- Генетическая инженерия: молекулярные основы

Содержание
- 2. Предмет, задачи, история возникновения генетической инженерии 1
- 3. Генетическая инженерия - конструирование в искусственных условиях функционально активных генетических структур – рекомбинантных ДНК. Рекомбинантные ДНК
- 4. Научное познание живой материи: возможность получения генетического материала (количества вещества генов) для последующего изучения геномов живых
- 5. От открытия ДНК – к ДНК-технологиям 1871 г. – открытие ДНК Ф. Мишер выделил из изолированных
- 6. 1928 г. – Ф. Гриффитс, открытие генетической трансформации у пневмококков. Фредерик Гриффитс (1881 – 1941) английский
- 7. 1944 г., О. Эвери – доказательсво роли ДНК как носителя наследственной информации. Освальд Эвери, американский биолог
- 8. 1953 г. – Ф. Крик, Д. Уотсон, М. Уилкинс – открытие химической структуры ДНК
- 9. 1960 - гг. – открытие рестриктаз – ферментов, разрезающих молекулу ДНК («молекулярные ножницы») Вернер Арбер швейцарский
- 10. Создание первой рекомбинантной молекулы ДНК: вирус SV40, бактериофаг λ и E. coli. Пол Берг 1972 -73
- 11. Строение и свойства ДНК 2 «Нуклеиновые кислоты в основе своей просты. Они лежат у истока самых
- 12. ДНК (дезоксирибонуклеиновая кислота) - биополимер, состоящий из мономерных молекул - нуклеотидов. В состав ДНК входят 4
- 13. Состав нуклеотида ( тимидин ) 1 4 3 2 5
- 14. Как образуется полимерная молекула? Молекула ДНК имеет несколько уровней структурной организации.
- 15. Первичная структура ДНК образуется за счет возникновения 3’,5’-фосфодиэфирной связи между 3’-гидроксилом и 5’-фосфатом соседних нуклеотидов О
- 16. А Т Г Т А А Т Ц Вторичная структура ДНК образуется за счет водородных связей
- 17. 5’ 3’ --А Т Г А Т А Г Т А Ц Ц А А Ц
- 18. Двойная спираль ДНК – две антипараллельные цепи закручены вокруг продольной оси.
- 19. Размер молекулы ДНК измеряется числом пар нуклеотидов, за единицу измерения взята килобаза - тысяча пар нуклеотидов,
- 20. Свойства ДНК
- 21. Денатурация - плавление ДНК, разрушение вторичной структуры молекулы ДНК вследствие разрыва водородных связей T=60…900C
- 22. Ренатурация - отжиг ДНК, восстановление вторичной структуры вследствие образования водородных связей между комплементарными азотистыми основаниями
- 23. Ферменты генетической инженерии 3
- 24. 1. Рестриктазы (эндонуклеазы рестрикции) ферменты, расщепляющие фосфодиэфирную связь и разрезающие молекулу ДНК на отдельные фрагменты.
- 25. Рестриктазы открыты и выделены В. Арбером в 1962 г. в опытах по инфицированию бактериофагом λ штаммов
- 26. Номенклатура рестриктаз: при обозначении рестриктаз используют родо-видовое название микроорганизма из которого выделен фермент: co scherichia E
- 27. Субстратная специфичность - способны распознавать определенную последовательность нуклеотидов на молекуле ДНК, в которой происходит разрыв –
- 28. В генетической инженрии используются рестриктазы 2-го класса – узнают последовательности, одинаково читаемые по обеим цепям –
- 29. Характер расщепления сайта рестрикции По оси симметрии – с образованием «тупых» концов -- 5’ АЦЦТЦТТАТТАЦ --
- 30. Ступенчато – с образованием «липких» концов АЦГТААТТАТАЦЦЦТ 3’ -- ТТААТАТ ГГГА 5’ -- -- 5’ АЦЦТЦТТАТТ
- 31. Некоторые рестриктазы 2 класса
- 32. Получение рестриктных фрагментов ААТТЦАТЦГАТЦАТАЦЦЦАТГАААТАЦЦАТГАТЦААТГАА ГТАГЦТАГ ТАТГ Г ГТАЦТТТАТ ГГ ТАЦТАГТТАЦТТ ЦАТТГТ ТТТ Г Г ГТААЦААААЦЦ ТТАА
- 33. Разделение фрагментов электрофорезом Eco Bme Eco + Bme Источник питания Гелевая пластина Ванна с электролитом Лунки
- 34. 2. Экзонуклеазы – ферменты, гидролизующие фосфодиэфирную связь с 5’ или 3’ конца. Отщепляют от молекулы ДНК
- 35. 3. Лигазы – ферменты, катализирующие образование фосфодиэфирной связи и способствующие сшиванию разных фрагментов ДНК в единую
- 36. 4. ДНК-полимераза – фермент, катализирующий синтез полимерной молекулы ДНК на ДНК- матрице, способствующий репликации (удвоению) ДНК.
- 37. Для реакции полимеризации необходимы: ДНК – матрица Праймер (ДНК-затравка), комплементарный 3’-концу матрицы Свободные нуклеотиды Фермент ДНК-полимераза
- 38. 5. Обратная транскриптаза (ревертаза) – фермент, катализируюший синтез ДНК на РНК-матрице. ДНК, синтезированная таким образом называется
- 39. 6. Терминальная трансфераза – фермент, присоединяющий к 3’-концу ДНК отдельные нуклеотиды. ГГГГГ Гомополимерный липкий конец ГГГГГ
- 40. Методы создания рекомбинантных ДНК 4
- 41. 1. Коннекторный метод (от англ. to connect – соединять) – основан на использовании терминальной трансферазы, синтезирующей
- 42. АА АААААА ААА ТТТТ ТТТТТ Схема коннекторного метода ТТ
- 43. 2. Рестриктазно-лигазный метод – основан на использовании рестриктаз, образующих у разрезаемых фрагментов ДНК липкие концы. вырезание
- 44. Рис. 9. Схема получения рекомбинантной плазмиды рестриктазно- лигазным методом (по Щелкунову С.Н., 1995). Eco Схема рестриктазно-лигазного
- 45. Если невозможно выделить разные фрагменты ДНК одной и той же рестриктазой, используется линкер. Линкеры (от англ.
- 46. Рис. 9. Схема получения рекомбинантной плазмиды рестриктазно- лигазным методом (по Щелкунову С.Н., 1995). Eco Схема использования
- 47. Варианты конфигурации рекомбинантных ДНК Линейная Кольцевая Восьмерки и петли
- 49. Скачать презентацию

Предмет, задачи,
история возникновения
генетической инженерии
1
Предмет, задачи,
история возникновения
генетической инженерии
1

Генетическая инженерия - конструирование в искусственных условиях функционально активных генетических структур
Генетическая инженерия - конструирование в искусственных условиях функционально активных генетических структур

Научное познание живой материи:
возможность получения генетического материала (количества вещества генов)
Научное познание живой материи:
возможность получения генетического материала (количества вещества генов)

От открытия ДНК – к ДНК-технологиям
1871 г. – открытие ДНК
Ф.
От открытия ДНК – к ДНК-технологиям
1871 г. – открытие ДНК
Ф.
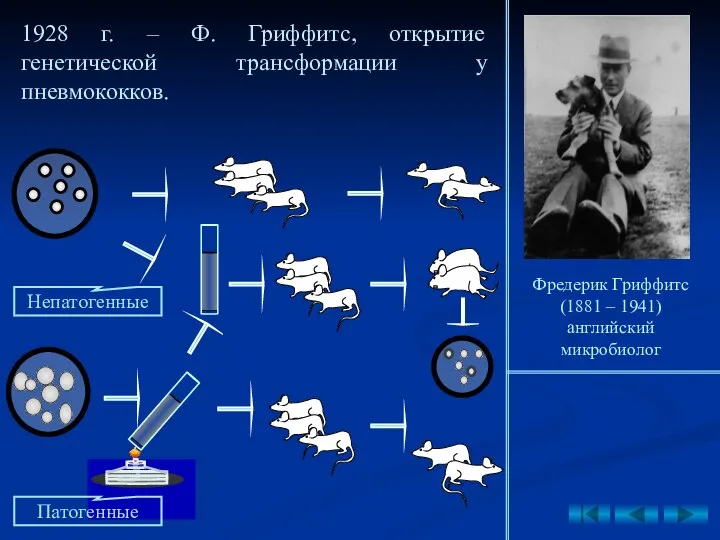
1928 г. – Ф. Гриффитс, открытие генетической трансформации у пневмококков.
Фредерик
1928 г. – Ф. Гриффитс, открытие генетической трансформации у пневмококков.
Фредерик
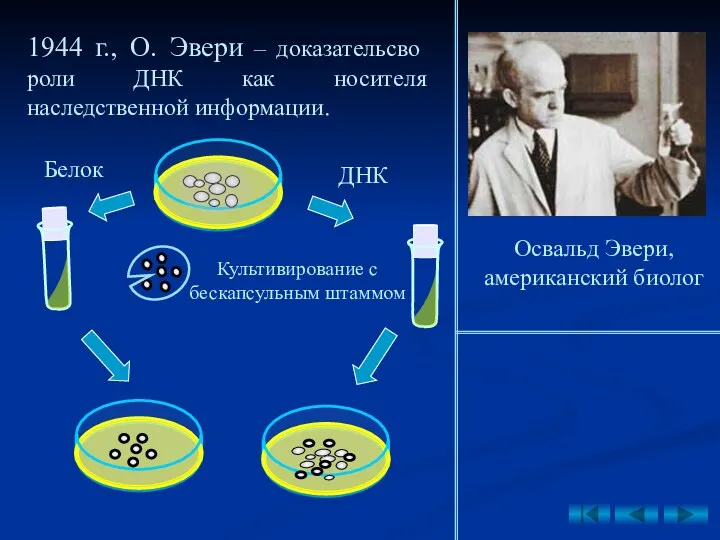
1944 г., О. Эвери – доказательсво роли ДНК как носителя наследственной
1944 г., О. Эвери – доказательсво роли ДНК как носителя наследственной

1953 г. – Ф. Крик, Д. Уотсон, М. Уилкинс – открытие
1953 г. – Ф. Крик, Д. Уотсон, М. Уилкинс – открытие

1960 - гг. – открытие рестриктаз – ферментов, разрезающих молекулу ДНК
1960 - гг. – открытие рестриктаз – ферментов, разрезающих молекулу ДНК

Создание первой рекомбинантной молекулы ДНК: вирус SV40, бактериофаг λ и E.
Создание первой рекомбинантной молекулы ДНК: вирус SV40, бактериофаг λ и E.

Строение и свойства ДНК
2
«Нуклеиновые кислоты в основе своей просты.
Они лежат
Строение и свойства ДНК
2
«Нуклеиновые кислоты в основе своей просты.
Они лежат
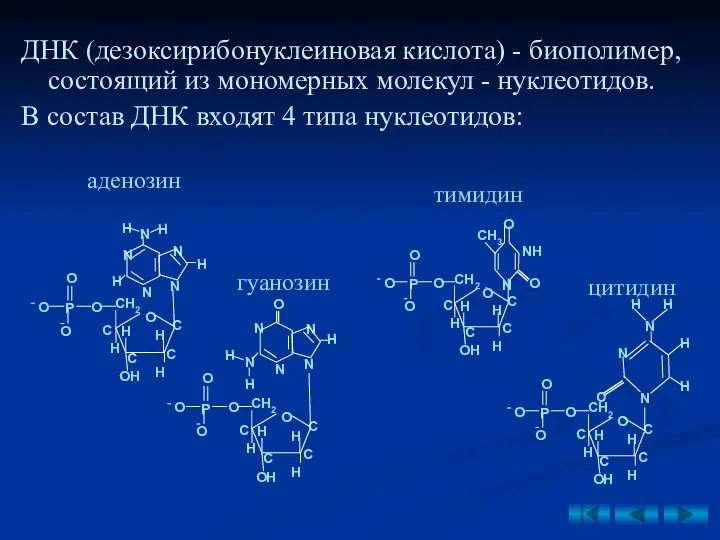
ДНК (дезоксирибонуклеиновая кислота) - биополимер, состоящий из мономерных молекул - нуклеотидов.
В
ДНК (дезоксирибонуклеиновая кислота) - биополимер, состоящий из мономерных молекул - нуклеотидов.
В
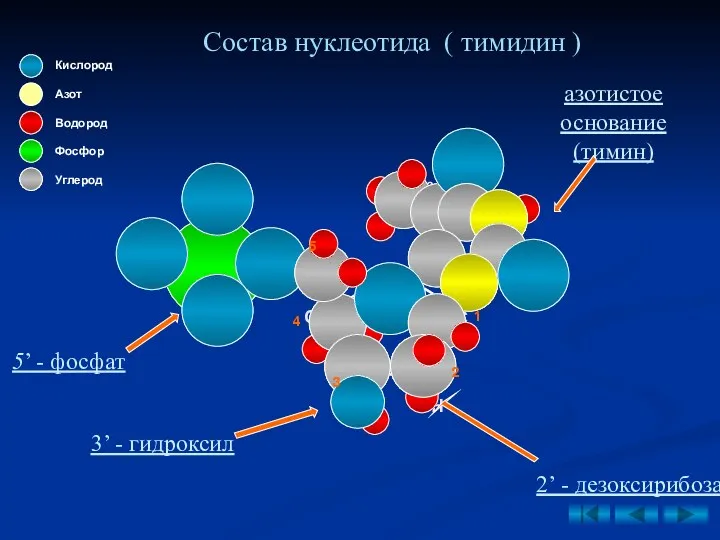
Состав нуклеотида ( тимидин )
1
4
3
2
5
Состав нуклеотида ( тимидин )
1
4
3
2
5

Как образуется полимерная молекула?
Молекула ДНК имеет несколько уровней структурной организации.
Как образуется полимерная молекула?
Молекула ДНК имеет несколько уровней структурной организации.
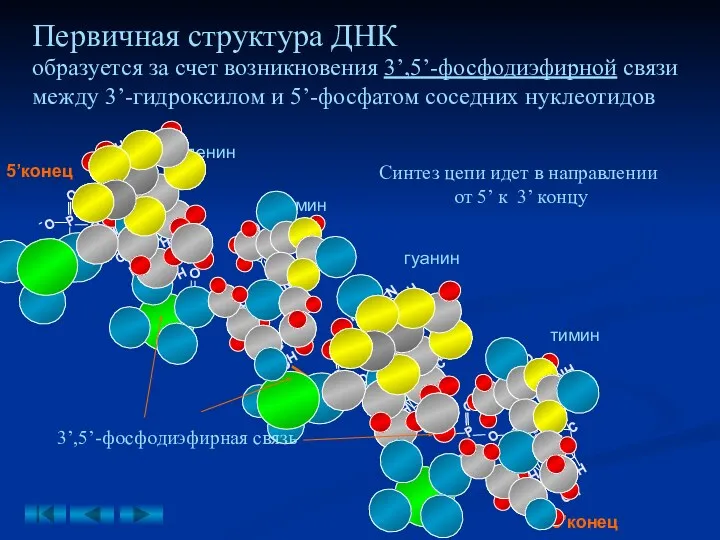
Первичная структура ДНК
образуется за счет возникновения 3’,5’-фосфодиэфирной связи между 3’-гидроксилом
Первичная структура ДНК
образуется за счет возникновения 3’,5’-фосфодиэфирной связи между 3’-гидроксилом
А
Т
Г
Т
А
А
Т
Ц
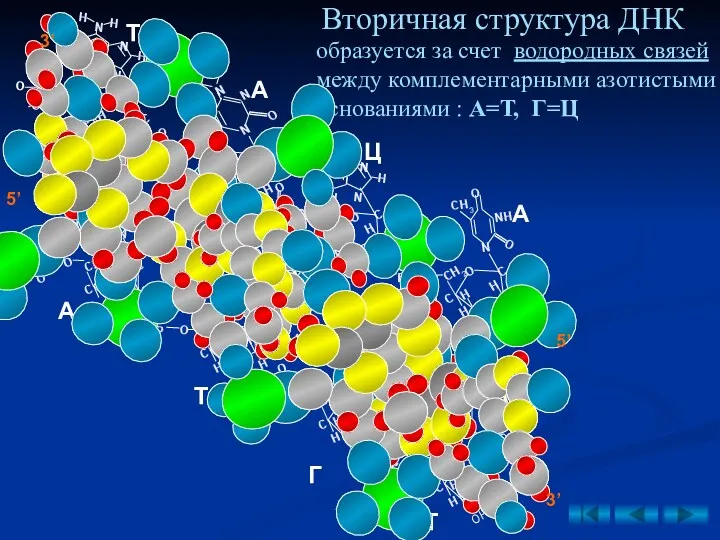
Вторичная структура ДНК
образуется за
А
Т
Г
Т
А
А
Т
Ц
Вторичная структура ДНК
образуется за

5’ 3’
--А Т Г А Т А Г Т А
5’ 3’
--А Т Г А Т А Г Т А
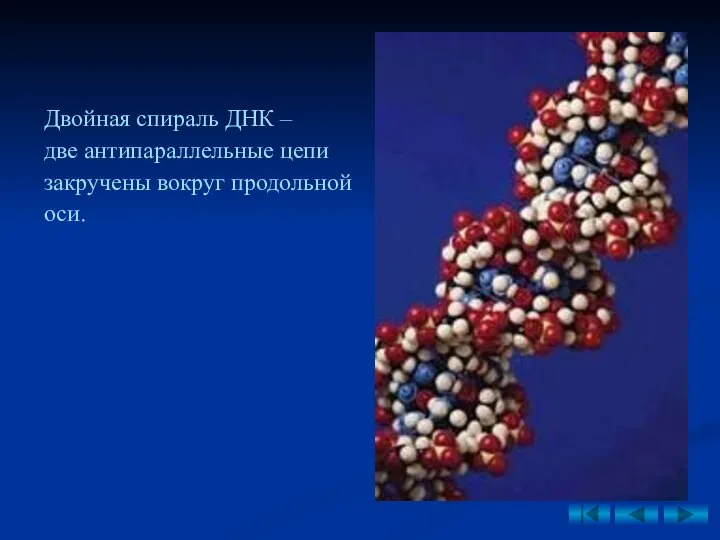
Двойная спираль ДНК –
две антипараллельные цепи закручены вокруг продольной оси.
Двойная спираль ДНК –
две антипараллельные цепи закручены вокруг продольной оси.

Размер молекулы ДНК измеряется числом пар нуклеотидов, за единицу измерения взята
Размер молекулы ДНК измеряется числом пар нуклеотидов, за единицу измерения взята

Свойства ДНК
Свойства ДНК
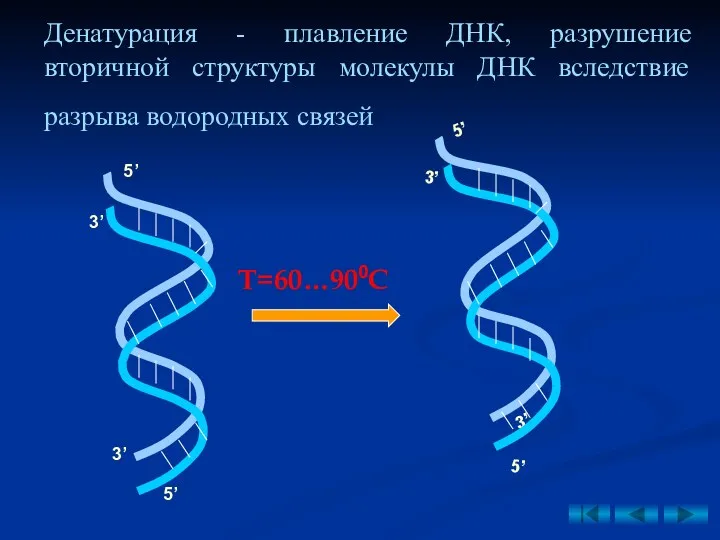
Денатурация - плавление ДНК, разрушение вторичной структуры молекулы ДНК вследствие разрыва
Денатурация - плавление ДНК, разрушение вторичной структуры молекулы ДНК вследствие разрыва

Ренатурация - отжиг ДНК, восстановление вторичной структуры вследствие образования водородных связей
Ренатурация - отжиг ДНК, восстановление вторичной структуры вследствие образования водородных связей

Ферменты
генетической инженерии
3
Ферменты
генетической инженерии
3

1. Рестриктазы (эндонуклеазы рестрикции)
ферменты, расщепляющие фосфодиэфирную связь и
разрезающие молекулу
1. Рестриктазы (эндонуклеазы рестрикции)
ферменты, расщепляющие фосфодиэфирную связь и
разрезающие молекулу
Рестриктазы открыты и выделены В. Арбером в 1962 г. в опытах
Рестриктазы открыты и выделены В. Арбером в 1962 г. в опытах

Номенклатура рестриктаз:
при обозначении рестриктаз используют родо-видовое название микроорганизма из которого выделен
Номенклатура рестриктаз:
при обозначении рестриктаз используют родо-видовое название микроорганизма из которого выделен

Субстратная специфичность - способны распознавать определенную последовательность нуклеотидов на молекуле ДНК,
Субстратная специфичность - способны распознавать определенную последовательность нуклеотидов на молекуле ДНК,

В генетической инженрии используются рестриктазы 2-го класса – узнают последовательности, одинаково
В генетической инженрии используются рестриктазы 2-го класса – узнают последовательности, одинаково

Характер расщепления сайта рестрикции
По оси симметрии – с образованием «тупых» концов
--
Характер расщепления сайта рестрикции
По оси симметрии – с образованием «тупых» концов
--

Ступенчато – с образованием «липких» концов
АЦГТААТТАТАЦЦЦТ 3’ --
ТТААТАТ ГГГА 5’
Ступенчато – с образованием «липких» концов
АЦГТААТТАТАЦЦЦТ 3’ --
ТТААТАТ ГГГА 5’

Некоторые рестриктазы 2 класса
Некоторые рестриктазы 2 класса

Получение рестриктных фрагментов
ААТТЦАТЦГАТЦАТАЦЦЦАТГАААТАЦЦАТГАТЦААТГАА
ГТАГЦТАГ ТАТГ Г ГТАЦТТТАТ ГГ ТАЦТАГТТАЦТТ
ЦАТТГТ ТТТ Г
Получение рестриктных фрагментов
ААТТЦАТЦГАТЦАТАЦЦЦАТГАААТАЦЦАТГАТЦААТГАА
ГТАГЦТАГ ТАТГ Г ГТАЦТТТАТ ГГ ТАЦТАГТТАЦТТ
ЦАТТГТ ТТТ Г
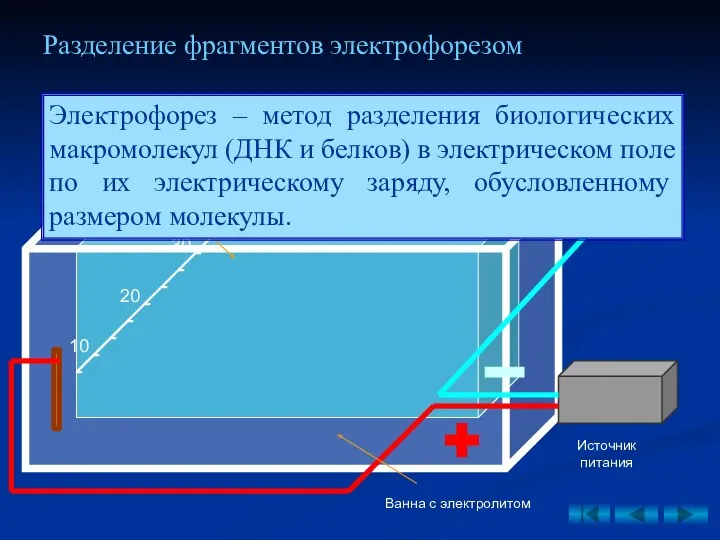
Разделение фрагментов электрофорезом
Eco
Bme
Eco + Bme
Источник питания
Гелевая пластина
Ванна с электролитом
Лунки с образцами
Разделение фрагментов электрофорезом
Eco
Bme
Eco + Bme
Источник питания
Гелевая пластина
Ванна с электролитом
Лунки с образцами
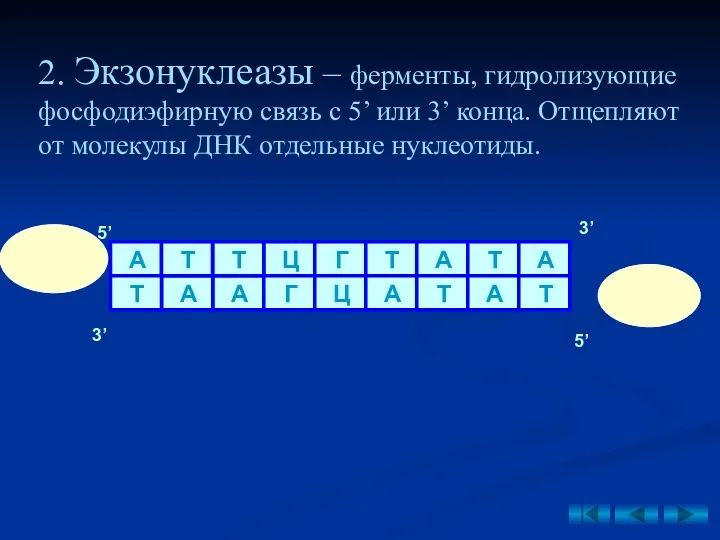
2. Экзонуклеазы – ферменты, гидролизующие фосфодиэфирную связь с 5’ или 3’
2. Экзонуклеазы – ферменты, гидролизующие фосфодиэфирную связь с 5’ или 3’

3. Лигазы – ферменты, катализирующие образование фосфодиэфирной связи и способствующие сшиванию
3. Лигазы – ферменты, катализирующие образование фосфодиэфирной связи и способствующие сшиванию

4. ДНК-полимераза – фермент,
катализирующий синтез полимерной молекулы ДНК
на ДНК-
4. ДНК-полимераза – фермент,
катализирующий синтез полимерной молекулы ДНК
на ДНК-

Для реакции полимеризации необходимы:
ДНК – матрица
Праймер (ДНК-затравка), комплементарный 3’-концу матрицы
Свободные нуклеотиды
Для реакции полимеризации необходимы:
ДНК – матрица
Праймер (ДНК-затравка), комплементарный 3’-концу матрицы
Свободные нуклеотиды
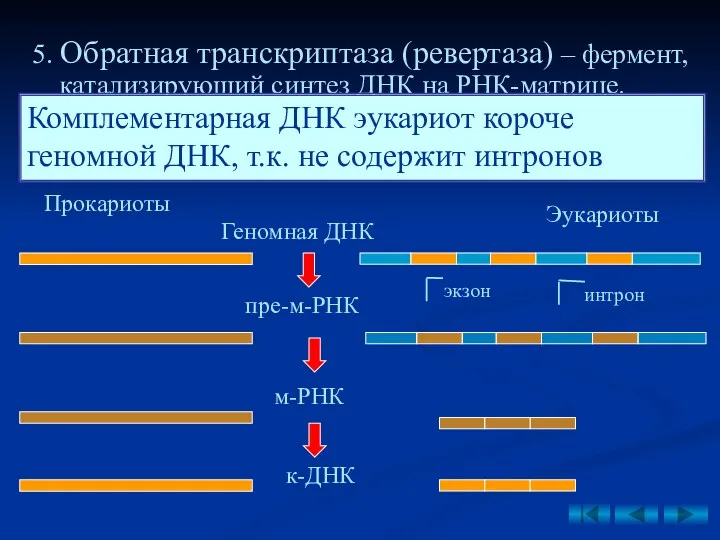
5. Обратная транскриптаза (ревертаза) – фермент, катализируюший синтез ДНК на РНК-матрице.
5. Обратная транскриптаза (ревертаза) – фермент, катализируюший синтез ДНК на РНК-матрице.
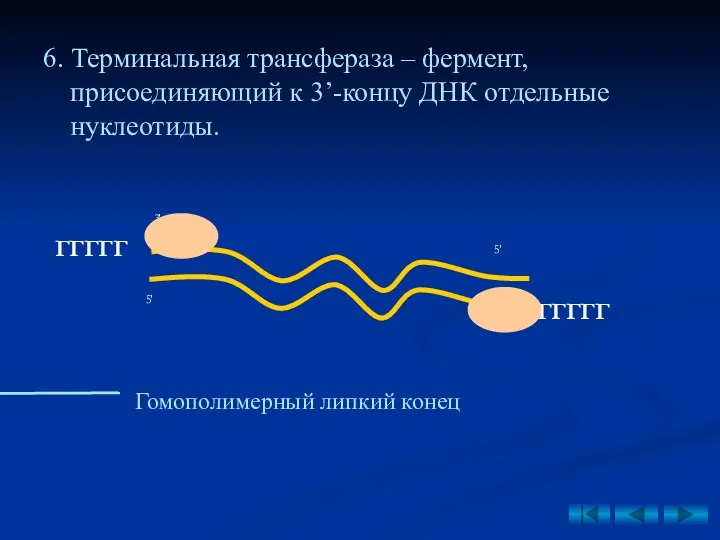
6. Терминальная трансфераза – фермент, присоединяющий к 3’-концу ДНК отдельные нуклеотиды.
ГГГГГ
Гомополимерный
6. Терминальная трансфераза – фермент, присоединяющий к 3’-концу ДНК отдельные нуклеотиды.
ГГГГГ
Гомополимерный

Методы создания рекомбинантных ДНК
4
Методы создания рекомбинантных ДНК
4

1. Коннекторный метод (от англ. to connect – соединять) – основан
1. Коннекторный метод (от англ. to connect – соединять) – основан
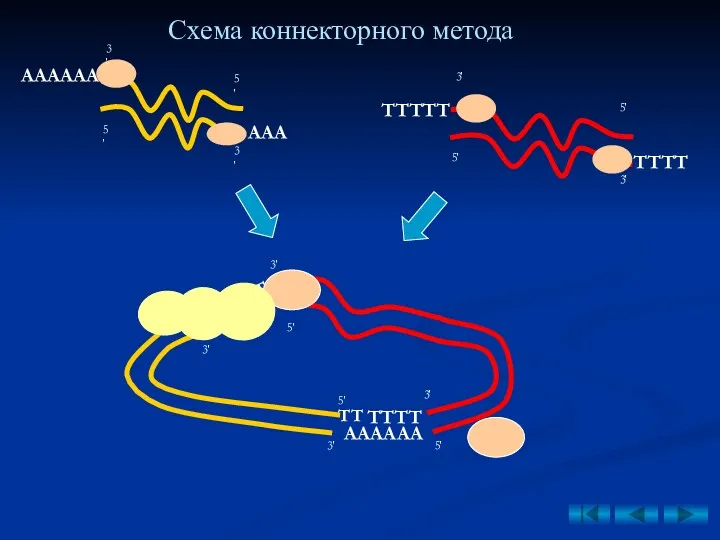
АА
АААААА
ААА
ТТТТ
ТТТТТ
Схема коннекторного метода
ТТ
АА
АААААА
ААА
ТТТТ
ТТТТТ
Схема коннекторного метода
ТТ

2. Рестриктазно-лигазный метод – основан на использовании рестриктаз, образующих у разрезаемых
2. Рестриктазно-лигазный метод – основан на использовании рестриктаз, образующих у разрезаемых
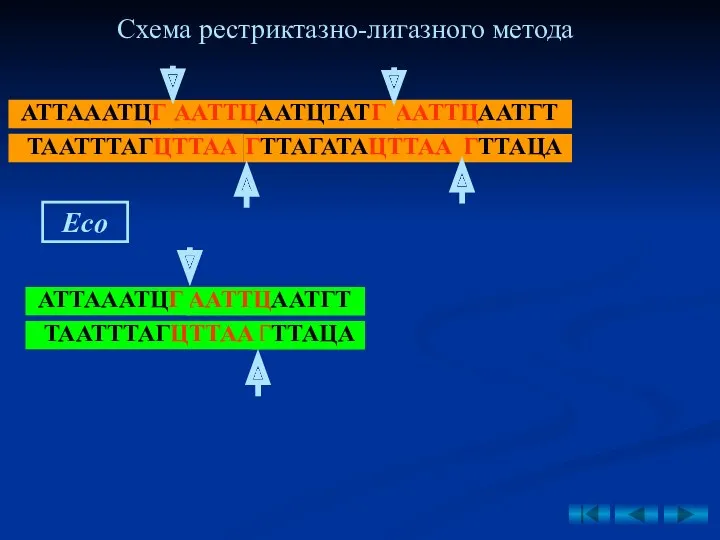
Рис. 9. Схема получения рекомбинантной плазмиды рестриктазно-
лигазным методом (по Щелкунову
Рис. 9. Схема получения рекомбинантной плазмиды рестриктазно-
лигазным методом (по Щелкунову

Если невозможно выделить разные фрагменты ДНК одной и той же рестриктазой,
Если невозможно выделить разные фрагменты ДНК одной и той же рестриктазой,
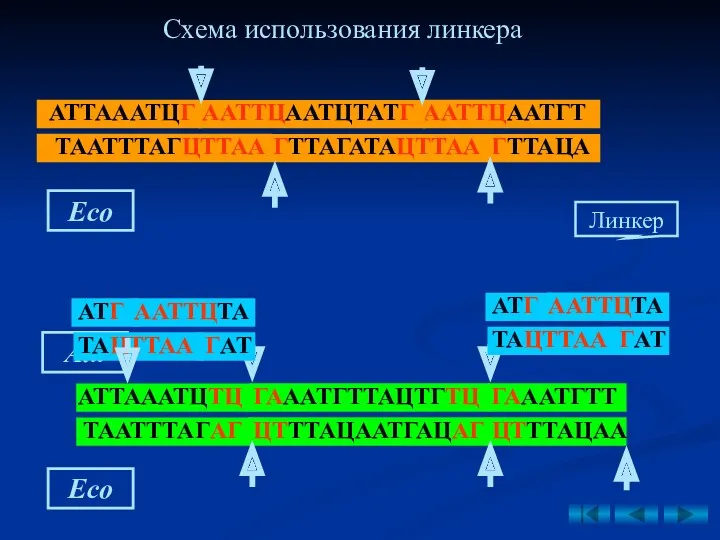
Рис. 9. Схема получения рекомбинантной плазмиды рестриктазно-
лигазным методом (по Щелкунову
Рис. 9. Схема получения рекомбинантной плазмиды рестриктазно-
лигазным методом (по Щелкунову
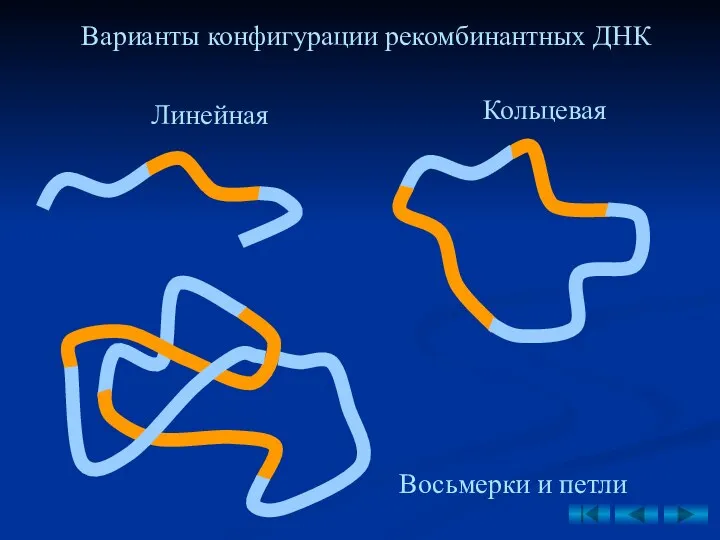
Варианты конфигурации рекомбинантных ДНК
Линейная
Кольцевая
Восьмерки и петли
Варианты конфигурации рекомбинантных ДНК
Линейная
Кольцевая
Восьмерки и петли
 Биология и распространение сурков
Биология и распространение сурков Селекция микроорганизмов. Биотехнология
Селекция микроорганизмов. Биотехнология Презентация по биологии Внутренняя среда организма. Кровь
Презентация по биологии Внутренняя среда организма. Кровь Учение об иммунитете
Учение об иммунитете Решение задач по генетике
Решение задач по генетике Основы систематики микроорганизмов
Основы систематики микроорганизмов Эндемики Австралии
Эндемики Австралии Рецепция (қабылдағыштар рөлі). Жүйке талшықтары, тұрлері, қозуды өткізу механизмі. Парабиоз
Рецепция (қабылдағыштар рөлі). Жүйке талшықтары, тұрлері, қозуды өткізу механизмі. Парабиоз слайды к уроку Курочка-ряба
слайды к уроку Курочка-ряба Процесс кровообращения. Вены большого круга кровообращения
Процесс кровообращения. Вены большого круга кровообращения Белки II. Уровни организации белков
Белки II. Уровни организации белков Corn
Corn Размножение и индивидуальное развитие организмов. Бесполое размножение
Размножение и индивидуальное развитие организмов. Бесполое размножение Презентация, Обобщающий урок по теме скелет.
Презентация, Обобщающий урок по теме скелет. Внешнее строение листа. Многообразие листьев
Внешнее строение листа. Многообразие листьев Исследовательский проект на тему: Мандариновое дерево
Исследовательский проект на тему: Мандариновое дерево Получение и применение моноклональных антител
Получение и применение моноклональных антител Электронный задачник. Часть 1. Содержание (Моногибридное скрещивание)
Электронный задачник. Часть 1. Содержание (Моногибридное скрещивание) Условия жизни на Земле Среды жизни на Земле и экологические факторы 9 класс По УМК И.Н. Пономаревой.
Условия жизни на Земле Среды жизни на Земле и экологические факторы 9 класс По УМК И.Н. Пономаревой. Догляд за руками
Догляд за руками Где живут белые медведи (окружающий мир, 1 класс)
Где живут белые медведи (окружающий мир, 1 класс) Скелет и соединения конечностей. Кости черепа человека
Скелет и соединения конечностей. Кости черепа человека Пищевые добавки. Классификация пищевых добавок
Пищевые добавки. Классификация пищевых добавок Изменения в жизни домашних животных весной. Труд людей весной (человек и мир, 1 класс)
Изменения в жизни домашних животных весной. Труд людей весной (человек и мир, 1 класс) Отдел: Покрытосеменные (Angiospermae), класс Двудольные (Dicotyledones)
Отдел: Покрытосеменные (Angiospermae), класс Двудольные (Dicotyledones) Айран дайындау технологиясы
Айран дайындау технологиясы Митоз. Митоздық цикл
Митоз. Митоздық цикл Эпителиальная ткань. Соединительная ткань. Лекция 3
Эпителиальная ткань. Соединительная ткань. Лекция 3