- Генетические основы эволюции

Содержание
- 2. Изменчивость основное свойство живого в основе изменчивости - конвариантная редупликация
- 3. ДНК – основа изменчивости в процессе матричного копирования ДНК и РНК - ошибки последовательности расположения нуклеотидов
- 4. Ч. Дарвин вся изменчивость наследственная ненаследственную ненаследственных признаков нет все признаки и свойства организма наследственно обусловлены
- 5. Норма реакции В процессе размножения передаются не признаки, а гены гены определяют возможность развития будущих признаков
- 6. Норма реакции это способность генотипа формировать в онтогенезе разные фенотипы - в зависимости от условий среды
- 7. Все биологические виды имеют определённую норму реакции генотипа, без изменения генотипа амплитуда изменений фенотипа не может
- 8. водный лютик (Ranunculus delphinifolius) и стрелолист (Sagittaria sagittifolia) формируют различные листья под водой и в воздушной
- 9. Фенотипическая изменчивость. Фенотип это совокупность всех внутренних и внешних признаков особи Изменчивость признака в пределах нормы
- 10. В фенотипической изменчивости популяции выделяют Генотипическую – наследственную Паратипическую - вызванную внешними условиями. Доля общей изменчивости,
- 11. Пример Жирность молока, содержание белка в молоке - величина наследуемости высока и колеблется от 60 до
- 12. Пример У большинства пород (популяции) кур наследуемость яйценоскости невелика (12—30%) наследуемость массы яйца значительна (60— 74%).
- 13. Внутрипопуляционная изменчивость состоит из разнообразных выражений нормы реакции по любому признаку или свойству. Изменения наследственного материала
- 14. Мутации — элементарный эволюционный материал мутации — дискретные изменения наследственной информации особи генные хромосомные геномные внеядерные
- 15. Мутации спонтанные индуцированные внешними агентами
- 16. Генные мутации изменения молекулярной структуры генов возникают в результате замен, вставок или выпадения нуклеотидов могут затрагивать
- 17. Хромосомные мутации структурные изменения хромосом возникают при перемещении или выпадении частей хромосом. инверсии транслокации нехватки дупликации.
- 18. Геномные мутации представляют изменение числа хромосом. изменение числа наборов хромосом, - полиплоидия или гаплоидия уменьшение или
- 19. Частота возникновения мутаций Частота возникновения отдельных спонтанных мутаций выражается числом гамет одного поколения, несущих определенную мутацию
- 20. Общая частота мутаций складывается из частот мутаций отдельных генов колеблется от нескольких процентов (одноклеточные водоросли, низшие
- 21. Особенности проявления мутаций. Проявление мутаций зависит от генетической среды, в которую попадает мутантный аллель Один и
- 22. Пример - экспрессивность Мутация bar («лентовидная») вызывает редукцию передних и задних фасеток глаза у дрозофилы, в
- 23. У наездника Habrobracon hebetor известен ген Kidney со 100%-ной пенетрантностью, как леталь при 30 °С, и
- 24. Спектр мутантных признаков Нет признаков и свойств, которые не затрагивались бы мутациями. вариации по средним значениям
- 25. Встречаемость мутаций в природных популяциях все популяции насыщены мутациями. нет двух популяций, имеющих одинаковые частоты встречаемости
- 26. Генетические процессы в попупяциях Сейчас известно, что все природные популяции гетерогенны - насыщены мутациями. Генетическая гетерогенность
- 29. формула Харди — Вайнберга Если частота встречаемости одного аллеля - q, частота альтернативного аллеля того же
- 30. Внутрипопуляционный полиморфизм Полиморфизм – наличие двух или более генетически различных форм в популяции Эти формы находятся
- 31. Выделяют: гетерозиготный полиморфизм адаптационный полиморфизм.
- 32. Гетерозиготный полиморфизм устанавливается в результате давления на популяцию положительного отбора гетерозигот Пример: численно равновесная экспериментальная популяция
- 33. Адаптационный полиморфизм В этом случае две или несколько генетически различных форм внутри популяции подвергаются отбору в
- 34. Гомологическая изменчивость Дарвин неоднократно упоминает о такой изменчивости, когда одинаковые признаки время от времени проявляются у
- 35. Гомологическая изменчивость Сегодня можно разделить все случаи гомологической изменчивости на три категории: полной, неполной и ложной
- 37. Скачать презентацию

Изменчивость
основное свойство живого
в основе изменчивости - конвариантная редупликация
Изменчивость
основное свойство живого
в основе изменчивости - конвариантная редупликация

ДНК – основа изменчивости
в процессе матричного копирования ДНК и РНК -
ДНК – основа изменчивости
в процессе матричного копирования ДНК и РНК -

Ч. Дарвин
вся изменчивость
наследственная
ненаследственную
ненаследственных признаков нет
все признаки и
Ч. Дарвин
вся изменчивость
наследственная
ненаследственную
ненаследственных признаков нет
все признаки и

Норма реакции
В процессе размножения передаются не признаки, а гены
гены определяют возможность
Норма реакции
В процессе размножения передаются не признаки, а гены
гены определяют возможность

Норма реакции
это способность генотипа формировать в онтогенезе разные фенотипы - в
Норма реакции
это способность генотипа формировать в онтогенезе разные фенотипы - в

Все биологические виды имеют определённую норму реакции генотипа,
без изменения генотипа
Все биологические виды имеют определённую норму реакции генотипа,
без изменения генотипа
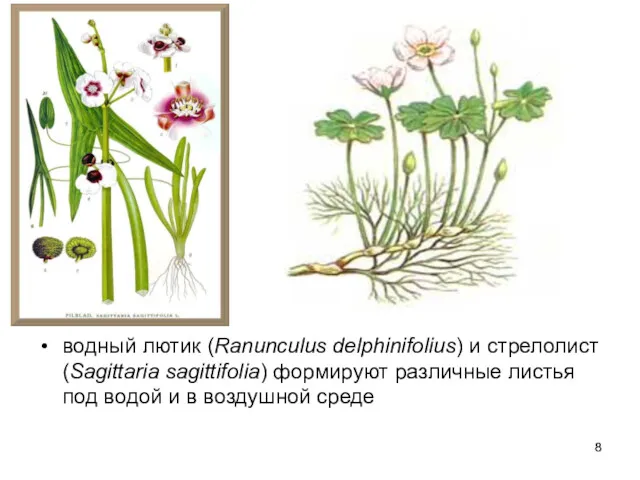
водный лютик (Ranunculus delphinifolius) и стрелолист (Sagittaria sagittifolia) формируют различные листья
водный лютик (Ranunculus delphinifolius) и стрелолист (Sagittaria sagittifolia) формируют различные листья

Фенотипическая изменчивость.
Фенотип это совокупность всех внутренних и внешних признаков особи
Фенотипическая изменчивость.
Фенотип это совокупность всех внутренних и внешних признаков особи

В фенотипической изменчивости популяции выделяют
Генотипическую – наследственную
Паратипическую - вызванную внешними условиями.
В фенотипической изменчивости популяции выделяют
Генотипическую – наследственную
Паратипическую - вызванную внешними условиями.

Пример
Жирность молока, содержание белка в молоке - величина наследуемости высока и
Пример
Жирность молока, содержание белка в молоке - величина наследуемости высока и

Пример
У большинства пород (популяции) кур наследуемость яйценоскости невелика (12—30%)
наследуемость массы яйца
Пример
У большинства пород (популяции) кур наследуемость яйценоскости невелика (12—30%)
наследуемость массы яйца

Внутрипопуляционная изменчивость
состоит из разнообразных выражений нормы реакции по любому признаку или
Внутрипопуляционная изменчивость
состоит из разнообразных выражений нормы реакции по любому признаку или

Мутации — элементарный эволюционный материал
мутации — дискретные изменения наследственной информации
Мутации — элементарный эволюционный материал
мутации — дискретные изменения наследственной информации

Мутации
спонтанные
индуцированные внешними агентами
Мутации
спонтанные
индуцированные внешними агентами

Генные мутации
изменения молекулярной структуры генов
возникают в результате замен, вставок или выпадения
Генные мутации
изменения молекулярной структуры генов
возникают в результате замен, вставок или выпадения

Хромосомные мутации
структурные изменения хромосом
возникают при перемещении или выпадении частей хромосом.
инверсии
транслокации
нехватки
Хромосомные мутации
структурные изменения хромосом
возникают при перемещении или выпадении частей хромосом.
инверсии
транслокации
нехватки

Геномные мутации
представляют изменение числа хромосом.
изменение числа наборов хромосом, - полиплоидия
Геномные мутации
представляют изменение числа хромосом.
изменение числа наборов хромосом, - полиплоидия

Частота возникновения мутаций
Частота возникновения отдельных спонтанных мутаций выражается числом гамет одного
Частота возникновения мутаций
Частота возникновения отдельных спонтанных мутаций выражается числом гамет одного

Общая частота мутаций
складывается из частот мутаций отдельных генов
колеблется от нескольких процентов
Общая частота мутаций
складывается из частот мутаций отдельных генов
колеблется от нескольких процентов

Особенности проявления мутаций.
Проявление мутаций зависит от генетической среды, в которую попадает
Особенности проявления мутаций.
Проявление мутаций зависит от генетической среды, в которую попадает

Пример - экспрессивность
Мутация bar («лентовидная») вызывает редукцию передних и задних фасеток
Пример - экспрессивность
Мутация bar («лентовидная») вызывает редукцию передних и задних фасеток

У наездника Habrobracon hebetor известен ген Kidney со 100%-ной пенетрантностью, как
У наездника Habrobracon hebetor известен ген Kidney со 100%-ной пенетрантностью, как

Спектр мутантных признаков
Нет признаков и свойств, которые не затрагивались бы мутациями.
Спектр мутантных признаков
Нет признаков и свойств, которые не затрагивались бы мутациями.

Встречаемость мутаций в природных популяциях
все популяции насыщены мутациями.
нет двух
Встречаемость мутаций в природных популяциях
все популяции насыщены мутациями.
нет двух

Генетические процессы в попупяциях
Сейчас известно, что все природные популяции гетерогенны
Генетические процессы в попупяциях
Сейчас известно, что все природные популяции гетерогенны
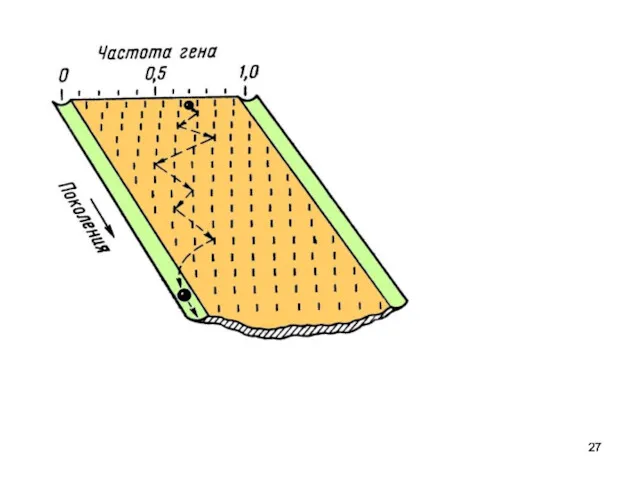
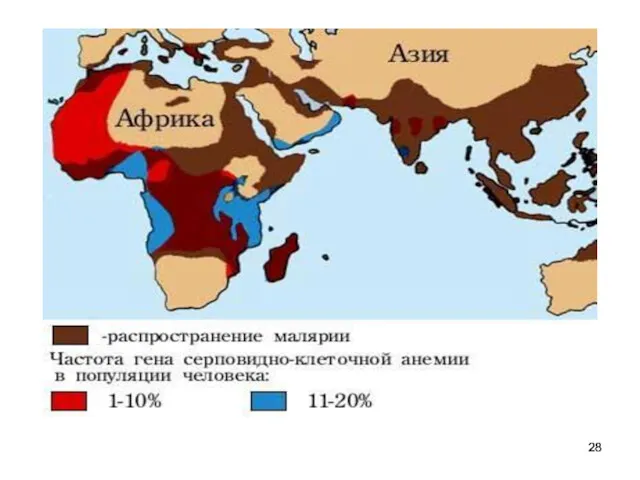

формула Харди — Вайнберга
Если частота встречаемости одного аллеля - q,
формула Харди — Вайнберга
Если частота встречаемости одного аллеля - q,

Внутрипопуляционный полиморфизм
Полиморфизм – наличие двух или более генетически различных форм в
Внутрипопуляционный полиморфизм
Полиморфизм – наличие двух или более генетически различных форм в

Выделяют:
гетерозиготный полиморфизм
адаптационный полиморфизм.
Выделяют:
гетерозиготный полиморфизм
адаптационный полиморфизм.

Гетерозиготный полиморфизм
устанавливается в результате давления на популяцию положительного отбора гетерозигот
Пример:
Гетерозиготный полиморфизм
устанавливается в результате давления на популяцию положительного отбора гетерозигот
Пример:

Адаптационный полиморфизм
В этом случае две или несколько генетически различных форм внутри
Адаптационный полиморфизм
В этом случае две или несколько генетически различных форм внутри

Гомологическая изменчивость
Дарвин неоднократно упоминает о такой изменчивости, когда одинаковые признаки время
Гомологическая изменчивость
Дарвин неоднократно упоминает о такой изменчивости, когда одинаковые признаки время

Гомологическая изменчивость
Сегодня можно разделить все случаи гомологической изменчивости на три категории:
Гомологическая изменчивость
Сегодня можно разделить все случаи гомологической изменчивости на три категории:
 Класс двудольные. Семейство Крестоцветные
Класс двудольные. Семейство Крестоцветные Селекція тварин
Селекція тварин основы возбудимости
основы возбудимости Воздействие человека и его деятельности на животных
Воздействие человека и его деятельности на животных Биология клетки. Основы молекулярной биологии клетки
Биология клетки. Основы молекулярной биологии клетки Приспособления организмов к жизни в природе
Приспособления организмов к жизни в природе Анатомия центральной нервной системы. Задний мозг
Анатомия центральной нервной системы. Задний мозг Питание животных
Питание животных Пищеварение в желудке жвачных
Пищеварение в желудке жвачных Будова і функції спинного мозку
Будова і функції спинного мозку Изменения в поведении животных в процессе приручения и одомашнивания человеком
Изменения в поведении животных в процессе приручения и одомашнивания человеком Своя игра Цветы в формате Microsoft PowerPoint
Своя игра Цветы в формате Microsoft PowerPoint Организация выращивания технического каннабиса и производства промышленных продуктов из него
Организация выращивания технического каннабиса и производства промышленных продуктов из него Симбиотические организмы лишайники
Симбиотические организмы лишайники Организменный уровень жизни. Значение и роль в природе
Организменный уровень жизни. Значение и роль в природе презентация Вооружены и опасны
презентация Вооружены и опасны Общие свойства живых организмов
Общие свойства живых организмов Круги кровообращения
Круги кровообращения Характеристика вирусов и прокариот
Характеристика вирусов и прокариот Среда обитания
Среда обитания Презентация Основные понятия и законы генетики
Презентация Основные понятия и законы генетики Деление клетки. Митоз
Деление клетки. Митоз Введение в иммунологию. Иммунная система
Введение в иммунологию. Иммунная система Генная инженерия растений
Генная инженерия растений Красная книга. Животные
Красная книга. Животные Зеленая аптека Донбасса
Зеленая аптека Донбасса Водоёмы по происхождению
Водоёмы по происхождению Широколиственный лес
Широколиственный лес