она имеется, бывает в покоящемся состоянии в виде плотного темно-окрашенного тельца, называемого тельцем Барра (факультативный гетерохроматин). Число телец Барра всегда на единицу меньше числа наличных х-хромосом, т.е. в мужском организме их нет вовсе, у женщин (ХХ) - одно.
У человека Y-хромосома является генетически инертной, так как в ней очень мало генов. Однако влияние Y-хромосомы на детерминацию пола у человека очень сильное. Хромосомная структура мужчины 44A+XY и женщины 44A+XX такая же, как и у дрозофилы, однако у человека особь с кариотипом 44A+XО оказалась женщиной, а особь 44A+XXY мужчиной. В обоих случаях они проявляли дефекты развития, но все же пол определялся наличием или отсутствием У-хромосомы.

 Презентация для учащихся 8 класса Внутренняя среда организма. Кровь, её состав и значение
Презентация для учащихся 8 класса Внутренняя среда организма. Кровь, её состав и значение Презентация Класс Птицы
Презентация Класс Птицы Красная книга Донецка
Красная книга Донецка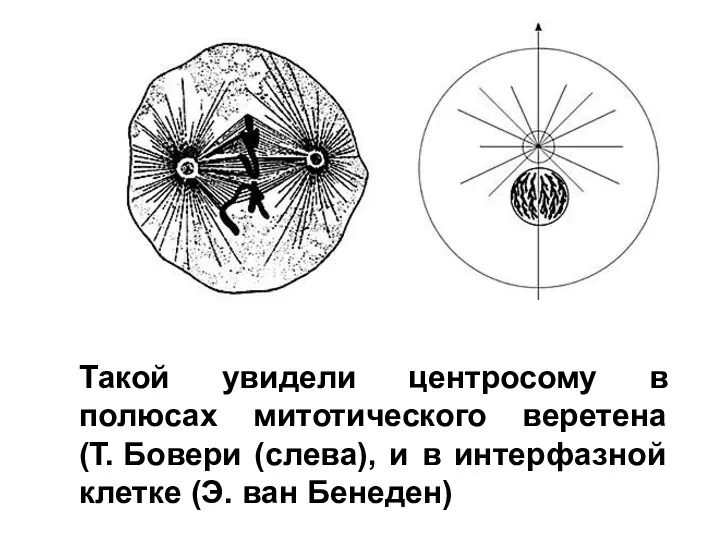 Клеточный центр (лекция 8)
Клеточный центр (лекция 8) Естественный отбор . Вид. Популяция
Естественный отбор . Вид. Популяция Екотоксикологічна характеристика мікотоксинів
Екотоксикологічна характеристика мікотоксинів Селекция животных
Селекция животных Органы цветкового растения. Лист
Органы цветкового растения. Лист Комнатные растения в интерьере квартиры
Комнатные растения в интерьере квартиры Урок по теме Ткани 6 класс
Урок по теме Ткани 6 класс Алкогольді мастықты талдаудың физико-химиялық әдістері
Алкогольді мастықты талдаудың физико-химиялық әдістері Галерея ученых - биологов
Галерея ученых - биологов Класс двудольные. Семейство сложноцветные (астровые)
Класс двудольные. Семейство сложноцветные (астровые) Влажные Экваториальные леса Африки
Влажные Экваториальные леса Африки Органические вещества клетки
Органические вещества клетки Размножение - воспроизведение себе подобных, обеспечивающее непрерывность и преемственность жизни
Размножение - воспроизведение себе подобных, обеспечивающее непрерывность и преемственность жизни Сущность жизни и свойства живого. Уровни организации живой материи
Сущность жизни и свойства живого. Уровни организации живой материи Анатомия и физиология человека
Анатомия и физиология человека Общая и медицинская генетика
Общая и медицинская генетика Идиоадаптации. 2 часть
Идиоадаптации. 2 часть Рослини - живі організми. Різноманітність рослин
Рослини - живі організми. Різноманітність рослин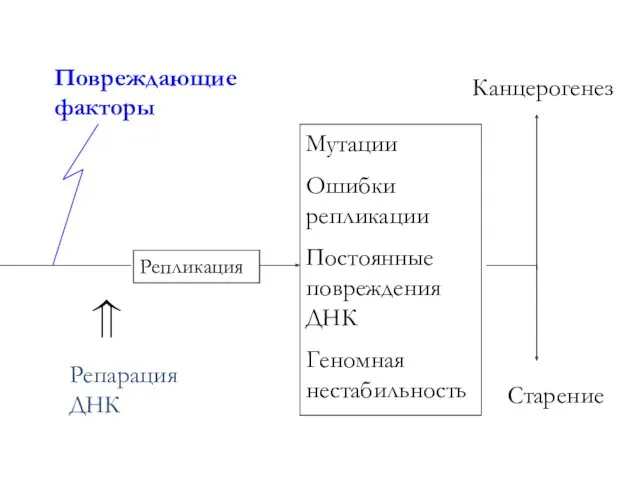 Репарация ДНК
Репарация ДНК Презентация Классификация споровых растений
Презентация Классификация споровых растений Особенности молекулярно-генетической и морфологической идентификации грибов подотдела Glomeromycotina
Особенности молекулярно-генетической и морфологической идентификации грибов подотдела Glomeromycotina Наука биология и условия жизни организмов
Наука биология и условия жизни организмов Пищеварение в желудке. Роль 12-ти перстной кишки в пищеварения. Пищеварительная функция печени
Пищеварение в желудке. Роль 12-ти перстной кишки в пищеварения. Пищеварительная функция печени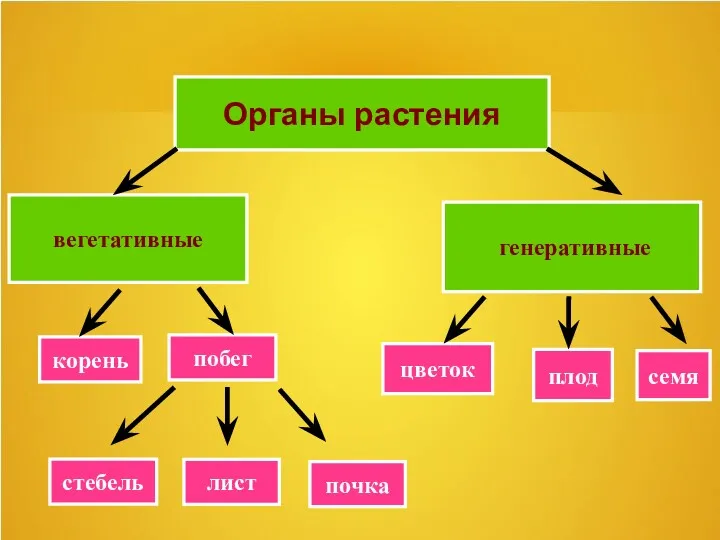 Цветок и его строение
Цветок и его строение анатомия и физиология звукового анализатора
анатомия и физиология звукового анализатора