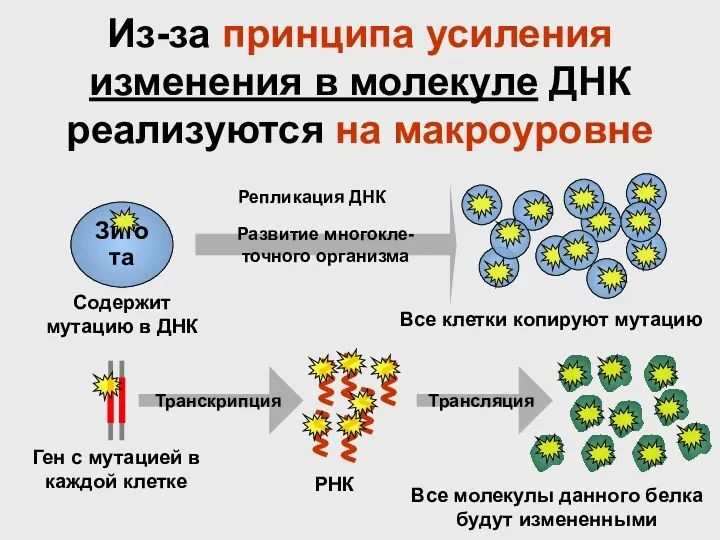
Из-за принципа усиления изменения в молекуле ДНК реализуются на макроуровне
Зигота
Развитие многокле-
точного
Из-за принципа усиления изменения в молекуле ДНК реализуются на макроуровне
Зигота
Развитие многокле-
точного

Репликация ДНК
Репликация ДНК

Универсальный биологический процесс передачи генетической информации в поколениях клеток и организмов,
Универсальный биологический процесс передачи генетической информации в поколениях клеток и организмов,
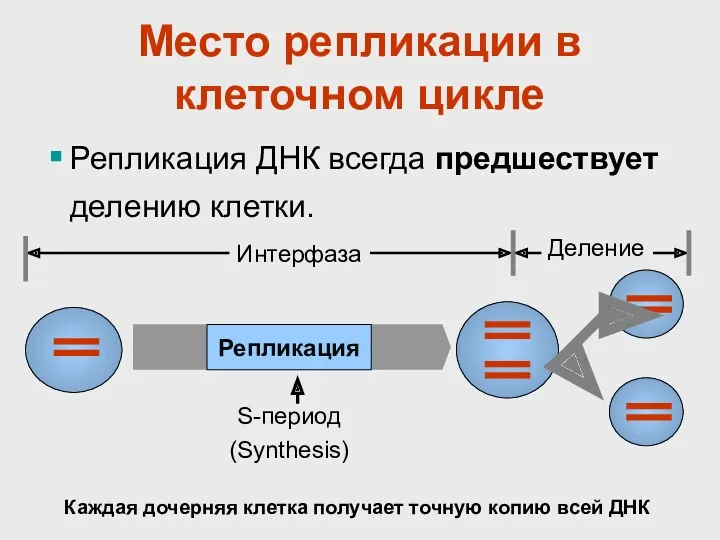
Место репликации в клеточном цикле
Репликация ДНК всегда предшествует делению клетки.
Репликация
S-период
(Synthesis)
Интерфаза
Деление
Каждая дочерняя
Место репликации в клеточном цикле
Репликация ДНК всегда предшествует делению клетки.
Репликация
S-период
(Synthesis)
Интерфаза
Деление
Каждая дочерняя

Принципы репликации
1. Комплементарность
2. Антипараллельность
3. Полуконсервативность
4. Униполярность
5. Прерывистость
6. Потребность в затравке
Принципы репликации
1. Комплементарность
2. Антипараллельность
3. Полуконсервативность
4. Униполярность
5. Прерывистость
6. Потребность в затравке
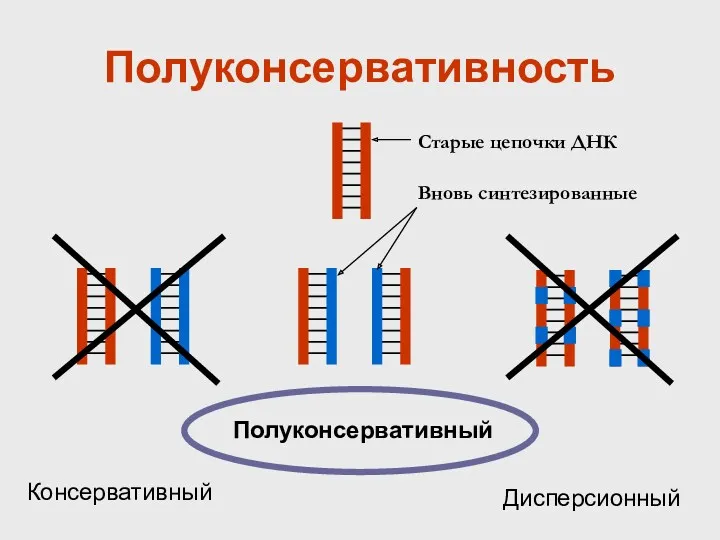
Полуконсервативность
Полуконсервативный
Консервативный
Дисперсионный
Полуконсервативность
Полуконсервативный
Консервативный
Дисперсионный
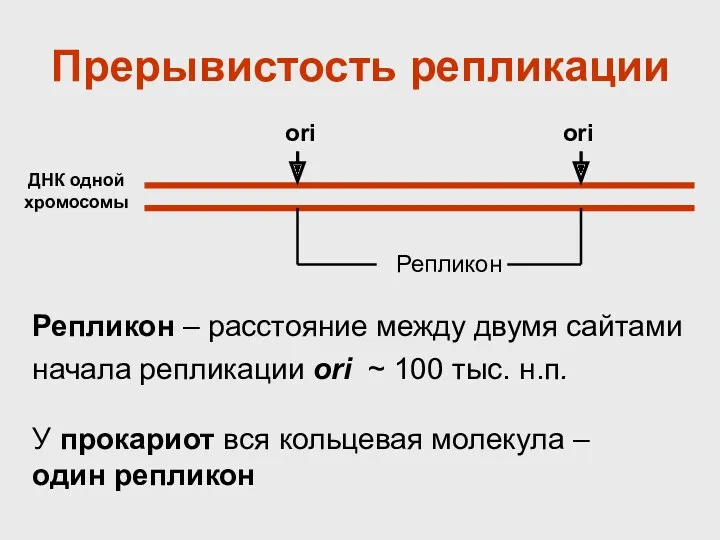
Репликон – расстояние между двумя сайтами начала репликации ori ~ 100
Репликон – расстояние между двумя сайтами начала репликации ori ~ 100
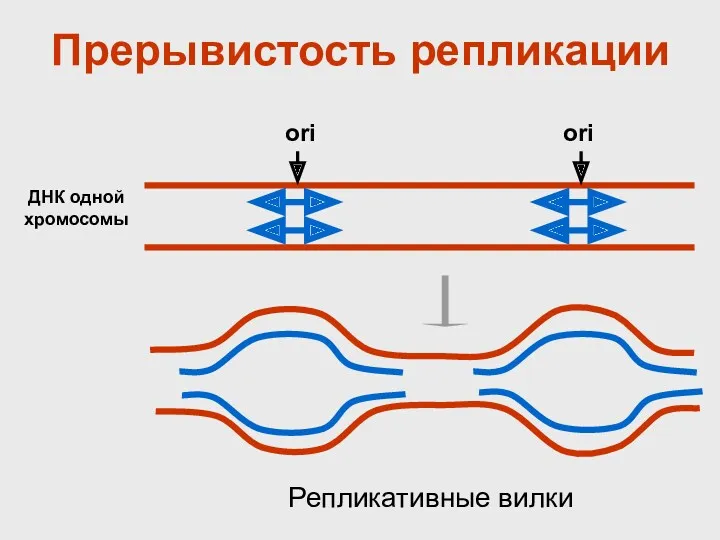
Прерывистость репликации
ДНК одной хромосомы
ori
ori
Репликативные вилки
Прерывистость репликации
ДНК одной хромосомы
ori
ori
Репликативные вилки
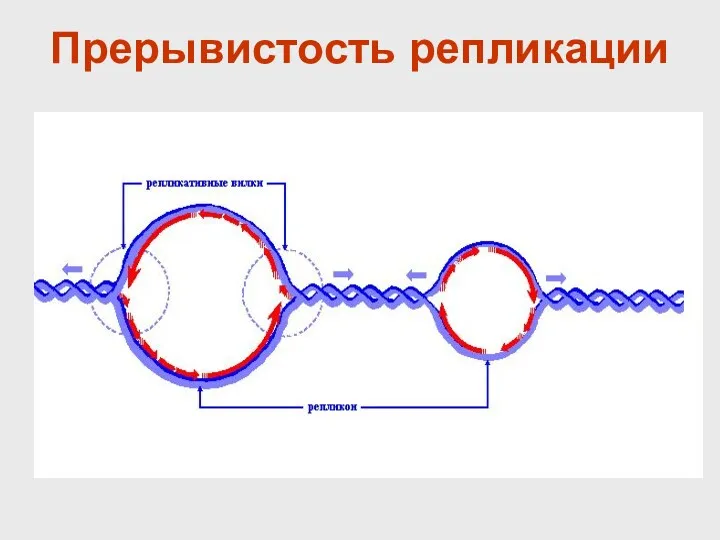
Прерывистость репликации
Прерывистость репликации
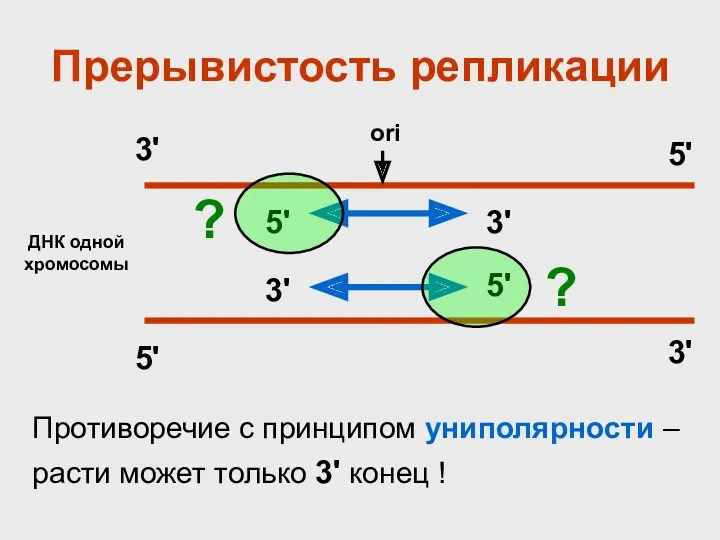
Прерывистость репликации
ДНК одной хромосомы
ori
3'
5'
3'
5'
5'
5'
3'
3'
Противоречие с принципом униполярности – расти может только
Прерывистость репликации
ДНК одной хромосомы
ori
3'
5'
3'
5'
5'
5'
3'
3'
Противоречие с принципом униполярности – расти может только
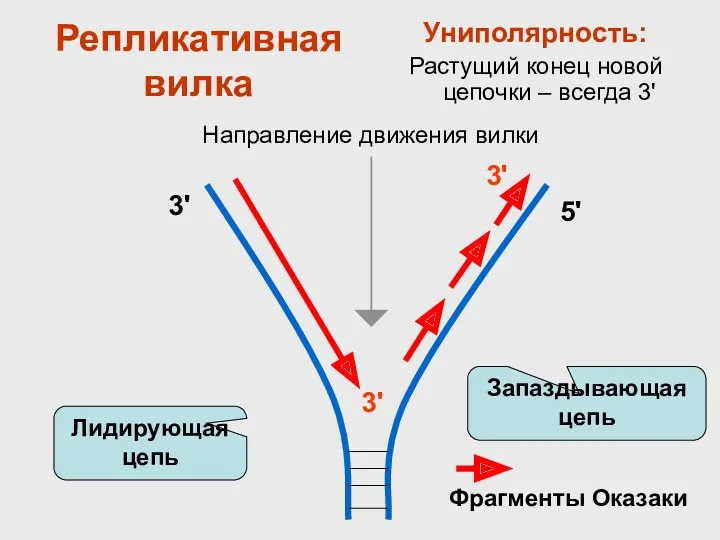
Репликативная вилка
Униполярность:
Растущий конец новой цепочки – всегда 3'
3'
5'
3'
3'
Запаздывающая цепь
Лидирующая цепь
Направление движения
Репликативная вилка
Униполярность:
Растущий конец новой цепочки – всегда 3'
3'
5'
3'
3'
Запаздывающая цепь
Лидирующая цепь
Направление движения

Молекулярные машины
Комплекс белков и ферментов, действующих согласованно
Реплисома
Рибосома
Сплайсосома
Молекулярные машины
Комплекс белков и ферментов, действующих согласованно
Реплисома
Рибосома
Сплайсосома
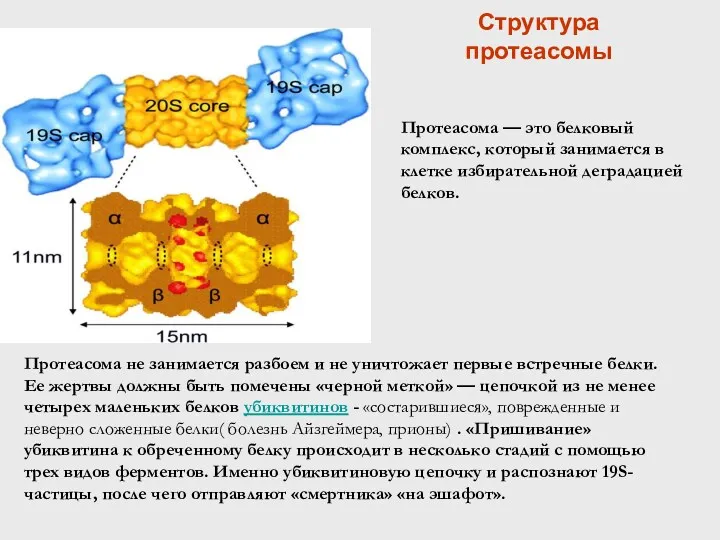
Структура протеасомы
Протеасома — это белковый комплекс, который занимается в клетке избирательной деградацией
Структура протеасомы
Протеасома — это белковый комплекс, который занимается в клетке избирательной деградацией

Молекулярная машина репликации
Молекулярная машина репликации
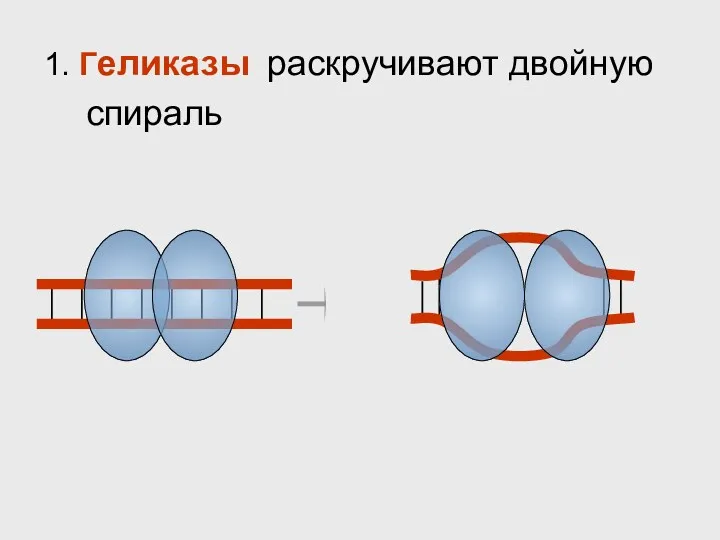
1. Геликазы раскручивают двойную спираль
1. Геликазы раскручивают двойную спираль
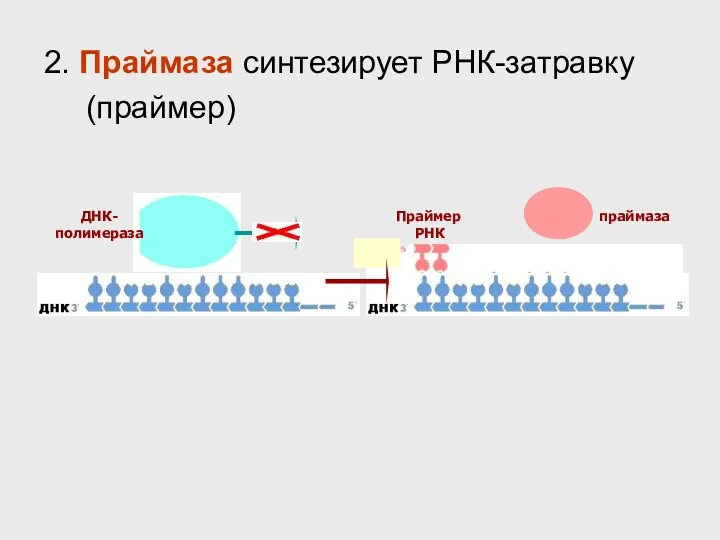
ДНК-
полимераза
праймаза
Праймер
РНК
2. Праймаза синтезирует РНК-затравку (праймер)
ДНК-
полимераза
праймаза
Праймер
РНК
2. Праймаза синтезирует РНК-затравку (праймер)
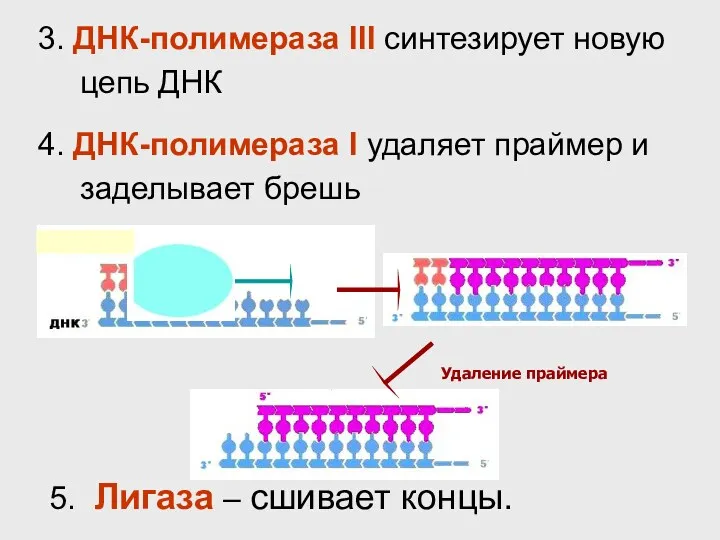
Удаление праймера
3. ДНК-полимераза III синтезирует новую цепь ДНК
4. ДНК-полимераза I удаляет
Удаление праймера
3. ДНК-полимераза III синтезирует новую цепь ДНК
4. ДНК-полимераза I удаляет
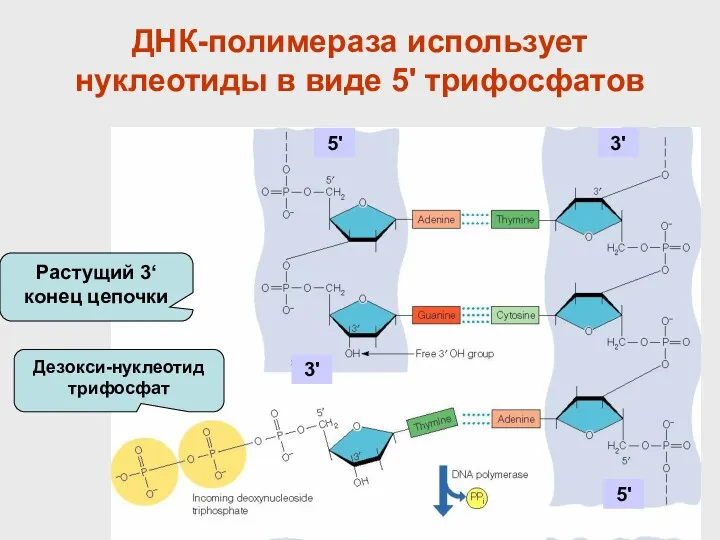
ДНК-полимераза использует нуклеотиды в виде 5' трифосфатов
Растущий 3‘ конец цепочки
Дезокси-нуклеотид трифосфат
5'
3'
5'
3'
ДНК-полимераза использует нуклеотиды в виде 5' трифосфатов
Растущий 3‘ конец цепочки
Дезокси-нуклеотид трифосфат
5'
3'
5'
3'
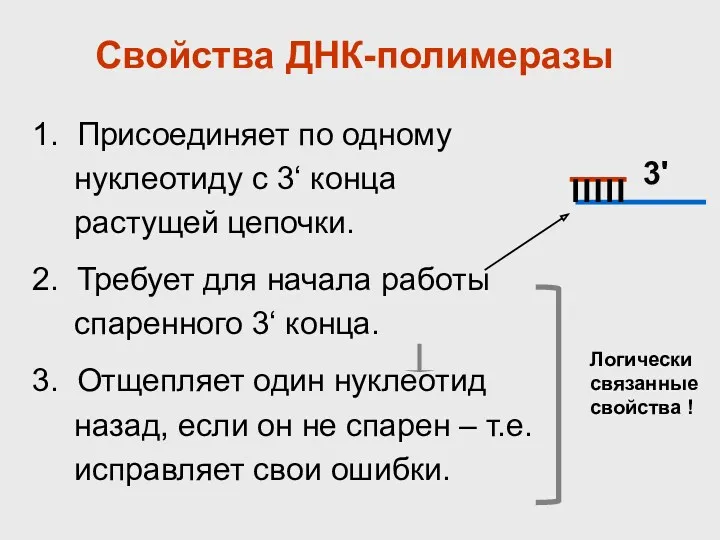
Свойства ДНК-полимеразы
1. Присоединяет по одному нуклеотиду с 3‘ конца растущей цепочки.
2.
Свойства ДНК-полимеразы
1. Присоединяет по одному нуклеотиду с 3‘ конца растущей цепочки.
2.
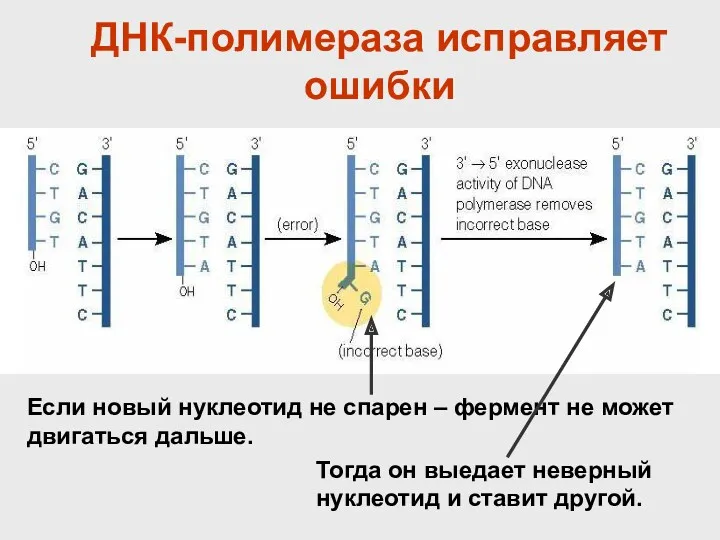
ДНК-полимераза исправляет ошибки
Если новый нуклеотид не спарен – фермент не может
ДНК-полимераза исправляет ошибки
Если новый нуклеотид не спарен – фермент не может

Скорость репликации ДНК
У прокариот – 1000 нуклеотидов /сек
У эукариот – 100
Скорость репликации ДНК
У прокариот – 1000 нуклеотидов /сек
У эукариот – 100

Выводы по репликации ДНК
В результате репликации каждая дочерняя клетка получает точную
Выводы по репликации ДНК
В результате репликации каждая дочерняя клетка получает точную

Мутации
и
системы репарации
Мутации
и
системы репарации

Частота ошибок ДНК-полимеразы ~1٠10– 9
– 1 нуклеотид на миллиард.
Мутации –
Частота ошибок ДНК-полимеразы ~1٠10– 9
– 1 нуклеотид на миллиард.
Мутации –

Спонтанный уровень (внутренние причины мутаций):
Ошибки репликации
Ошибки деления клеток
Перемещение
Спонтанный уровень (внутренние причины мутаций):
Ошибки репликации
Ошибки деления клеток
Перемещение
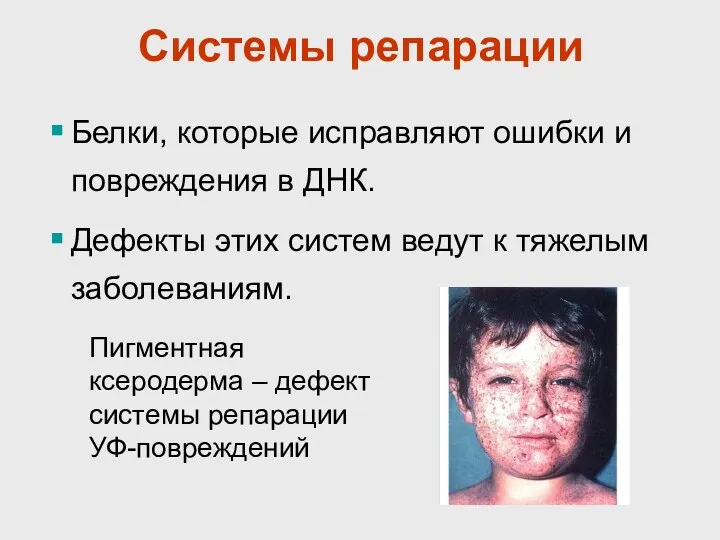
Системы репарации
Белки, которые исправляют ошибки и повреждения в ДНК.
Дефекты этих систем
Системы репарации
Белки, которые исправляют ошибки и повреждения в ДНК.
Дефекты этих систем
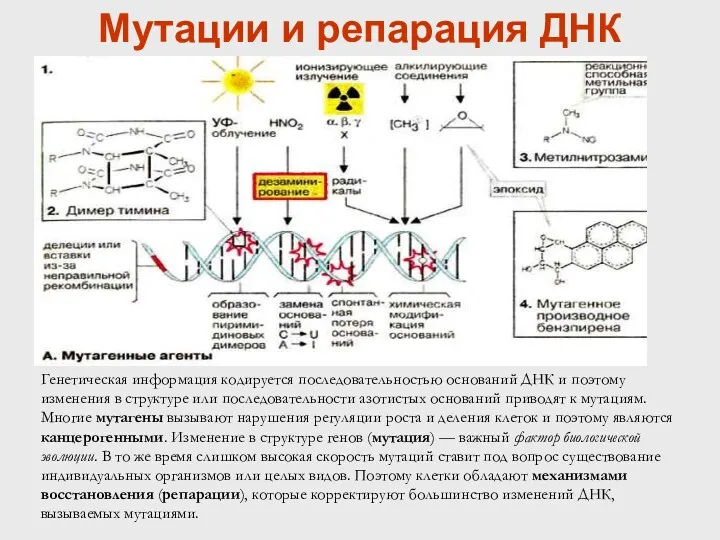
Мутации и репарация ДНК
Генетическая информация кодируется последовательностью оснований ДНК и поэтому
Мутации и репарация ДНК
Генетическая информация кодируется последовательностью оснований ДНК и поэтому
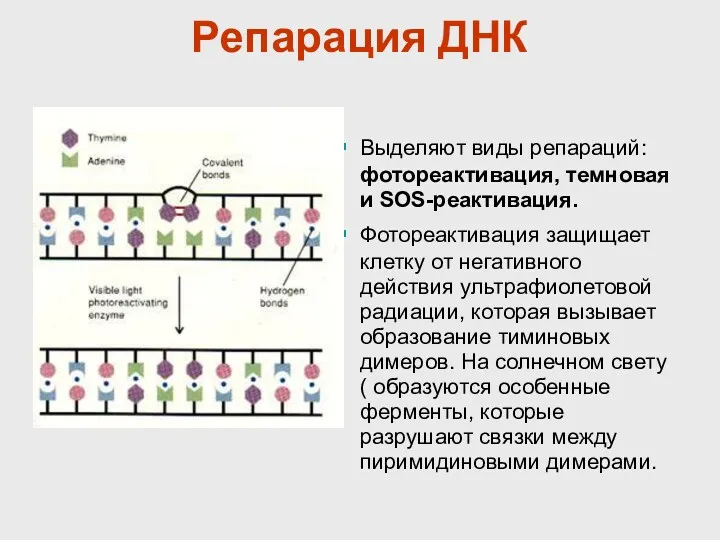
Репарация ДНК
Выделяют виды репараций: фотореактивация, темновая и SOS-реактивация.
Фотореактивация защищает клетку
Репарация ДНК
Выделяют виды репараций: фотореактивация, темновая и SOS-реактивация.
Фотореактивация защищает клетку
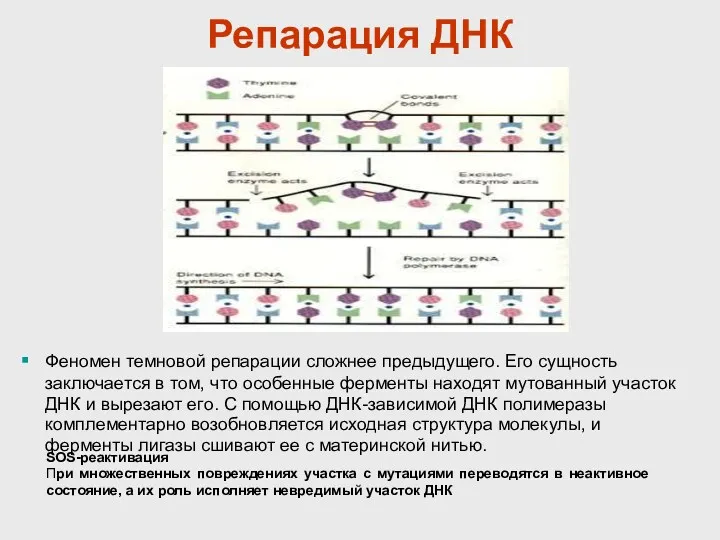
Репарация ДНК
Феномен темновой репарации сложнее предыдущего. Его сущность заключается в том,
Репарация ДНК
Феномен темновой репарации сложнее предыдущего. Его сущность заключается в том,

Основные понятия по теме «Репликация»
Место репликации в клеточном цикле
Принципы репликации
Лидирующая и
Основные понятия по теме «Репликация»
Место репликации в клеточном цикле
Принципы репликации
Лидирующая и

Теломеры
и
теломераза
Дополнение к теме
Теломеры
и
теломераза
Дополнение к теме
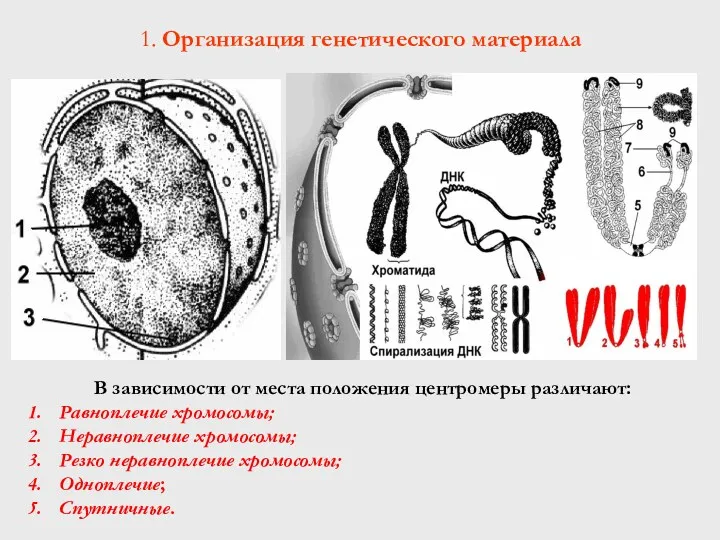
В зависимости от места положения центромеры различают:
Равноплечие хромосомы;
Неравноплечие хромосомы;
Резко неравноплечие
В зависимости от места положения центромеры различают:
Равноплечие хромосомы;
Неравноплечие хромосомы;
Резко неравноплечие
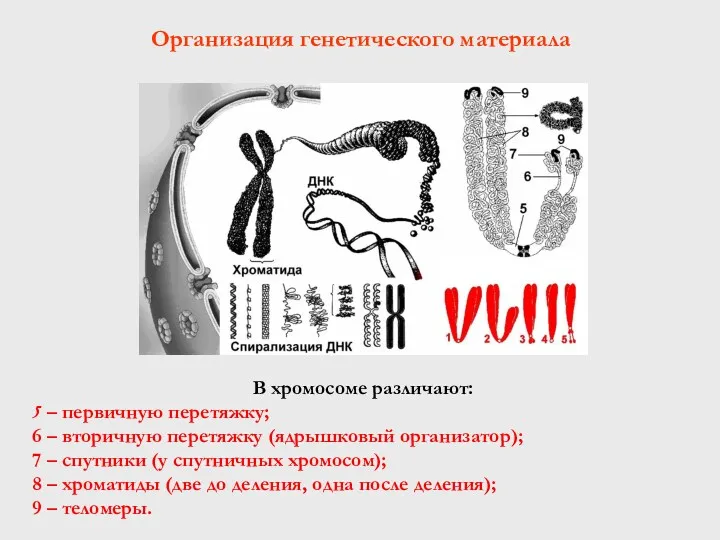
В хромосоме различают:
5 – первичную перетяжку;
6 – вторичную перетяжку (ядрышковый
В хромосоме различают:
5 – первичную перетяжку;
6 – вторичную перетяжку (ядрышковый

Проблема укорочения концов у линейных ДНК
Сформулирована – А.М. Оловников, 1971
При каждой
Проблема укорочения концов у линейных ДНК
Сформулирована – А.М. Оловников, 1971
При каждой
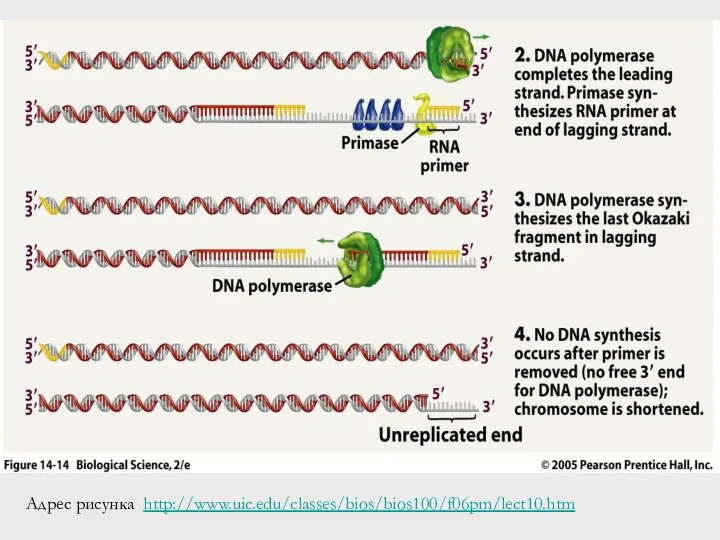
Адрес рисунка http://www.uic.edu/classes/bios/bios100/f06pm/lect10.htm
Адрес рисунка http://www.uic.edu/classes/bios/bios100/f06pm/lect10.htm

Гипотеза Оловникова
Укорочение концов – это внутренние часы, отмеряющие время жизни многоклеточного
Гипотеза Оловникова
Укорочение концов – это внутренние часы, отмеряющие время жизни многоклеточного

Но почему тогда клетки зародышевой линии делятся бесконечно?
Оловников: должен существовать механизм
Но почему тогда клетки зародышевой линии делятся бесконечно?
Оловников: должен существовать механизм

Теломераза
фермент, надстраивающий концы хромосом, содержит РНК.
удлинение происходит путем
обратной транскрипции:
РНК →
Теломераза
фермент, надстраивающий концы хромосом, содержит РНК.
удлинение происходит путем
обратной транскрипции:
РНК →

 Память и обучение
Память и обучение Строение тела человека. (2 класс)
Строение тела человека. (2 класс) Класс Насекомые
Класс Насекомые Влияние электромагнитного поля на биологические объекты
Влияние электромагнитного поля на биологические объекты Потоки вещества и энергии, динамика экосистем
Потоки вещества и энергии, динамика экосистем Биоценоз и биогеоценоз
Биоценоз и биогеоценоз Строение цветка
Строение цветка Дополнительная презентация Что такое фитодизайн?к элективному курсу Основы фитодизайна, 9 класс
Дополнительная презентация Что такое фитодизайн?к элективному курсу Основы фитодизайна, 9 класс Ара шаруашылығындағы азық қоры
Ара шаруашылығындағы азық қоры Характеристика вирусов и прокариот
Характеристика вирусов и прокариот Отдел Покрытосеменные: Ч.1
Отдел Покрытосеменные: Ч.1 Отличие черного хлеба от белого
Отличие черного хлеба от белого методическая разработка урока на тему: клеточное строение листа для 6 класса
методическая разработка урока на тему: клеточное строение листа для 6 класса Ағзаларды клондау
Ағзаларды клондау Различные виды растений мира
Различные виды растений мира Познавательная игра для 6-х классов Интересные зеленые
Познавательная игра для 6-х классов Интересные зеленые Пищеварение в ротовой полости
Пищеварение в ротовой полости Гиацинт (лат. Hyacínthus)
Гиацинт (лат. Hyacínthus) Красота и гармония в природе зимой. 6 класс
Красота и гармония в природе зимой. 6 класс Молекулярно-генетический уровень организации живого
Молекулярно-генетический уровень организации живого Бады - лечим или калечим
Бады - лечим или калечим Кто такие рыбы
Кто такие рыбы Гусеобразные Мелеузовского района: земляные и речные утки. Часть 2
Гусеобразные Мелеузовского района: земляные и речные утки. Часть 2 Введение в биохимию. Структура и функции белков
Введение в биохимию. Структура и функции белков Endargered animals
Endargered animals Надцарство доядерные
Надцарство доядерные Биологическое действие ионизирующего излучения
Биологическое действие ионизирующего излучения Земноводные. Лягушка квакша
Земноводные. Лягушка квакша