- Гормоны. Общее понятие о гормонах

Содержание
- 2. ОБЩЕЕ ПОНЯТИЕ О ГОРМОНАХ В 1849 году немецкий физиолог и зоолог Арнольд Адольф Бертольд впервые осуществил
- 3. ОБЩЕЕ ПОНЯТИЕ О ГОРМОНАХ Это дало основание профессору Лондонского Университета Эрнесту Генри Старлингу в 1905 году
- 4. ОБЩЕЕ ПОНЯТИЕ О ГОРМОНАХ Гормоны интегрируют обмен веществ, то есть регулируют соподчинённость и взаимосвязь разнообразных химических
- 5. ОБЩЕЕ ПОНЯТИЕ О ГОРМОНАХ Учение о гормонах выделено в самостоятельную науку – эндокринологию. Современная эндокринология изучает
- 6. ОБЩЕЕ ПОНЯТИЕ О ГОРМОНАХ Гормоны относятся к биологически активным веществам, определяющим в известной степени состояние физиологических
- 7. ОБЩЕЕ ПОНЯТИЕ О ГОРМОНАХ Одной из удивительных особенностей живых организмов является их способность сохранять пос-тоянство внутренней
- 8. ОБЩЕЕ ПОНЯТИЕ О ГОРМОНАХ Гормоны вызывают: либо быструю (срочную) ответную реакцию, повышая активность предобразованных, имеющихся в
- 9. ОБЩЕЕ ПОНЯТИЕ О ГОРМОНАХ К настоящему времени открыто более сотни различных веществ, наделенных гормональной активностью, синтезирующихся
- 10. ОБЩЕЕ ПОНЯТИЕ О ГОРМОНАХ Под влиянием разнообразных внешних и внутренних раздражителей возникают импульсы в специализированных, весьма
- 11. ОБЩЕЕ ПОНЯТИЕ О ГОРМОНАХ Действие гормонов на внутриклеточный обмен осуществляется через так называемые гормональные рецепторы, под
- 12. НОМЕНКЛАТУРА И КЛАССИФИКАЦИЯ ГОРМОНОВ Химическая природа почти всех известных гормонов выяснена в деталях, однако до настоящего
- 13. НОМЕНКЛАТУРА И КЛАССИФИКАЦИЯ ГОРМОНОВ Аналогичен подход и к классификации гормонов. Гормоны классифицируют в зависимости от места
- 14. НОМЕНКЛАТУРА И КЛАССИФИКАЦИЯ ГОРМОНОВ С учетом этих обстоятельств были предприняты попытки создания современной классификации гормонов, основанной
- 15. НОМЕНКЛАТУРА И КЛАССИФИКАЦИЯ ГОРМОНОВ Пептидные и белковые гормоны включают от 3 до 250 и более аминокислотных
- 16. НОМЕНКЛАТУРА И КЛАССИФИКАЦИЯ ГОРМОНОВ Гормоны стероидной природы представлены жирорастворимыми гормонами коркового вещества надпочечников (кортикостероиды), половыми гормонами
- 18. НОМЕНКЛАТУРА И КЛАССИФИКАЦИЯ ГОРМОНОВ Рассмотрим химическое строение, функции и пути биосинтеза и распада основных классов гормонов,
- 19. ГОРМОНЫ ГИПОТАЛАМУСА Гипоталамус служит местом непосредственного взаимодействия высших отделов ЦНС и эндокринной системы. Это взаимодействие осуществляется
- 20. ГОРМОНЫ ГИПОТАЛАМУСА
- 21. ГОРМОНЫ ГИПОТАЛАМУСА В гипоталамусе открыто 7 стимуляторов (либерины) и 3 ингибитора (статины) секреции гормонов гипофиза, а
- 22. ГОРМОНЫ ГИПОТАЛАМУСА Не все гормоны гипоталамуса строго специфичны в отношении одного какого-либо гипофизарного гормона. В частности,
- 23. Гипоталомические гормоны, контролирующие освобождение гормонов гипофиза
- 24. По химическому строению все гормоны гипоталамуса являются низкомолекулярными пептидами, так называемыми олигопептидами необычного строения. Приводим полученные
- 25. 2. Гонадолиберин является декапептидом, состоящим из 10 аминокислот в последовательности: Пиро-Глу–Гис–Трп–Сер–Тир–Гли–Лей–Арг–Про–Гли-NН2 Концевая С-аминокислота представлена глицинамидом. ГОРМОНЫ
- 26. ГОРМОНЫ ГИПОТАЛАМУСА 3. Соматостатин является циклическим тетрадекапептидом (состоит из 14 аминокислотных остатков): Отличается этот гормон от
- 27. ГОРМОНЫ ГИПОТАЛАМУСА. Соматостатин
- 28. ГОРМОНЫ ГИПОТАЛАМУСА 4. Соматолиберин выделен из природных источников. Он представлен 44 аминокислотными остатками с полностью раскрытой
- 29. ГОРМОНЫ ГИПОТАЛАМУСА 6. Меланостатин (меланотропинингибирующий фактор) представлен или трипептидом: Пиро-Глу–Лей–Гли-NН2, или пентапептидом со следующей последовательностью: Пиро-Глу–Гис–Фен–Aрг–Гли–NН2.
- 30. ГОРМОНЫ ГИПОТАЛАМУСА Местом синтеза гипоталамических гормонов, являются нервные окончания – синаптосомы гипоталамуса, поскольку именно там отмечена
- 31. ГОРМОНЫ ГИПОТАЛАМУСА Гипоталамические гормоны непосредственно влияют на секрецию (точнее, освобождение) «готовых» гормонов и биосинтез этих гормонов
- 32. ГОРМОНЫ ГИПОФИЗА В гипофизе синтезируется ряд биологически активных гормонов белковой и пептидной природы, оказывающих стимулирующий эффект
- 33. ГОРМОНЫ ГИПОФИЗА
- 34. ГОРМОНЫ ГИПОФИЗА Из ткани мозга животных было выделено более 50 пептидов, получивших название нейропептидов и определяющих
- 35. Вазопрессин и окситоцин Гормоны вазопрессин и окситоцин синтезируются рибосомальным путем, причем одновременно в гипоталамусе синтезируются 3
- 36. Вазопрессин и окситоцин Химическое строение обоих гормонов было расшифровано классическими работами американского биохимика Винсента дю Виньо
- 37. Вазопрессин и окситоцин Вазопрессин отличается от окситоцина двумя аминокислотами: он содержит в положении 3 от N-конца
- 38. Вазопрессин и окситоцин Основной биологический эффект окситоцина у млекопитающих связан со стимуляцией сокращения гладких мышц матки
- 40. Вазопрессин и окситоцин Вазопрессин стимулирует сокращение гладких мышечных волокон сосудов, оказывая сильное вазопрессорное действие, однако основная
- 41. Меланоцитстимулирующие гормоны (МСГ, меланотропины) Меланотропины синтезируются и секретируются в кровь промежуточной долей гипофиза. Выделены и расшифрованы
- 42. Меланоцитстимулирующие гормоны (МСГ, меланотропины) Состав и структура β-МСГ оказались более сложными. У большинства животных молекула β-МСГ
- 43. Адренокортикотропный гормон (АКТГ, кортикотропин) Еще в 1926 г. было установлено, что гипофиз оказывает стимулирующее влияние на
- 44. Адренокортикотропный гормон (АКТГ, кортикотропин) Различия в структуре АКТГ овцы, свиньи и быка касаются только природы 31-го
- 45. Соматотропный гормон (СТГ, гормон роста, соматотропин) Гормон роста был открыт в экстрактах передней доли гипофиза еще
- 46. Соматотропный гормон (СТГ, гормон роста, соматотропин) СТГ обладает широким спектром биологического действия. Он влияет на все
- 47. Соматотропный гормон (СТГ, гормон роста, соматотропин) СТГ регулирует процессы роста и развития всего организме, что подтверждается
- 48. Лактотропный гормон (пролактин, лютеотропный гормон) Пролактин считается одним из наиболее «древних» гормонов гипофиза, поскольку его удается
- 49. Лактотропный гормон (пролактин, лютеотропный гормон) Расшифрована структура пролактина из гипофиза овцы, быка и человека. Это крупный
- 50. Тиреотропный гормон (ТТГ, тиротропин) Тиротропин является сложным гликопротеином, выделяемым передней долей гипофиза и содержит по две
- 51. Гонадотропные гормоны (гонадотррпины) К гонадотропинам относятся фолликулостимулирующий гормон (ФСГ, фоллитропин) и лютеинизирующий гормон (ЛГ, лютропин), или
- 52. Липотропные гормоны (ЛТГ, липотропины) Среди гормонов передней доли гипофиза следует отметить липотропины, в частности β- и
- 53. Липотропные гормоны (ЛТГ, липотропины) Перечисленные биологические свойства обусловлены не β-липотропином, оказавшимся лишенным гормональной активности, а продуктами
- 54. Липотропные гормоны (ЛТГ, липотропины) Общим типом структуры для всех трех соединений является тетрапептидная последовательность на N-конце.
- 55. Липотропные гормоны (ЛТГ, липотропины) Ниже представлены пептидные гормоны, образующиеся из β-липотропина путем специфического протеолиза.
- 56. Липотропные гормоны (ЛТГ, липотропины) Учитывая исключительную роль β-липотропина как предшественника перечисленных гормонов, приводим первичную структуру β-липотропина
- 57. ГОРМОНЫ ПАРАЩИТОВИДНЫХ ЖЕЛЕЗ (ПАРАТГОРМОНЫ) К гормонам белковой природы относится группа паратгормонов (паратиреоидных гормонов), различающихся последовательностью аминокислот.
- 58. ГОРМОНЫ ПАРАЩИТОВИДНЫХ ЖЕЛЕЗ (ПАРАТГОРМОНЫ) Паратгормон участвует в регуляции концентрации катионов кальция и связанных с ними анионов
- 59. ГОРМОНЫ ПАРАЩИТОВИДНЫХ ЖЕЛЕЗ (ПАРАТГОРМОНЫ) Другой орган-мишень паратгормона – это почка. Паратгормон уменьшает реабсорбцию фосфата в дистальных
- 60. ГОРМОНЫ ПАРАЩИТОВИДНЫХ ЖЕЛЕЗ (ПАРАТГОРМОНЫ)
- 61. ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ Щитовидная железа играет исключительно важную роль в обмене веществ. Об этом свидетельствуют резкое
- 62. ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ Впервые на существование кальцитонина, обладающего способностью поддерживать постоянный уровень кальция в крови, указал
- 63. ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ Кальцитонин человека содержит дисульфидный мостик (между 1-м и 7-м аминокислотными остатками) и характеризуется
- 64. ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ Химическая природа гормонов фолликулярной части щитовидной железы выяснена в деталях. Все йодсодержащие гормоны,
- 65. ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ Из L-тиронина легко синтезируется гормон щитовидной железы тироксин, содержащий в 4 положениях кольцевой
- 66. ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ Катаболизм гормонов щитовидной железы протекает по двум направлениям: распад гормонов с освобождением йода
- 67. ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ Гипофункция щитовидной железы в раннем детском возрасте приводит к развитию болезни, известной в
- 68. ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ Следует отметить еще одно поражение щитовидной железы – эндемический зоб. Болезнь обычно развивается
- 69. ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ При гиперфункции щитовидной железы и показано оперативное удаление всей железы или введение 131I
- 70. ГОРМОНЫ ПОДЖЕЛУДОЧНОЙ ЖЕЛЕЗЫ Поджелудочная железа относится к железам со смешанной секрецией. Внешнесекреторная функция ее заключается в
- 71. Инсулин Инсулин получивший свое название от наименования панкреатических островков (лат. insula – островок), был первым белком,
- 72. Инсулин Биосинтез инсулина осуществляется в β-клетках панкреатических островков из своего предшественника проинсулина. Уже не только выяснена
- 73. Инсулин Синтезированный из проинсулина инсулин может существовать в нескольких формах, различающихся по биологическим, иммунологическим и физико-химическим
- 74. Инсулин В физиологической регуляции синтеза инсулина доминирующую роль играет концентрация глюкозы в крови. Так, повышение содержания
- 75. Инсулин При недостаточной секреции (точнее, недостаточном синтезе) инсулина развивается специфическое заболевание – сахарный диабет. Помимо клинически
- 76. Инсулин Инсулиновый рецептор (схема). Две α-цепи на наружной поверхности мембраны клетки и две трансмембранные β-цепи. Связывание
- 77. Инсулин Наиболее вероятной в настоящее время представляется мембранная локализация первичного действия почти всех белковых гормонов, включая
- 78. Глюкагон Глюкагон впервые был обнаружен в коммерческих препаратах инсулина еще в 1923 г., однако только в
- 79. Глюкагон По биологическому действию глюкагон относятся к гипергликемическим факторам, вызывает увеличение концентрации глюкозы в крови главным
- 80. ГОРМОНЫ НАДПОЧЕЧНИКОВ Надпочечники состоят из двух индивидуальных в морфологическом и функциональном отношениях частей – мозгового и
- 81. Гормоны мозгового вещества надпочечников О способности экстрактов из надпочечников повышать кровяное давление было известно еще в
- 82. Гормоны мозгового вещества надпочечников Эти гормоны по строению напоминают аминокислоту тирозин, от которого они отличаются наличием
- 83. Гормоны мозгового вещества надпочечников В мозговом веществе надпочечников человека массой 10 г содержится около 5 мг
- 84. Гормоны коркового вещества надпочечников Со второй половины XIX в. известно заболевание, названное бронзовой болезнью, или болезнью
- 85. Химическое строение, биосинтез и биологическое действие кортикостероидов К настоящему времени из коркового вещества надпочечников человека, свиньи
- 86. Химическое строение, биосинтез и биологическое действие кортикостероидов
- 87. Химическое строение, биосинтез и биологическое действие кортикостероидов Предшественником кортикостероидов является холестерин(ол). В свою очередь синтез АКТГ
- 88. Химическое строение, биосинтез и биологическое действие кортикостероидов Ферменты катализируют минимум две последовательные реакции гидроксилирования и реакцию
- 89. Химическое строение, биосинтез и биологическое действие кортикостероидов Минералокортикоиды (дезоксикортикостерон и альдостерон) регулируют главным образом обмен натрия,
- 90. Химическое строение, биосинтез и биологическое действие кортикостероидов
- 91. ПОЛОВЫЕ ГОРМОНЫ Половые гормоны синтезируются в основном в половых железах женщин (яичники) и мужчин (семенники); некоторое
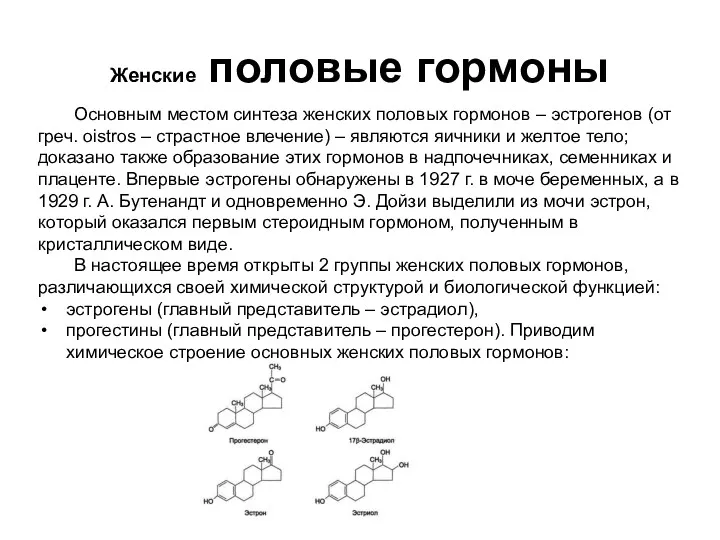
- 92. Женские половые гормоны Основным местом синтеза женских половых гормонов – эстрогенов (от греч. oistros – страстное
- 93. Женские половые гормоны Наиболее активный эстроген – эстрадиол, синтезируется преимущественно в фолликулах; два остальных эстрогена являются
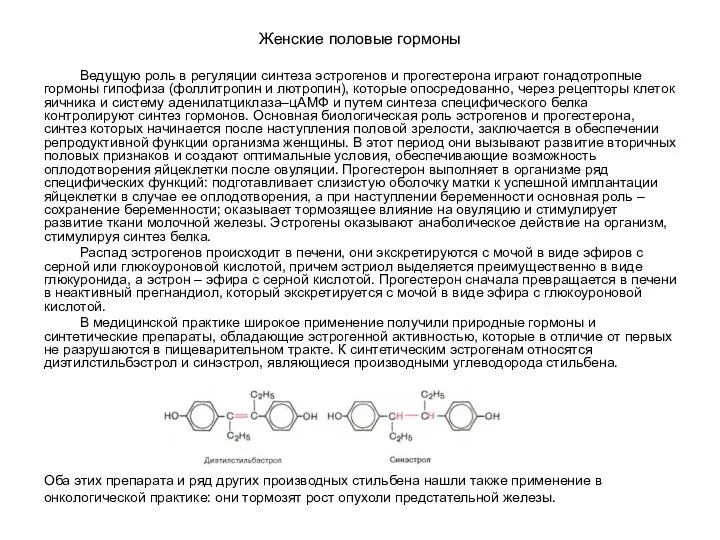
- 94. Женские половые гормоны Ведущую роль в регуляции синтеза эстрогенов и прогестерона играют гонадотропные гормоны гипофиза (фоллитропин
- 95. Мужские половые гормоны Внутрисекреторная функция мужских половых желез была установлена в 1849 г., однако только в
- 96. Мужские половые гормоны Биосинтез андрогенов осуществляется главным образом в семенниках и частично в яичниках и надпочечниках.
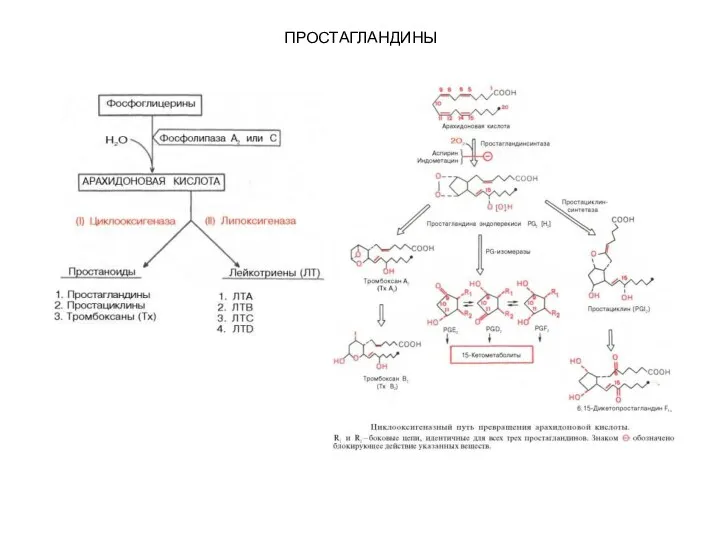
- 97. ПРОСТАГЛАНДИНЫ Термин «простагландины» был введен У. Эйлером, впервые показавшим, что в сперме человека и экстрактах из
- 98. ПРОСТАГЛАНДИНЫ
- 99. ПРОСТАГЛАНДИНЫ Первый путь получил наименование циклооксигеназного пути превращения арахидоновой кислоты, поскольку первые стадии синтеза простагландинов катализируются
- 100. ПРОСТАГЛАНДИНЫ
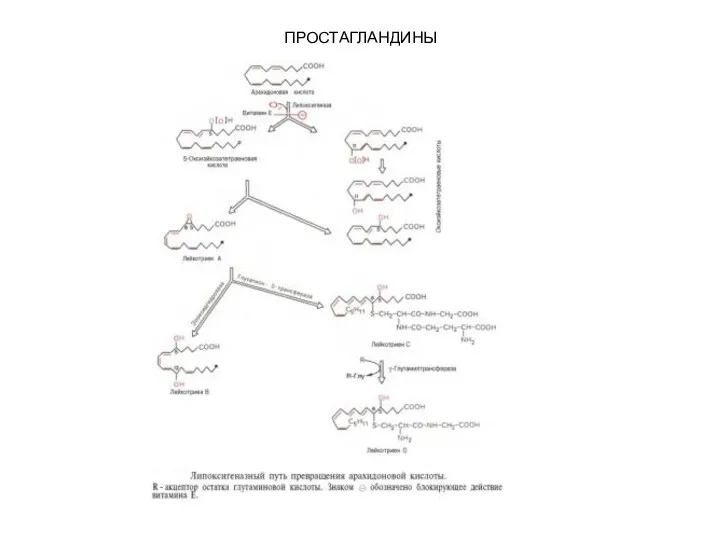
- 101. ПРОСТАГЛАНДИНЫ Второй путь превращения арахидоновой кислоты – липоксигеназный путь – отличается тем, что дает начало синтезу
- 102. ГОРМОНЫ ВИЛОЧКОВОЙ ЖЕЛЕЗЫ (ТИМУСА) Роль тимуса как эндокринной железы известна давно. Известно, что тимус вскоре после
- 103. Все известные к настоящему времени гормональные вещества описать почти не возможно, их слишком много. В шишковидной
- 104. МОЛЕКУЛЯРНЫЕ МЕХАНИЗМЫ ПЕРЕДАЧИ ГОРМОНАЛЬНОГО СИГНАЛА Несмотря на огромное разнообразие гормонов и гормоноподобных веществ, в основе биологического
- 105. МОЛЕКУЛЯРНЫЕ МЕХАНИЗМЫ ПЕРЕДАЧИ ГОРМОНАЛЬНОГО СИГНАЛА Общим фундаментальным механизмом, посредством которого реализуются биологические эффекты «вторичных» мессенджеров внутри
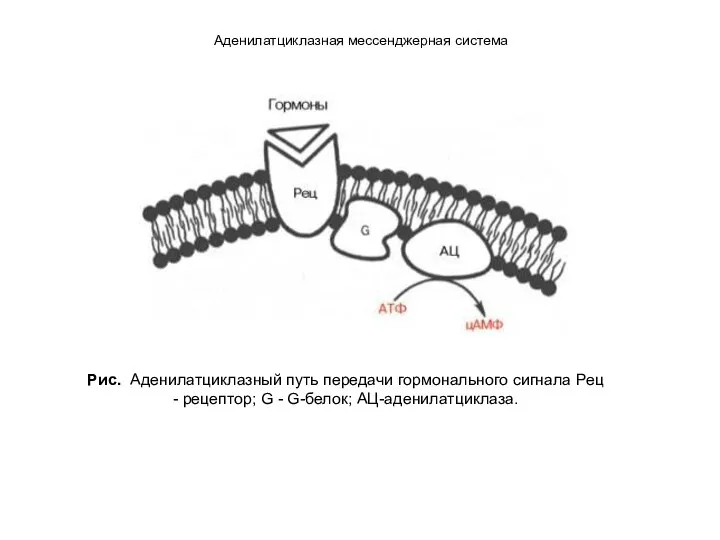
- 106. Аденилатциклазная мессенджерная система Наиболее изученным является аденилатциклазный путь передачи гормонального сигнала. В нем задействовано мимимум пять
- 107. Аденилатциклазная мессенджерная система Рис. Аденилатциклазный путь передачи гормонального сигнала Рец - рецептор; G - G-белок; АЦ-аденилатциклаза.
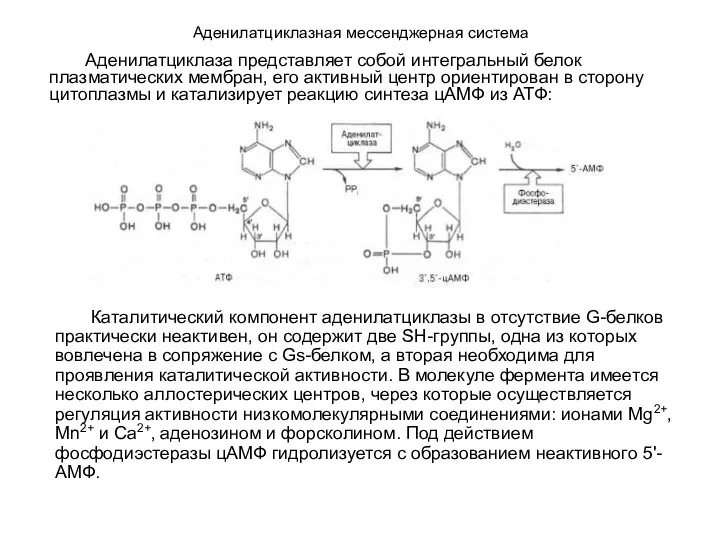
- 108. Аденилатциклазная мессенджерная система Каталитический компонент аденилатциклазы в отсутствие G-белков практически неактивен, он содержит две SH-группы, одна
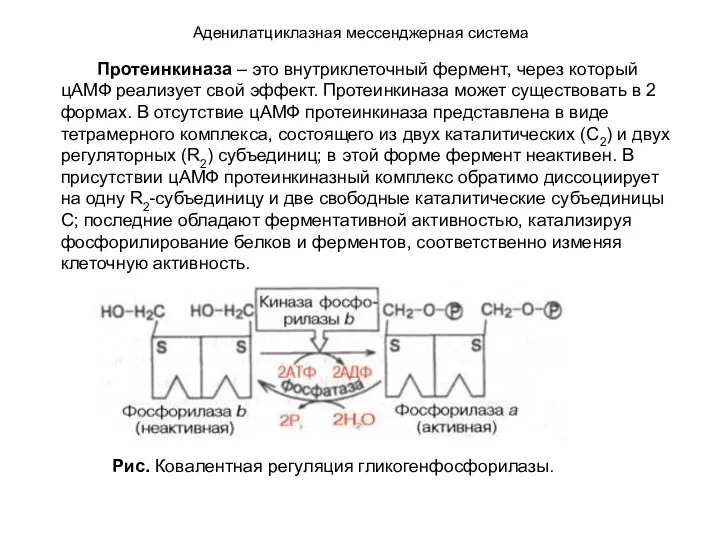
- 109. Аденилатциклазная мессенджерная система Протеинкиназа – это внутриклеточный фермент, через который цАМФ реализует свой эффект. Протеинкиназа может
- 110. Аденилатциклазная мессенджерная система В клетках открыт большой класс цАМФ-зависимых протеинкиназ, названных протеинкиназами А; они катализируют перенос
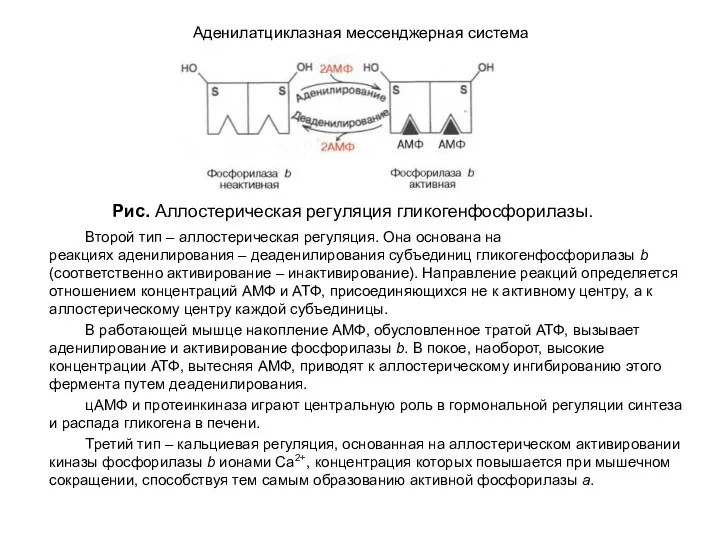
- 111. Аденилатциклазная мессенджерная система Второй тип – аллостерическая регуляция. Она основана на реакциях аденилирования – деаденилирования субъединиц
- 112. Гуанилатциклазная мессенджерная система Довольно долгое время циклический гуанозинмонофосфат (цГМФ) рассматривался как антипод цАМФ. Ему приписывали функции,
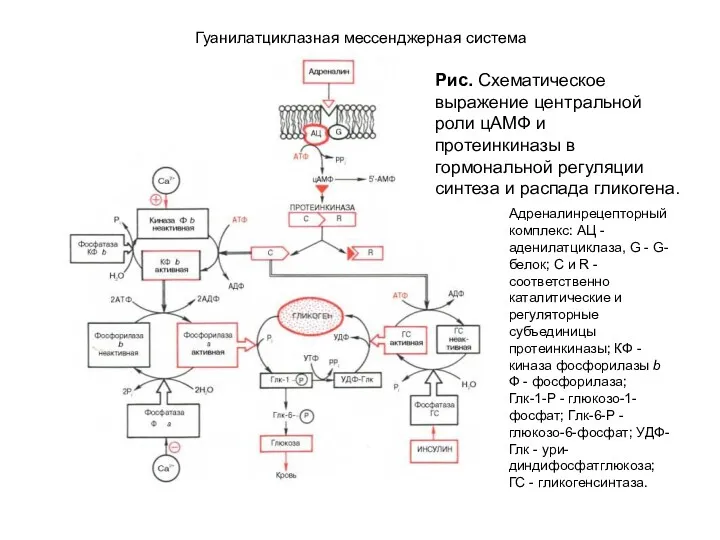
- 113. Гуанилатциклазная мессенджерная система Рис. Схематическое выражение центральной роли цАМФ и протеинкиназы в гормональной регуляции синтеза и
- 114. Гуанилатциклазная мессенджерная система Известны четыре разные формы гуанилатциклазы, три из которых являются мембраносвязанными и одна –
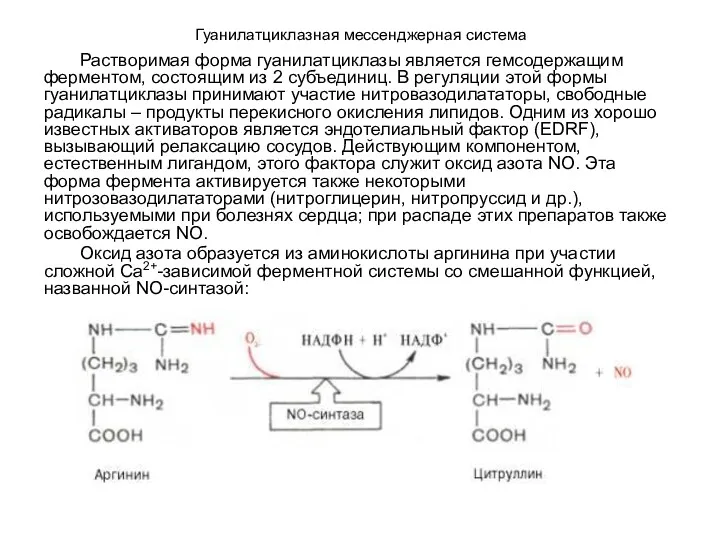
- 115. Гуанилатциклазная мессенджерная система Растворимая форма гуанилатциклазы является гемсодержащим ферментом, состоящим из 2 субъединиц. В регуляции этой
- 116. Гуанилатциклазная мессенджерная система Оксид азота при взаимодействии с гемом гуанилатциклазы способствует быстрому образованию цГМФ, который снижает
- 117. Са2+-мессенджерная система Ионам Са2+ принадлежит центральная роль в регуляции многих клеточных функций. Изменение концентрации внутриклеточного свободного
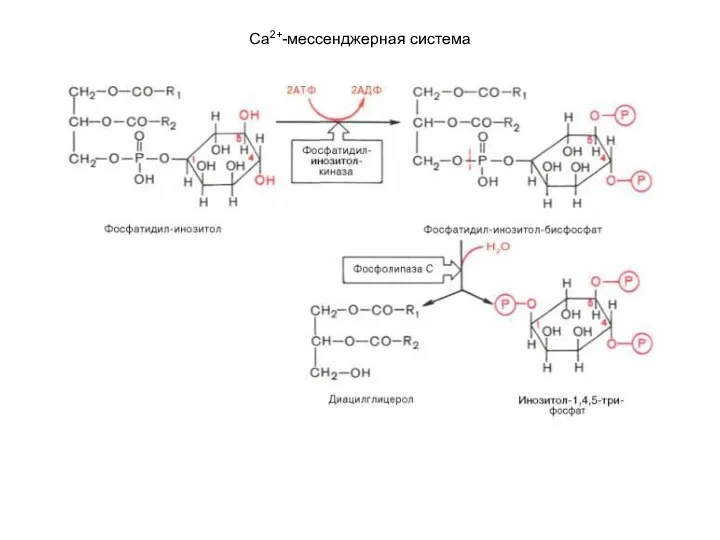
- 118. Са2+-мессенджерная система К внутриклеточной системе мессенджеров относят также производные фосфолипидов мембран эукариотических клеток, в частности фосфорилированные
- 119. Са2+-мессенджерная система
- 121. Скачать презентацию

ОБЩЕЕ ПОНЯТИЕ О ГОРМОНАХ
В 1849 году немецкий физиолог и зоолог Арнольд
ОБЩЕЕ ПОНЯТИЕ О ГОРМОНАХ
В 1849 году немецкий физиолог и зоолог Арнольд

ОБЩЕЕ ПОНЯТИЕ О ГОРМОНАХ
Это дало основание профессору Лондонского Университета Эрнесту Генри
ОБЩЕЕ ПОНЯТИЕ О ГОРМОНАХ
Это дало основание профессору Лондонского Университета Эрнесту Генри

ОБЩЕЕ ПОНЯТИЕ О ГОРМОНАХ
Гормоны интегрируют обмен веществ, то есть регулируют соподчинённость
ОБЩЕЕ ПОНЯТИЕ О ГОРМОНАХ
Гормоны интегрируют обмен веществ, то есть регулируют соподчинённость

ОБЩЕЕ ПОНЯТИЕ О ГОРМОНАХ
Учение о гормонах выделено в самостоятельную науку – эндокринологию.
ОБЩЕЕ ПОНЯТИЕ О ГОРМОНАХ
Учение о гормонах выделено в самостоятельную науку – эндокринологию.

ОБЩЕЕ ПОНЯТИЕ О ГОРМОНАХ
Гормоны относятся к биологически активным веществам, определяющим в
ОБЩЕЕ ПОНЯТИЕ О ГОРМОНАХ
Гормоны относятся к биологически активным веществам, определяющим в

ОБЩЕЕ ПОНЯТИЕ О ГОРМОНАХ
Одной из удивительных особенностей живых организмов является их
ОБЩЕЕ ПОНЯТИЕ О ГОРМОНАХ
Одной из удивительных особенностей живых организмов является их

ОБЩЕЕ ПОНЯТИЕ О ГОРМОНАХ
Гормоны вызывают:
либо быструю (срочную) ответную реакцию, повышая активность предобразованных,
ОБЩЕЕ ПОНЯТИЕ О ГОРМОНАХ
Гормоны вызывают:
либо быструю (срочную) ответную реакцию, повышая активность предобразованных,

ОБЩЕЕ ПОНЯТИЕ О ГОРМОНАХ
К настоящему времени открыто более сотни различных веществ,
ОБЩЕЕ ПОНЯТИЕ О ГОРМОНАХ
К настоящему времени открыто более сотни различных веществ,

ОБЩЕЕ ПОНЯТИЕ О ГОРМОНАХ
Под влиянием разнообразных внешних и внутренних раздражителей возникают
ОБЩЕЕ ПОНЯТИЕ О ГОРМОНАХ
Под влиянием разнообразных внешних и внутренних раздражителей возникают

ОБЩЕЕ ПОНЯТИЕ О ГОРМОНАХ
Действие гормонов на внутриклеточный обмен осуществляется через так
ОБЩЕЕ ПОНЯТИЕ О ГОРМОНАХ
Действие гормонов на внутриклеточный обмен осуществляется через так

НОМЕНКЛАТУРА И КЛАССИФИКАЦИЯ ГОРМОНОВ
Химическая природа почти всех известных гормонов выяснена в
НОМЕНКЛАТУРА И КЛАССИФИКАЦИЯ ГОРМОНОВ
Химическая природа почти всех известных гормонов выяснена в

НОМЕНКЛАТУРА И КЛАССИФИКАЦИЯ ГОРМОНОВ
Аналогичен подход и к классификации гормонов.
Гормоны классифицируют в
НОМЕНКЛАТУРА И КЛАССИФИКАЦИЯ ГОРМОНОВ
Аналогичен подход и к классификации гормонов.
Гормоны классифицируют в

НОМЕНКЛАТУРА И КЛАССИФИКАЦИЯ ГОРМОНОВ
С учетом этих обстоятельств были предприняты попытки создания
НОМЕНКЛАТУРА И КЛАССИФИКАЦИЯ ГОРМОНОВ
С учетом этих обстоятельств были предприняты попытки создания

НОМЕНКЛАТУРА И КЛАССИФИКАЦИЯ ГОРМОНОВ
Пептидные и белковые гормоны включают от 3 до
НОМЕНКЛАТУРА И КЛАССИФИКАЦИЯ ГОРМОНОВ
Пептидные и белковые гормоны включают от 3 до

НОМЕНКЛАТУРА И КЛАССИФИКАЦИЯ ГОРМОНОВ
Гормоны стероидной природы представлены жирорастворимыми гормонами коркового вещества
НОМЕНКЛАТУРА И КЛАССИФИКАЦИЯ ГОРМОНОВ
Гормоны стероидной природы представлены жирорастворимыми гормонами коркового вещества
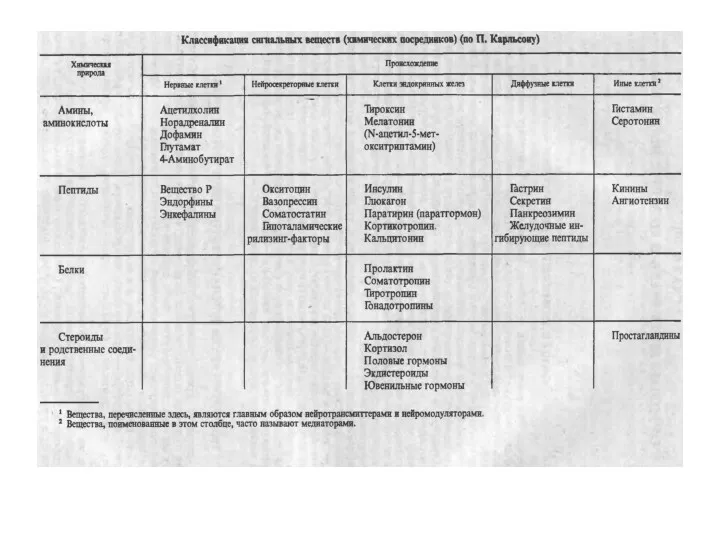

НОМЕНКЛАТУРА И КЛАССИФИКАЦИЯ ГОРМОНОВ
Рассмотрим химическое строение, функции и пути биосинтеза и
НОМЕНКЛАТУРА И КЛАССИФИКАЦИЯ ГОРМОНОВ
Рассмотрим химическое строение, функции и пути биосинтеза и

ГОРМОНЫ ГИПОТАЛАМУСА
Гипоталамус служит местом непосредственного взаимодействия высших отделов ЦНС и эндокринной
ГОРМОНЫ ГИПОТАЛАМУСА
Гипоталамус служит местом непосредственного взаимодействия высших отделов ЦНС и эндокринной
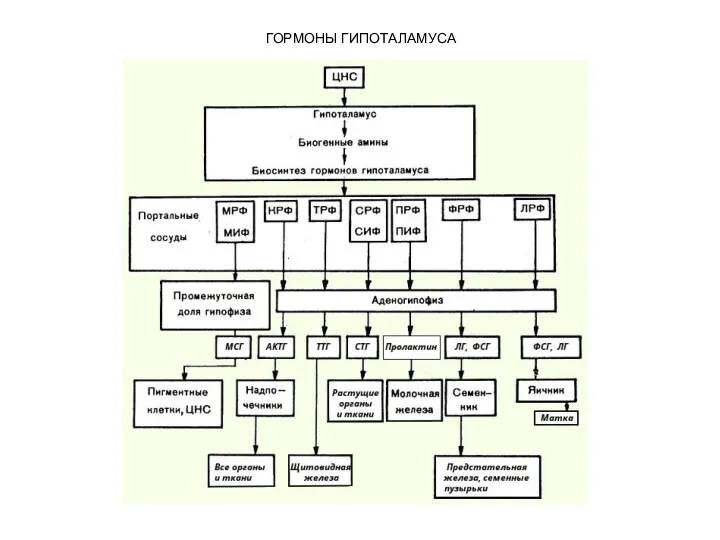
ГОРМОНЫ ГИПОТАЛАМУСА
ГОРМОНЫ ГИПОТАЛАМУСА

ГОРМОНЫ ГИПОТАЛАМУСА
В гипоталамусе открыто 7 стимуляторов (либерины) и 3 ингибитора (статины)
ГОРМОНЫ ГИПОТАЛАМУСА
В гипоталамусе открыто 7 стимуляторов (либерины) и 3 ингибитора (статины)

ГОРМОНЫ ГИПОТАЛАМУСА
Не все гормоны гипоталамуса строго специфичны в отношении одного какого-либо
ГОРМОНЫ ГИПОТАЛАМУСА
Не все гормоны гипоталамуса строго специфичны в отношении одного какого-либо
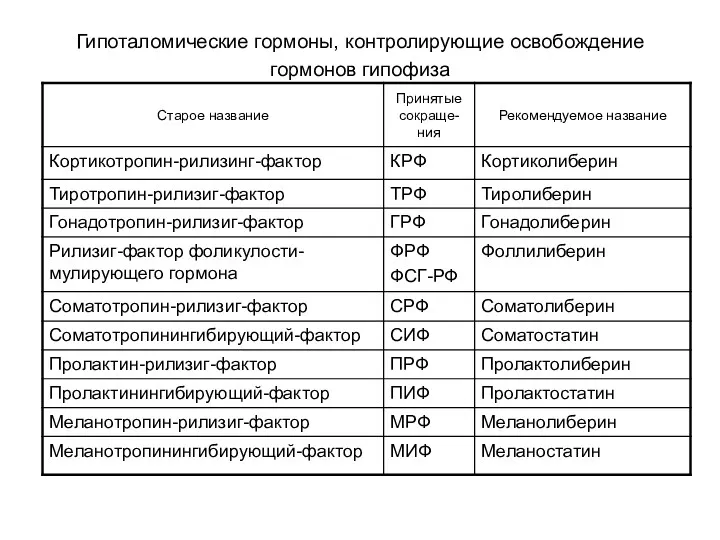
Гипоталомические гормоны, контролирующие освобождение гормонов гипофиза
Гипоталомические гормоны, контролирующие освобождение гормонов гипофиза
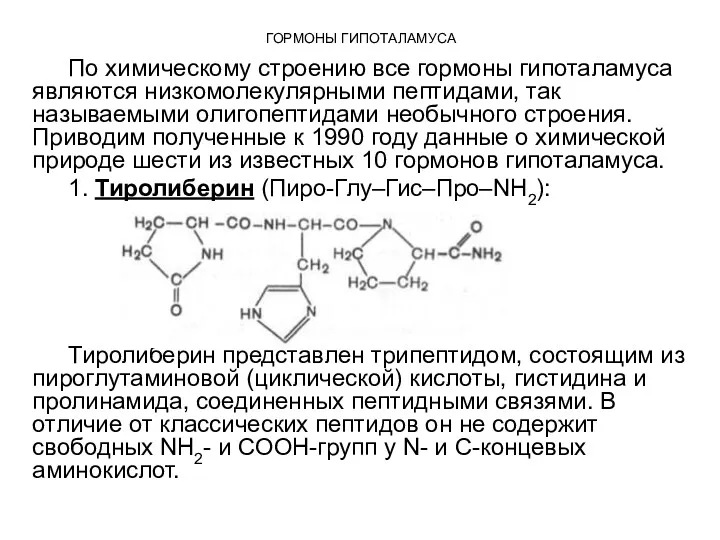
По химическому строению все гормоны гипоталамуса являются низкомолекулярными пептидами, так называемыми
По химическому строению все гормоны гипоталамуса являются низкомолекулярными пептидами, так называемыми
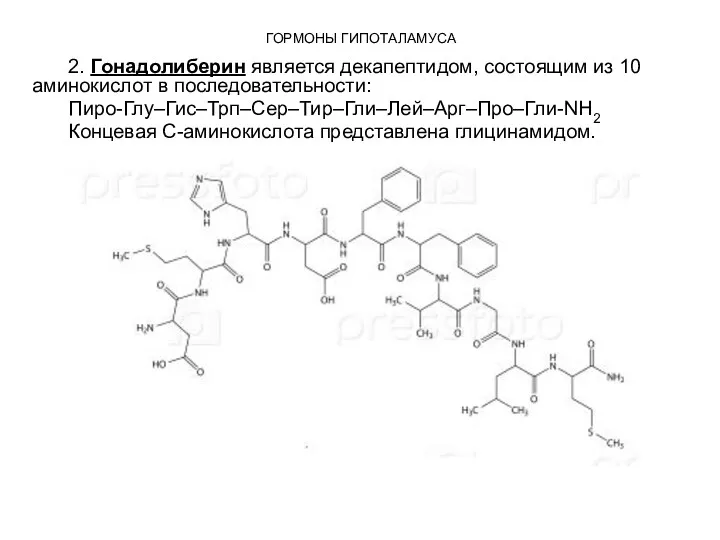
2. Гонадолиберин является декапептидом, состоящим из 10 аминокислот в последовательности:
Пиро-Глу–Гис–Трп–Сер–Тир–Гли–Лей–Арг–Про–Гли-NН2
Концевая С-аминокислота представлена
2. Гонадолиберин является декапептидом, состоящим из 10 аминокислот в последовательности:
Пиро-Глу–Гис–Трп–Сер–Тир–Гли–Лей–Арг–Про–Гли-NН2
Концевая С-аминокислота представлена

ГОРМОНЫ ГИПОТАЛАМУСА
3. Соматостатин является циклическим тетрадекапептидом (состоит из 14 аминокислотных остатков):
Отличается этот
ГОРМОНЫ ГИПОТАЛАМУСА
3. Соматостатин является циклическим тетрадекапептидом (состоит из 14 аминокислотных остатков):
Отличается этот
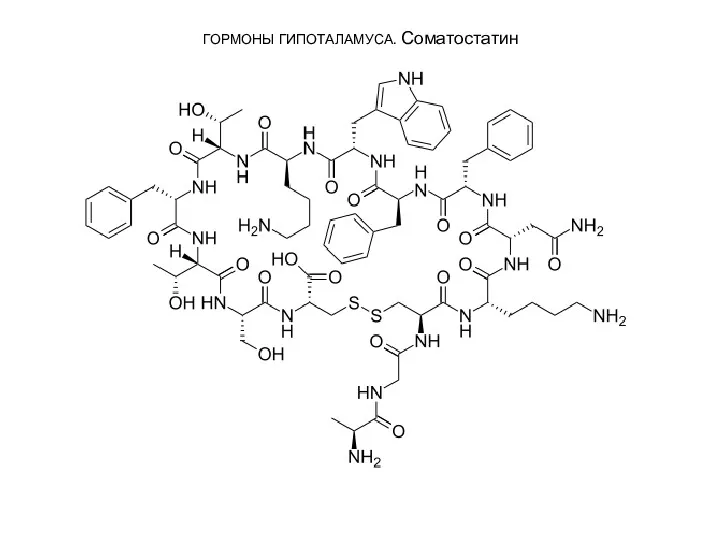
ГОРМОНЫ ГИПОТАЛАМУСА. Соматостатин
ГОРМОНЫ ГИПОТАЛАМУСА. Соматостатин

ГОРМОНЫ ГИПОТАЛАМУСА
4. Соматолиберин выделен из природных источников. Он представлен 44 аминокислотными остатками
ГОРМОНЫ ГИПОТАЛАМУСА
4. Соматолиберин выделен из природных источников. Он представлен 44 аминокислотными остатками

ГОРМОНЫ ГИПОТАЛАМУСА
6. Меланостатин (меланотропинингибирующий фактор) представлен или трипептидом:
Пиро-Глу–Лей–Гли-NН2,
или пентапептидом со следующей последовательностью:
Пиро-Глу–Гис–Фен–Aрг–Гли–NН2.
Меланолиберин оказывает
ГОРМОНЫ ГИПОТАЛАМУСА
6. Меланостатин (меланотропинингибирующий фактор) представлен или трипептидом:
Пиро-Глу–Лей–Гли-NН2,
или пентапептидом со следующей последовательностью:
Пиро-Глу–Гис–Фен–Aрг–Гли–NН2.
Меланолиберин оказывает

ГОРМОНЫ ГИПОТАЛАМУСА
Местом синтеза гипоталамических гормонов, являются нервные окончания – синаптосомы гипоталамуса,
ГОРМОНЫ ГИПОТАЛАМУСА
Местом синтеза гипоталамических гормонов, являются нервные окончания – синаптосомы гипоталамуса,

ГОРМОНЫ ГИПОТАЛАМУСА
Гипоталамические гормоны непосредственно влияют на секрецию (точнее, освобождение) «готовых» гормонов
ГОРМОНЫ ГИПОТАЛАМУСА
Гипоталамические гормоны непосредственно влияют на секрецию (точнее, освобождение) «готовых» гормонов

ГОРМОНЫ ГИПОФИЗА
В гипофизе синтезируется ряд биологически активных гормонов белковой и пептидной
ГОРМОНЫ ГИПОФИЗА
В гипофизе синтезируется ряд биологически активных гормонов белковой и пептидной

ГОРМОНЫ ГИПОФИЗА
ГОРМОНЫ ГИПОФИЗА

ГОРМОНЫ ГИПОФИЗА
Из ткани мозга животных было выделено более 50 пептидов, получивших
ГОРМОНЫ ГИПОФИЗА
Из ткани мозга животных было выделено более 50 пептидов, получивших

Вазопрессин и окситоцин
Гормоны вазопрессин и окситоцин синтезируются рибосомальным путем, причем одновременно
Вазопрессин и окситоцин
Гормоны вазопрессин и окситоцин синтезируются рибосомальным путем, причем одновременно

Вазопрессин и окситоцин
Химическое строение обоих гормонов было расшифровано классическими работами американского
Вазопрессин и окситоцин
Химическое строение обоих гормонов было расшифровано классическими работами американского

Вазопрессин и окситоцин
Вазопрессин отличается от окситоцина двумя аминокислотами: он содержит в
Вазопрессин и окситоцин
Вазопрессин отличается от окситоцина двумя аминокислотами: он содержит в

Вазопрессин и окситоцин
Основной биологический эффект окситоцина у млекопитающих связан со стимуляцией
Вазопрессин и окситоцин
Основной биологический эффект окситоцина у млекопитающих связан со стимуляцией
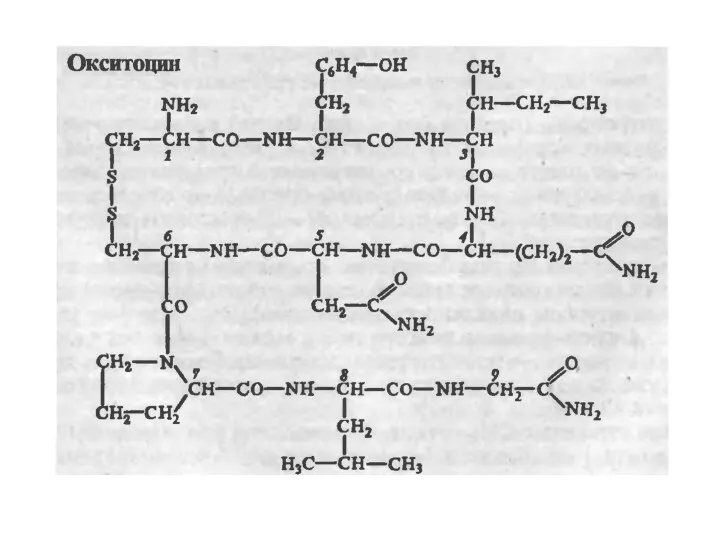

Вазопрессин и окситоцин
Вазопрессин стимулирует сокращение гладких мышечных волокон сосудов, оказывая сильное
Вазопрессин и окситоцин
Вазопрессин стимулирует сокращение гладких мышечных волокон сосудов, оказывая сильное

Меланоцитстимулирующие гормоны (МСГ, меланотропины)
Меланотропины синтезируются и секретируются в кровь промежуточной долей
Меланоцитстимулирующие гормоны (МСГ, меланотропины)
Меланотропины синтезируются и секретируются в кровь промежуточной долей

Меланоцитстимулирующие гормоны (МСГ, меланотропины)
Состав и структура β-МСГ оказались более сложными. У
Меланоцитстимулирующие гормоны (МСГ, меланотропины)
Состав и структура β-МСГ оказались более сложными. У

Адренокортикотропный гормон (АКТГ, кортикотропин)
Еще в 1926 г. было установлено, что гипофиз
Адренокортикотропный гормон (АКТГ, кортикотропин)
Еще в 1926 г. было установлено, что гипофиз

Адренокортикотропный гормон (АКТГ, кортикотропин)
Различия в структуре АКТГ овцы, свиньи и быка
Адренокортикотропный гормон (АКТГ, кортикотропин)
Различия в структуре АКТГ овцы, свиньи и быка

Соматотропный гормон (СТГ, гормон роста, соматотропин)
Гормон роста был открыт в экстрактах
Соматотропный гормон (СТГ, гормон роста, соматотропин)
Гормон роста был открыт в экстрактах

Соматотропный гормон (СТГ, гормон роста, соматотропин)
СТГ обладает широким спектром биологического действия.
Соматотропный гормон (СТГ, гормон роста, соматотропин)
СТГ обладает широким спектром биологического действия.

Соматотропный гормон (СТГ, гормон роста, соматотропин)
СТГ регулирует процессы роста и развития
Соматотропный гормон (СТГ, гормон роста, соматотропин)
СТГ регулирует процессы роста и развития

Лактотропный гормон (пролактин, лютеотропный гормон)
Пролактин считается одним из наиболее «древних» гормонов
Лактотропный гормон (пролактин, лютеотропный гормон)
Пролактин считается одним из наиболее «древних» гормонов

Лактотропный гормон (пролактин, лютеотропный гормон)
Расшифрована структура пролактина из гипофиза овцы, быка
Лактотропный гормон (пролактин, лютеотропный гормон)
Расшифрована структура пролактина из гипофиза овцы, быка

Тиреотропный гормон (ТТГ, тиротропин)
Тиротропин является сложным гликопротеином, выделяемым передней долей гипофиза и
Тиреотропный гормон (ТТГ, тиротропин)
Тиротропин является сложным гликопротеином, выделяемым передней долей гипофиза и

Гонадотропные гормоны (гонадотррпины)
К гонадотропинам относятся фолликулостимулирующий гормон (ФСГ, фоллитропин) и лютеинизирующий
Гонадотропные гормоны (гонадотррпины)
К гонадотропинам относятся фолликулостимулирующий гормон (ФСГ, фоллитропин) и лютеинизирующий

Липотропные гормоны (ЛТГ, липотропины)
Среди гормонов передней доли гипофиза следует отметить липотропины,
Липотропные гормоны (ЛТГ, липотропины)
Среди гормонов передней доли гипофиза следует отметить липотропины,

Липотропные гормоны (ЛТГ, липотропины)
Перечисленные биологические свойства обусловлены не β-липотропином, оказавшимся лишенным
Липотропные гормоны (ЛТГ, липотропины)
Перечисленные биологические свойства обусловлены не β-липотропином, оказавшимся лишенным

Липотропные гормоны (ЛТГ, липотропины)
Общим типом структуры для всех трех соединений является
Липотропные гормоны (ЛТГ, липотропины)
Общим типом структуры для всех трех соединений является

Липотропные гормоны (ЛТГ, липотропины)
Ниже представлены пептидные гормоны, образующиеся из β-липотропина путем
Липотропные гормоны (ЛТГ, липотропины)
Ниже представлены пептидные гормоны, образующиеся из β-липотропина путем

Липотропные гормоны (ЛТГ, липотропины)
Учитывая исключительную роль β-липотропина как предшественника перечисленных гормонов,
Липотропные гормоны (ЛТГ, липотропины)
Учитывая исключительную роль β-липотропина как предшественника перечисленных гормонов,

ГОРМОНЫ ПАРАЩИТОВИДНЫХ ЖЕЛЕЗ (ПАРАТГОРМОНЫ)
К гормонам белковой природы относится группа паратгормонов (паратиреоидных
ГОРМОНЫ ПАРАЩИТОВИДНЫХ ЖЕЛЕЗ (ПАРАТГОРМОНЫ)
К гормонам белковой природы относится группа паратгормонов (паратиреоидных

ГОРМОНЫ ПАРАЩИТОВИДНЫХ ЖЕЛЕЗ (ПАРАТГОРМОНЫ)
Паратгормон участвует в регуляции концентрации катионов кальция и
ГОРМОНЫ ПАРАЩИТОВИДНЫХ ЖЕЛЕЗ (ПАРАТГОРМОНЫ)
Паратгормон участвует в регуляции концентрации катионов кальция и

ГОРМОНЫ ПАРАЩИТОВИДНЫХ ЖЕЛЕЗ (ПАРАТГОРМОНЫ)
Другой орган-мишень паратгормона – это почка. Паратгормон уменьшает
ГОРМОНЫ ПАРАЩИТОВИДНЫХ ЖЕЛЕЗ (ПАРАТГОРМОНЫ)
Другой орган-мишень паратгормона – это почка. Паратгормон уменьшает
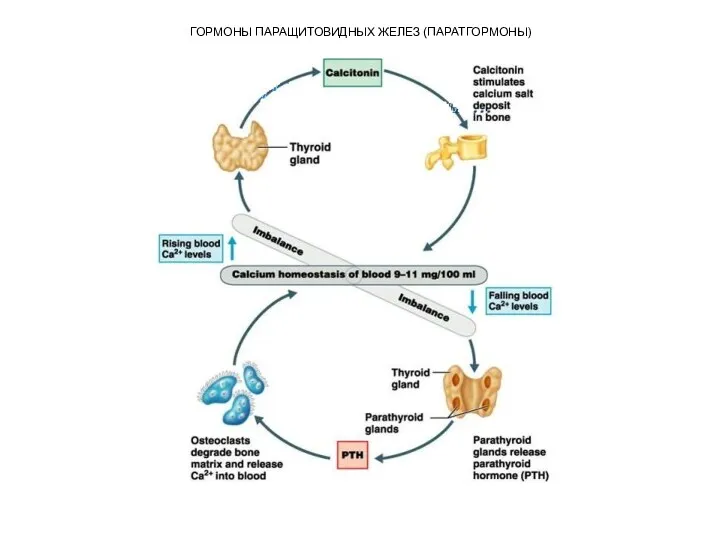
ГОРМОНЫ ПАРАЩИТОВИДНЫХ ЖЕЛЕЗ (ПАРАТГОРМОНЫ)
ГОРМОНЫ ПАРАЩИТОВИДНЫХ ЖЕЛЕЗ (ПАРАТГОРМОНЫ)

ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ
Щитовидная железа играет исключительно важную роль в обмене веществ.
ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ
Щитовидная железа играет исключительно важную роль в обмене веществ.

ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ
Впервые на существование кальцитонина, обладающего способностью поддерживать постоянный уровень
ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ
Впервые на существование кальцитонина, обладающего способностью поддерживать постоянный уровень

ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ
Кальцитонин человека содержит дисульфидный мостик (между 1-м и 7-м
ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ
Кальцитонин человека содержит дисульфидный мостик (между 1-м и 7-м
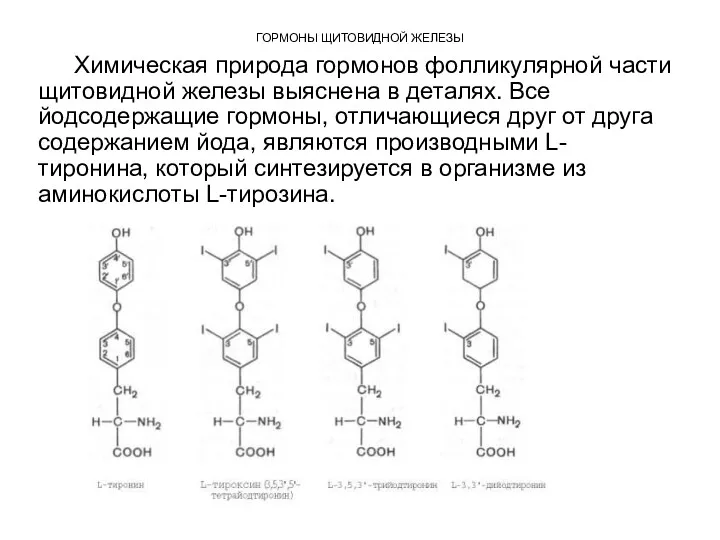
ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ
Химическая природа гормонов фолликулярной части щитовидной железы выяснена в
ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ
Химическая природа гормонов фолликулярной части щитовидной железы выяснена в

ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ
Из L-тиронина легко синтезируется гормон щитовидной железы тироксин,
ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ
Из L-тиронина легко синтезируется гормон щитовидной железы тироксин,

ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ
Катаболизм гормонов щитовидной железы протекает по двум направлениям: распад
ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ
Катаболизм гормонов щитовидной железы протекает по двум направлениям: распад

ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ
Гипофункция щитовидной железы в раннем детском возрасте приводит к
ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ
Гипофункция щитовидной железы в раннем детском возрасте приводит к

ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ
Следует отметить еще одно поражение щитовидной железы –
ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ
Следует отметить еще одно поражение щитовидной железы –
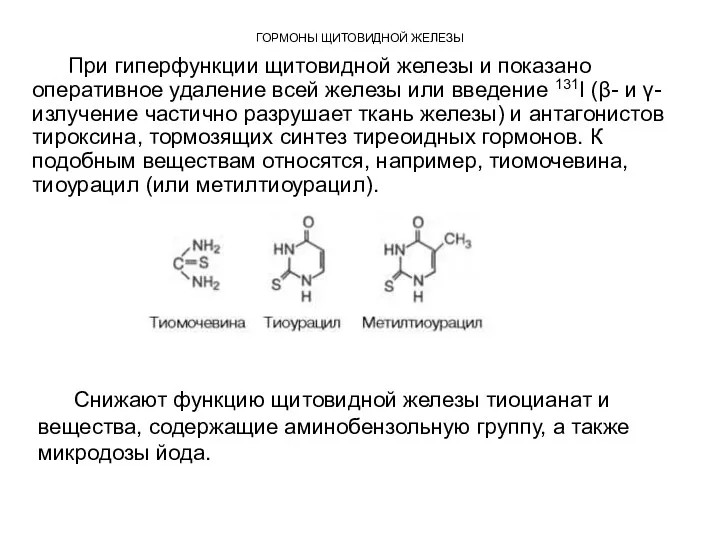
ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ
При гиперфункции щитовидной железы и показано оперативное удаление всей
ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ
При гиперфункции щитовидной железы и показано оперативное удаление всей

ГОРМОНЫ ПОДЖЕЛУДОЧНОЙ ЖЕЛЕЗЫ
Поджелудочная железа относится к железам со смешанной секрецией.
ГОРМОНЫ ПОДЖЕЛУДОЧНОЙ ЖЕЛЕЗЫ
Поджелудочная железа относится к железам со смешанной секрецией.

Инсулин
Инсулин получивший свое название от наименования панкреатических островков (лат. insula –
Инсулин
Инсулин получивший свое название от наименования панкреатических островков (лат. insula –

Инсулин
Биосинтез инсулина осуществляется в β-клетках панкреатических островков из своего предшественника проинсулина.
Инсулин
Биосинтез инсулина осуществляется в β-клетках панкреатических островков из своего предшественника проинсулина.

Инсулин
Синтезированный из проинсулина инсулин может существовать в нескольких формах, различающихся по
Инсулин
Синтезированный из проинсулина инсулин может существовать в нескольких формах, различающихся по

Инсулин
В физиологической регуляции синтеза инсулина доминирующую роль играет концентрация глюкозы в
Инсулин
В физиологической регуляции синтеза инсулина доминирующую роль играет концентрация глюкозы в

Инсулин
При недостаточной секреции (точнее, недостаточном синтезе) инсулина развивается специфическое заболевание –
Инсулин
При недостаточной секреции (точнее, недостаточном синтезе) инсулина развивается специфическое заболевание –
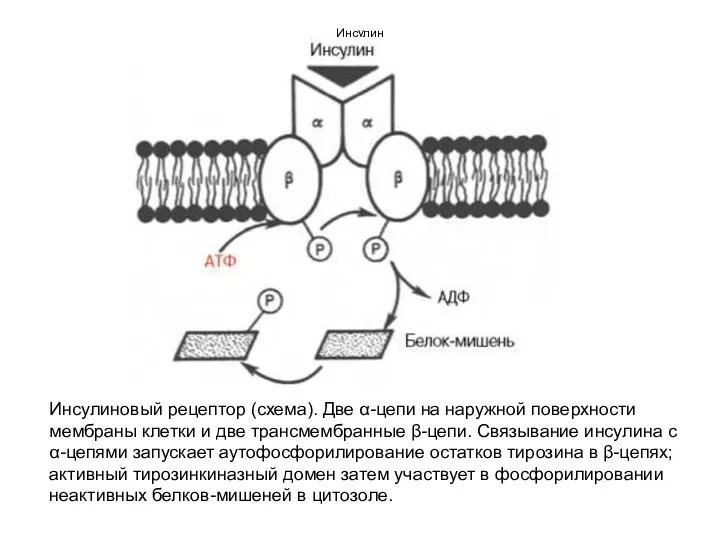
Инсулин
Инсулиновый рецептор (схема). Две α-цепи на наружной поверхности мембраны клетки и
Инсулин
Инсулиновый рецептор (схема). Две α-цепи на наружной поверхности мембраны клетки и

Инсулин
Наиболее вероятной в настоящее время представляется мембранная локализация первичного действия почти
Инсулин
Наиболее вероятной в настоящее время представляется мембранная локализация первичного действия почти

Глюкагон
Глюкагон впервые был обнаружен в коммерческих препаратах инсулина еще в 1923
Глюкагон
Глюкагон впервые был обнаружен в коммерческих препаратах инсулина еще в 1923

Глюкагон
По биологическому действию глюкагон относятся к гипергликемическим факторам, вызывает увеличение концентрации
Глюкагон
По биологическому действию глюкагон относятся к гипергликемическим факторам, вызывает увеличение концентрации

ГОРМОНЫ НАДПОЧЕЧНИКОВ
Надпочечники состоят из двух индивидуальных в морфологическом и функциональном отношениях
ГОРМОНЫ НАДПОЧЕЧНИКОВ
Надпочечники состоят из двух индивидуальных в морфологическом и функциональном отношениях
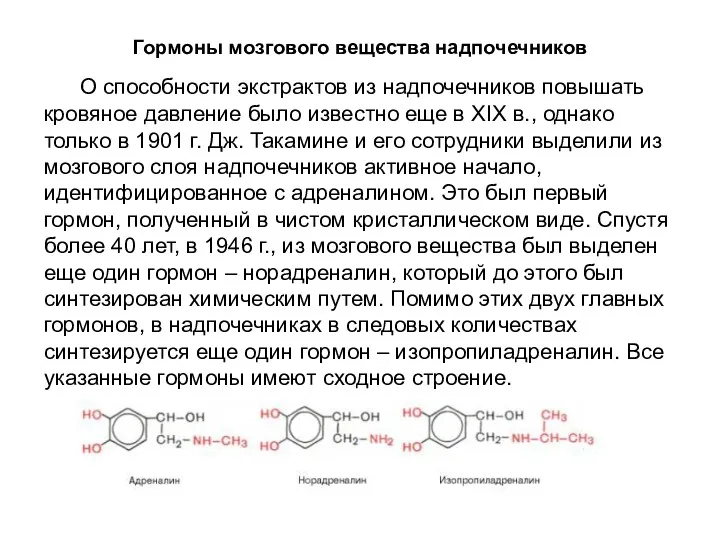
Гормоны мозгового вещества надпочечников
О способности экстрактов из надпочечников повышать кровяное давление
Гормоны мозгового вещества надпочечников
О способности экстрактов из надпочечников повышать кровяное давление
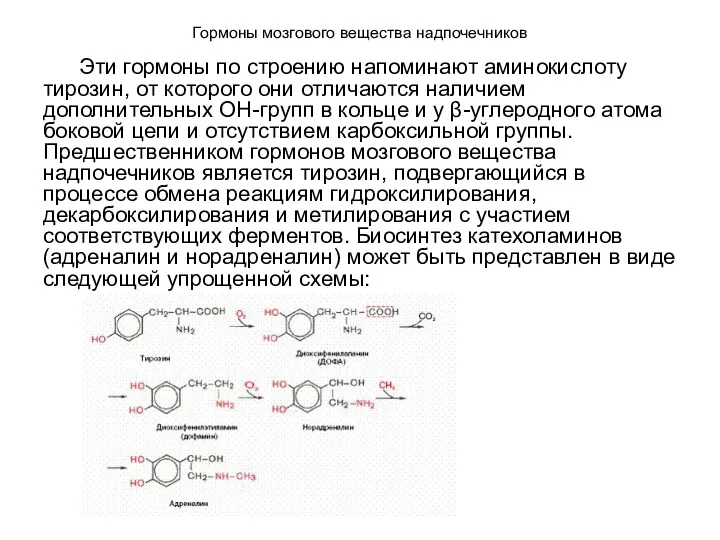
Гормоны мозгового вещества надпочечников
Эти гормоны по строению напоминают аминокислоту тирозин, от
Гормоны мозгового вещества надпочечников
Эти гормоны по строению напоминают аминокислоту тирозин, от
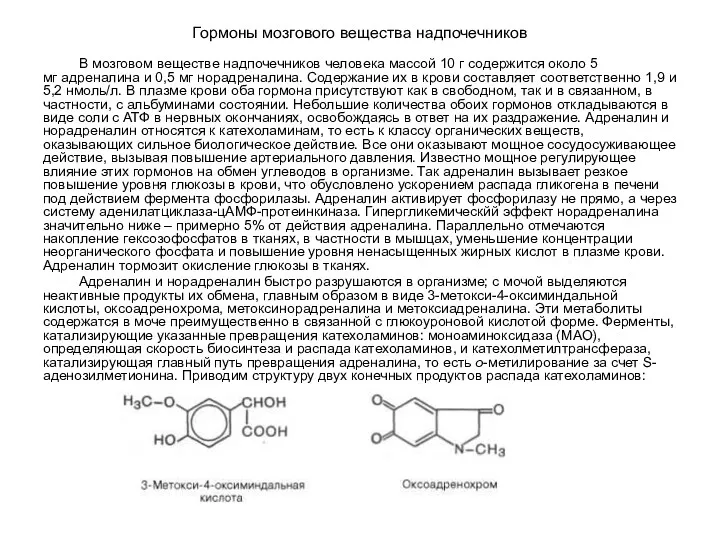
Гормоны мозгового вещества надпочечников
В мозговом веществе надпочечников человека массой 10 г
Гормоны мозгового вещества надпочечников
В мозговом веществе надпочечников человека массой 10 г

Гормоны коркового вещества надпочечников
Со второй половины XIX в. известно заболевание, названное
Гормоны коркового вещества надпочечников
Со второй половины XIX в. известно заболевание, названное

Химическое строение, биосинтез и биологическое действие кортикостероидов
К настоящему времени из коркового
Химическое строение, биосинтез и биологическое действие кортикостероидов
К настоящему времени из коркового
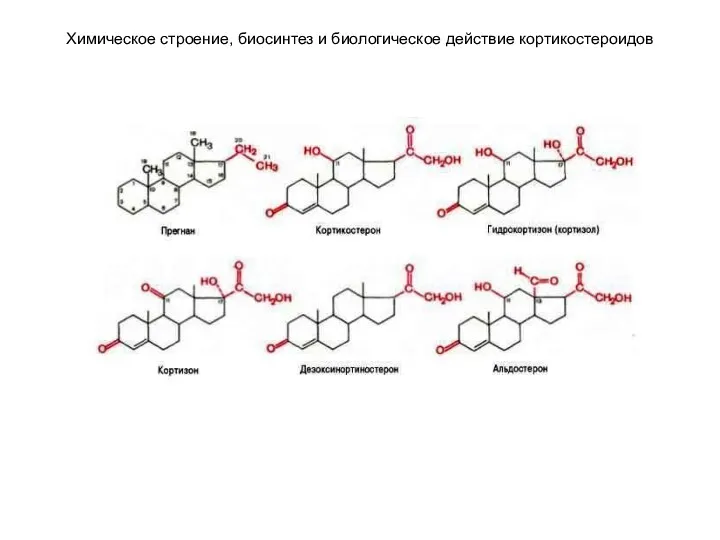
Химическое строение, биосинтез и биологическое действие кортикостероидов
Химическое строение, биосинтез и биологическое действие кортикостероидов

Химическое строение, биосинтез и биологическое действие кортикостероидов
Предшественником кортикостероидов является холестерин(ол). В
Химическое строение, биосинтез и биологическое действие кортикостероидов
Предшественником кортикостероидов является холестерин(ол). В

Химическое строение, биосинтез и биологическое действие кортикостероидов
Ферменты катализируют минимум две последовательные
Химическое строение, биосинтез и биологическое действие кортикостероидов
Ферменты катализируют минимум две последовательные

Химическое строение, биосинтез и биологическое действие кортикостероидов
Минералокортикоиды (дезоксикортикостерон и альдостерон) регулируют главным
Химическое строение, биосинтез и биологическое действие кортикостероидов
Минералокортикоиды (дезоксикортикостерон и альдостерон) регулируют главным
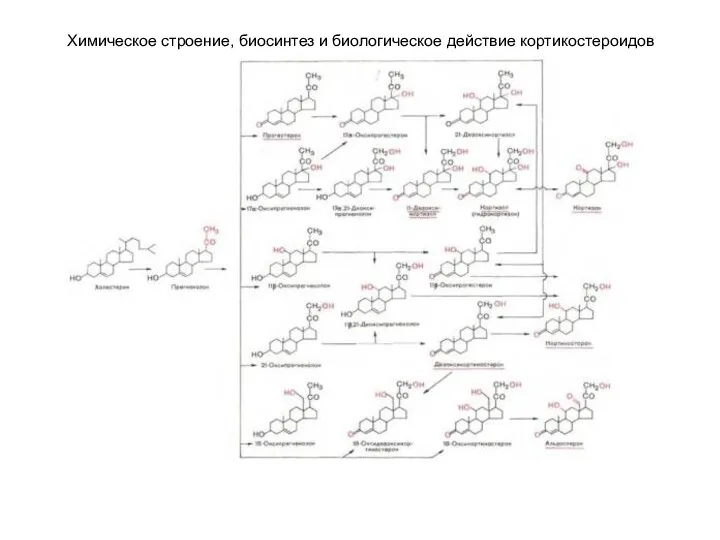
Химическое строение, биосинтез и биологическое действие кортикостероидов
Химическое строение, биосинтез и биологическое действие кортикостероидов

ПОЛОВЫЕ ГОРМОНЫ
Половые гормоны синтезируются в основном в половых железах женщин (яичники)
ПОЛОВЫЕ ГОРМОНЫ
Половые гормоны синтезируются в основном в половых железах женщин (яичники)
Женские половые гормоны
Основным местом синтеза женских половых гормонов – эстрогенов (от
Женские половые гормоны
Основным местом синтеза женских половых гормонов – эстрогенов (от

Женские половые гормоны
Наиболее активный эстроген – эстрадиол, синтезируется преимущественно в фолликулах;
Женские половые гормоны
Наиболее активный эстроген – эстрадиол, синтезируется преимущественно в фолликулах;
Женские половые гормоны
Ведущую роль в регуляции синтеза эстрогенов и прогестерона играют
Женские половые гормоны
Ведущую роль в регуляции синтеза эстрогенов и прогестерона играют

Мужские половые гормоны
Внутрисекреторная функция мужских половых желез была установлена в 1849 г.,
Мужские половые гормоны
Внутрисекреторная функция мужских половых желез была установлена в 1849 г.,

Мужские половые гормоны
Биосинтез андрогенов осуществляется главным образом в семенниках и частично
Мужские половые гормоны
Биосинтез андрогенов осуществляется главным образом в семенниках и частично

ПРОСТАГЛАНДИНЫ
Термин «простагландины» был введен У. Эйлером, впервые показавшим, что в сперме человека
ПРОСТАГЛАНДИНЫ
Термин «простагландины» был введен У. Эйлером, впервые показавшим, что в сперме человека
ПРОСТАГЛАНДИНЫ
ПРОСТАГЛАНДИНЫ

ПРОСТАГЛАНДИНЫ
Первый путь получил наименование циклооксигеназного пути превращения арахидоновой кислоты, поскольку первые
ПРОСТАГЛАНДИНЫ
Первый путь получил наименование циклооксигеназного пути превращения арахидоновой кислоты, поскольку первые
ПРОСТАГЛАНДИНЫ
ПРОСТАГЛАНДИНЫ

ПРОСТАГЛАНДИНЫ
Второй путь превращения арахидоновой кислоты – липоксигеназный путь – отличается тем, что
ПРОСТАГЛАНДИНЫ
Второй путь превращения арахидоновой кислоты – липоксигеназный путь – отличается тем, что

ГОРМОНЫ ВИЛОЧКОВОЙ ЖЕЛЕЗЫ (ТИМУСА)
Роль тимуса как эндокринной железы известна давно. Известно, что
ГОРМОНЫ ВИЛОЧКОВОЙ ЖЕЛЕЗЫ (ТИМУСА)
Роль тимуса как эндокринной железы известна давно. Известно, что

Все известные к настоящему времени гормональные вещества описать почти не возможно, их
Все известные к настоящему времени гормональные вещества описать почти не возможно, их

МОЛЕКУЛЯРНЫЕ МЕХАНИЗМЫ ПЕРЕДАЧИ ГОРМОНАЛЬНОГО СИГНАЛА
Несмотря на огромное разнообразие гормонов и гормоноподобных веществ,
МОЛЕКУЛЯРНЫЕ МЕХАНИЗМЫ ПЕРЕДАЧИ ГОРМОНАЛЬНОГО СИГНАЛА
Несмотря на огромное разнообразие гормонов и гормоноподобных веществ,

МОЛЕКУЛЯРНЫЕ МЕХАНИЗМЫ ПЕРЕДАЧИ ГОРМОНАЛЬНОГО СИГНАЛА
Общим фундаментальным механизмом, посредством которого реализуются биологические
МОЛЕКУЛЯРНЫЕ МЕХАНИЗМЫ ПЕРЕДАЧИ ГОРМОНАЛЬНОГО СИГНАЛА
Общим фундаментальным механизмом, посредством которого реализуются биологические

Аденилатциклазная мессенджерная система
Наиболее изученным является аденилатциклазный путь передачи гормонального сигнала. В
Аденилатциклазная мессенджерная система
Наиболее изученным является аденилатциклазный путь передачи гормонального сигнала. В
Аденилатциклазная мессенджерная система
Рис. Аденилатциклазный путь передачи гормонального сигнала Рец - рецептор;
Аденилатциклазная мессенджерная система
Рис. Аденилатциклазный путь передачи гормонального сигнала Рец - рецептор;
Аденилатциклазная мессенджерная система
Каталитический компонент аденилатциклазы в отсутствие G-белков практически неактивен, он
Аденилатциклазная мессенджерная система
Каталитический компонент аденилатциклазы в отсутствие G-белков практически неактивен, он
Аденилатциклазная мессенджерная система
Протеинкиназа – это внутриклеточный фермент, через который цАМФ реализует
Аденилатциклазная мессенджерная система
Протеинкиназа – это внутриклеточный фермент, через который цАМФ реализует

Аденилатциклазная мессенджерная система
В клетках открыт большой класс цАМФ-зависимых протеинкиназ, названных протеинкиназами А;
Аденилатциклазная мессенджерная система
В клетках открыт большой класс цАМФ-зависимых протеинкиназ, названных протеинкиназами А;
Аденилатциклазная мессенджерная система
Второй тип – аллостерическая регуляция. Она основана на реакциях аденилирования
Аденилатциклазная мессенджерная система
Второй тип – аллостерическая регуляция. Она основана на реакциях аденилирования

Гуанилатциклазная мессенджерная система
Довольно долгое время циклический гуанозинмонофосфат (цГМФ) рассматривался как антипод
Гуанилатциклазная мессенджерная система
Довольно долгое время циклический гуанозинмонофосфат (цГМФ) рассматривался как антипод
Гуанилатциклазная мессенджерная система
Рис. Схематическое выражение центральной роли цАМФ и протеинкиназы в гормональной регуляции
Гуанилатциклазная мессенджерная система
Рис. Схематическое выражение центральной роли цАМФ и протеинкиназы в гормональной регуляции

Гуанилатциклазная мессенджерная система
Известны четыре разные формы гуанилатциклазы, три из которых являются
Гуанилатциклазная мессенджерная система
Известны четыре разные формы гуанилатциклазы, три из которых являются
Гуанилатциклазная мессенджерная система
Растворимая форма гуанилатциклазы является гемсодержащим ферментом, состоящим из 2
Гуанилатциклазная мессенджерная система
Растворимая форма гуанилатциклазы является гемсодержащим ферментом, состоящим из 2

Гуанилатциклазная мессенджерная система
Оксид азота при взаимодействии с гемом гуанилатциклазы способствует быстрому образованию
Гуанилатциклазная мессенджерная система
Оксид азота при взаимодействии с гемом гуанилатциклазы способствует быстрому образованию

Са2+-мессенджерная система
Ионам Са2+ принадлежит центральная роль в регуляции многих клеточных функций. Изменение
Са2+-мессенджерная система
Ионам Са2+ принадлежит центральная роль в регуляции многих клеточных функций. Изменение

Са2+-мессенджерная система
К внутриклеточной системе мессенджеров относят также производные фосфолипидов мембран эукариотических
Са2+-мессенджерная система
К внутриклеточной системе мессенджеров относят также производные фосфолипидов мембран эукариотических
Са2+-мессенджерная система
Са2+-мессенджерная система
 Строение, свойства костей типы их соединения
Строение, свойства костей типы их соединения Презентации
Презентации Общая характеристика Типа Хордовые
Общая характеристика Типа Хордовые Применение грибов
Применение грибов Влияние витаминов на здоровье человека
Влияние витаминов на здоровье человека Соединительная ткань
Соединительная ткань Эфирные масла-афродизиаки
Эфирные масла-афродизиаки Ферменты. Классификация ферментов по структуре
Ферменты. Классификация ферментов по структуре Что такое метаболизм и как его измерить
Что такое метаболизм и как его измерить Выращивание томатов
Выращивание томатов Углеводы. Строение и функции
Углеводы. Строение и функции Анатомическое строение гортани человека
Анатомическое строение гортани человека Физиология вегетативной нервной системы (ВНС)
Физиология вегетативной нервной системы (ВНС) Биология культурных растений
Биология культурных растений Класс Трематоды. Трематодозы
Класс Трематоды. Трематодозы Скелет и мышцы
Скелет и мышцы Вклад учёных в развитие биологии
Вклад учёных в развитие биологии Зиянкестерге қарсы микробты, вирусты препараттар және антибиотиктер. (Лекция 11)
Зиянкестерге қарсы микробты, вирусты препараттар және антибиотиктер. (Лекция 11) Покриви тіла тварин
Покриви тіла тварин Алгоритм решения задач на транскрипцию и трансляцию
Алгоритм решения задач на транскрипцию и трансляцию Насекомые. Игра Как стать миллионером
Насекомые. Игра Как стать миллионером Тип Членистоногі. Різноманітність представників класу Павукоподібні
Тип Членистоногі. Різноманітність представників класу Павукоподібні Анализаторы. Рецепторы. Виды чувствительности
Анализаторы. Рецепторы. Виды чувствительности Вода, её значение и свойства
Вода, её значение и свойства Животноводство. Из каких отраслей состоит животноводство?
Животноводство. Из каких отраслей состоит животноводство? Круговорот веществ в природе
Круговорот веществ в природе Вегетативная нервная система в норме и патологии
Вегетативная нервная система в норме и патологии Тип хордовые животные
Тип хордовые животные