Химический синапс. Жизненный цикл медиатора: синтез, выброс в синаптическую щель, взаимодействие с рецепторами, инактивация презентация
- Химический синапс. Жизненный цикл медиатора: синтез, выброс в синаптическую щель, взаимодействие с рецепторами, инактивация

Содержание
- 2. хими- ческий синапс: передача сигнала идет за счет вы- деления медиа- тора 1 – пресинаптическое окончание
- 3. Синапсы бывают: центральные; нервно-мышечные; вегетативные и др. Центральные синапсы: чаще аксо-дендритные и аксо-соматические; реже дендро-дендритные, аксо-аксональные
- 4. Основные стадии передачи сигнала в синапсе: 1. ПД запускает движение везикул и выброс медиа- тора в
- 5. Главное «действующее лицо» в синаптической передаче – медиатор. синтез и накопление в пресинаптическом окончании; выброс в
- 6. синтез и накопление в пресинаптическом окончании; выброс в синаптическую щель при появлении ПД; действие на рецепторы
- 7. Если синтез идет в соме, то далее: (а) ЭПС переносит медиатор в комплекс Гольджи; (б) комплекс
- 8. Эволюционно это более древний вариант; есть риск остаться без медиатора в случае интенсивной работы синапса (перенос
- 9. Выброс (экзоцитоз) медиатора в синаптическую щель происходит после появления ПД, который вызывает открывание электрочувствительных Са2+-каналов (примерно
- 10. Фото, полу- ченное при помощи электронного микроскопа масштаб: 500 мкм синаптическая щель Везикула «разби-вается, как яйцо»,
- 11. Несколько дополнений: Увеличение концентрации Са2+ в межклеточной среде ведет к его более активному входу в пресинаптическое
- 12. Бактерия ботулизма – почвен-ная, анаэробная (не выносит О2). Ее токсин блокирует белки, отвечающие за экзоцитоз; отравление
- 13. Еще раз о последовательности событий, происходящих в синапсе: (1) распространение ПД; (2-4) вход ионов Са2+ и
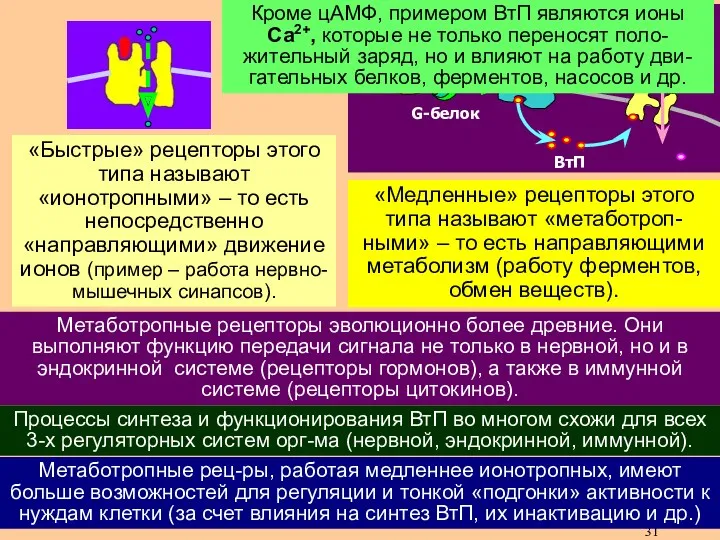
- 14. На следующем этапе ВтП запускает открывание ионных каналов для Na+, K+ либо Cl- (хемочувствительные ионные каналы).
- 15. Рассмотрим теперь, как откры- вание хемочувствительных Na+-каналов приводит к возбуждению постсинаптичес- кой мембраны и, в конечном
- 16. Рассмотрим теперь, как откры- вание хемочувствительных Na+-каналов приводит к возбуждению постсинаптичес- кой мембраны и, в конечном
- 17. ПД может быть за- пущен повторной стимуляцией одного и того же синапса («временнáя суммация»). Запуск ПД
- 18. Кроме «временнóй» выделяют также пространственную суммацию. В этом случае накладываются друг на друга ВПСП, обусловленные одновременным
- 19. Синапсы, запускающие ВПСП, называются «возбуждающими»; действующие в них медиаторы – «возбуждающими медиаторами». Однако, кроме этого, существуют
- 20. В этой точке мембраны нейрона произойдет запуск ПД, если ∑ всех ВПСП – ∑ всех ТПСП
- 21. Вернемся к ТПСП. Открывание не только К+-каналов, но и Cl--каналов ведет к развитию торможения. Ионов Cl-
- 22. Тем не менее, такой вход Cl- (как и «обычный» ТПСП) эффективно про-тиводействует запуску ПД. Итак, перечислим
- 23. Итак, перечислим основные функции мембраны постсинаптической клетки. Здесь – «лоскутное одеяло»: области с (1) рецеп- торами,
- 24. Итак, перечислим основные функции мембраны постсинаптической клетки. Здесь – «лоскутное одеяло»: области с (1) рецеп- торами,
- 25. Таким образом, в синапсе электрический сигнал (ПД аксона) сначала превраща-ется в химический (движение медиатора и вторичного
- 26. Инактивация – это процесс удаления медиатора с рецептора для предотвращения слишком длительной (сильной) передачи сигнала. Осталось
- 27. В каждом конкретном синапсе используется один трех путей инактивации: 1) разрушение медиатора с помощью фермента; 2)
- 28. В каждом конкретном синапсе используется один трех путей инактивации: 1) разрушение медиатора с помощью фермента; 2)
- 29. Совместная деятельность систем инактивации медиатора и ВтП ограничивает время открывания хемочувствитель-ных каналов и, в итоге, длительность
- 30. Этот способ превращения передаваемого медиатором сигнала в ВПСП или ТПСП не единственный. Для ускорения процесса эволюция
- 31. Для ускорения процесса эволюция отыскала прямой путь: «гибрид» [рецептор + ионный канал] – единая супермолекула, имеющая
- 32. Антагонист: вещество,противо- действующее эффектам медиато-ра. Молекула состоит защитной части и неполной ключевой части. Последняя из них
- 33. Знания о медиаторах, их функциях, свойствах аго- нистов и антагонистов – важнейший раздел физи- ологии ЦНС,
- 34. И наконец… Еще раз подчеркну, что именно синапс (а не нейрон) является элементарной функциональ- ной единицей
- 36. Скачать презентацию
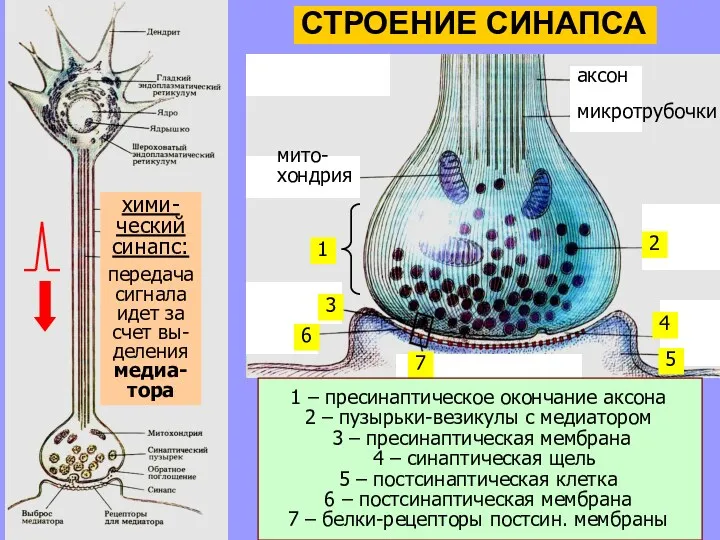
хими-
ческий
синапс:
передача
сигнала
идет за
счет вы-
деления
медиа-
тора
1 – пресинаптическое окончание аксона
2 – пузырьки-везикулы с медиатором
хими-
ческий
синапс:
передача
сигнала
идет за
счет вы-
деления
медиа-
тора
1 – пресинаптическое окончание аксона
2 – пузырьки-везикулы с медиатором
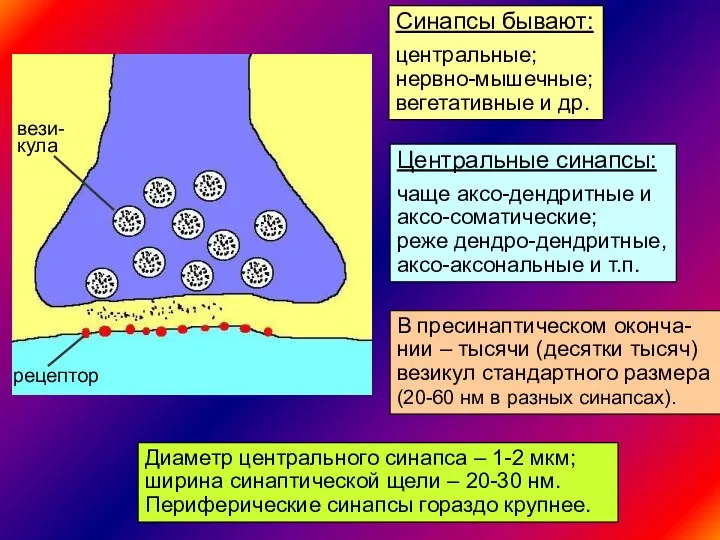
Синапсы бывают:
центральные;
нервно-мышечные;
вегетативные и др.
Центральные синапсы:
чаще аксо-дендритные и
аксо-соматические;
реже дендро-дендритные,
аксо-аксональные и т.п.
вези-
кула
рецептор
В пресинаптическом
Синапсы бывают:
центральные;
нервно-мышечные;
вегетативные и др.
Центральные синапсы:
чаще аксо-дендритные и
аксо-соматические;
реже дендро-дендритные,
аксо-аксональные и т.п.
вези-
кула
рецептор
В пресинаптическом
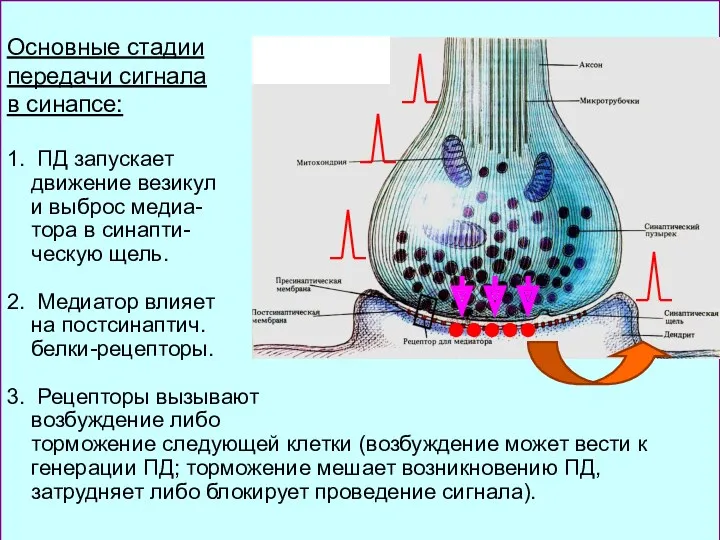
Основные стадии
передачи сигнала
в синапсе:
1. ПД запускает
движение везикул
передачи сигнала
в синапсе:
1. ПД запускает
движение везикул
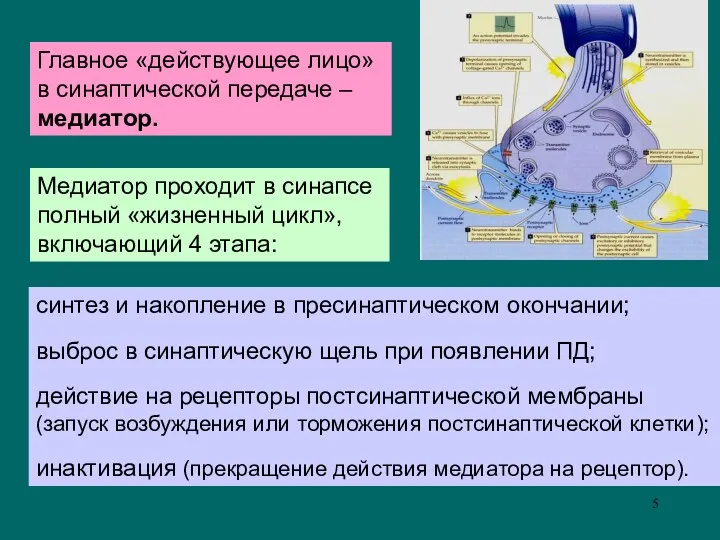
Главное «действующее лицо»
в синаптической передаче –
медиатор.
синтез и накопление в
Главное «действующее лицо»
в синаптической передаче –
медиатор.
синтез и накопление в
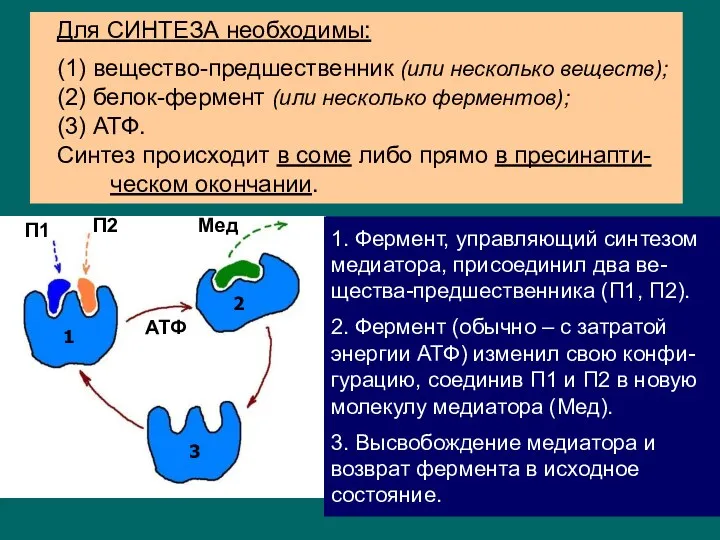
синтез и накопление в пресинаптическом окончании;
выброс в синаптическую щель при появлении
синтез и накопление в пресинаптическом окончании;
выброс в синаптическую щель при появлении
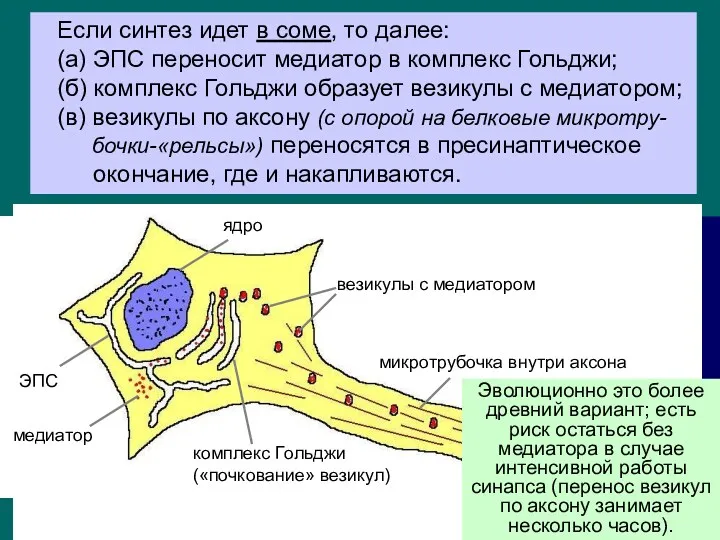
Если синтез идет в соме, то далее:
(а) ЭПС переносит
Если синтез идет в соме, то далее:
(а) ЭПС переносит
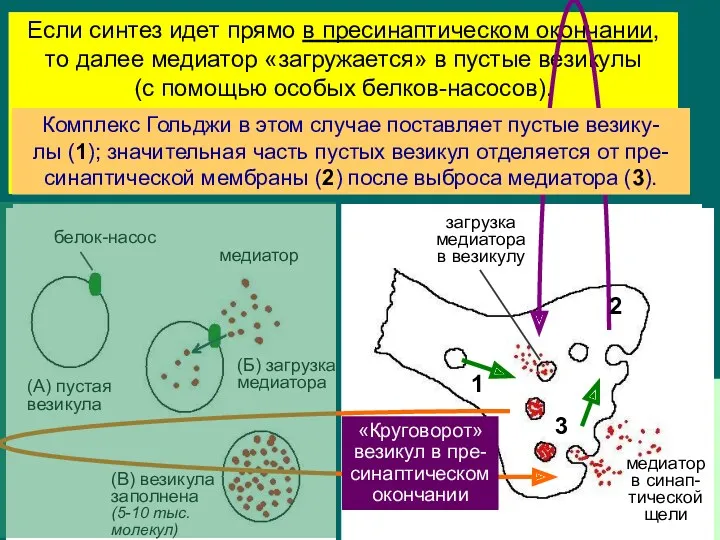
Эволюционно это более древний вариант; есть риск остаться без медиатора в
Эволюционно это более древний вариант; есть риск остаться без медиатора в
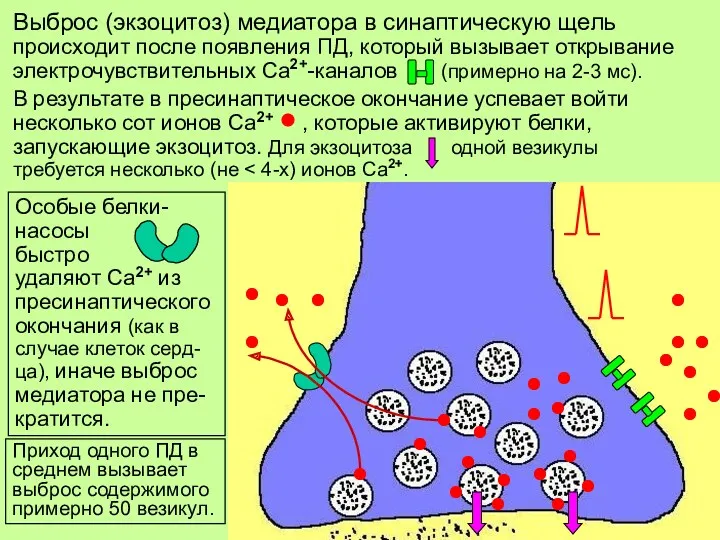
Выброс (экзоцитоз) медиатора в синаптическую щель
происходит после появления ПД, который
Выброс (экзоцитоз) медиатора в синаптическую щель
происходит после появления ПД, который
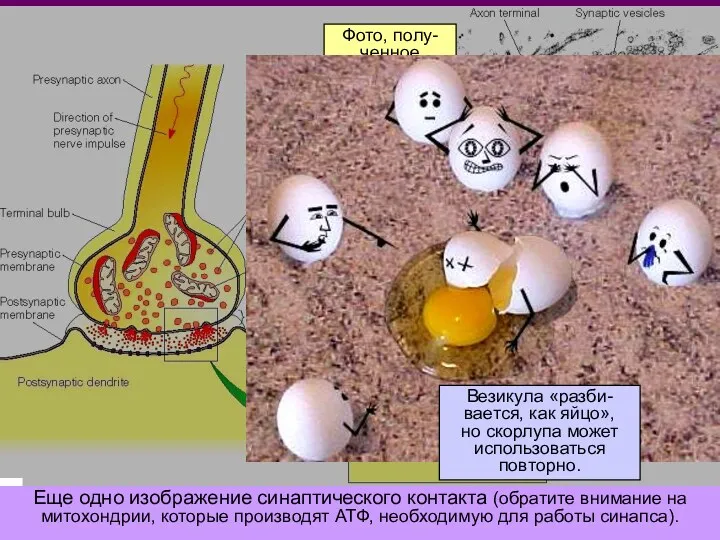
Фото, полу-
ченное
при помощи
электронного
микроскопа
масштаб:
500 мкм
синаптическая
щель
Везикула «разби-вается, как яйцо»,
но скорлупа может использоваться
Фото, полу-
ченное
при помощи
электронного
микроскопа
масштаб:
500 мкм
синаптическая
щель
Везикула «разби-вается, как яйцо»,
но скорлупа может использоваться
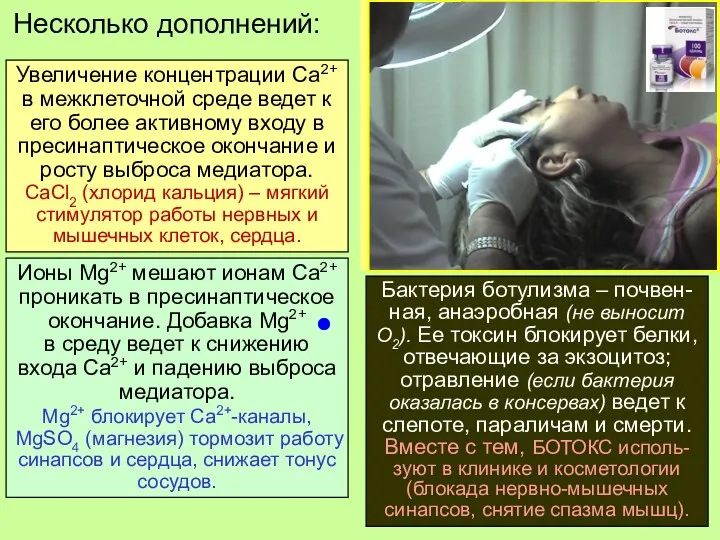
Несколько дополнений:
Увеличение концентрации Са2+ в межклеточной среде ведет к его более
Несколько дополнений:
Увеличение концентрации Са2+ в межклеточной среде ведет к его более

Бактерия ботулизма – почвен-ная, анаэробная (не выносит О2). Ее токсин блокирует
Бактерия ботулизма – почвен-ная, анаэробная (не выносит О2). Ее токсин блокирует
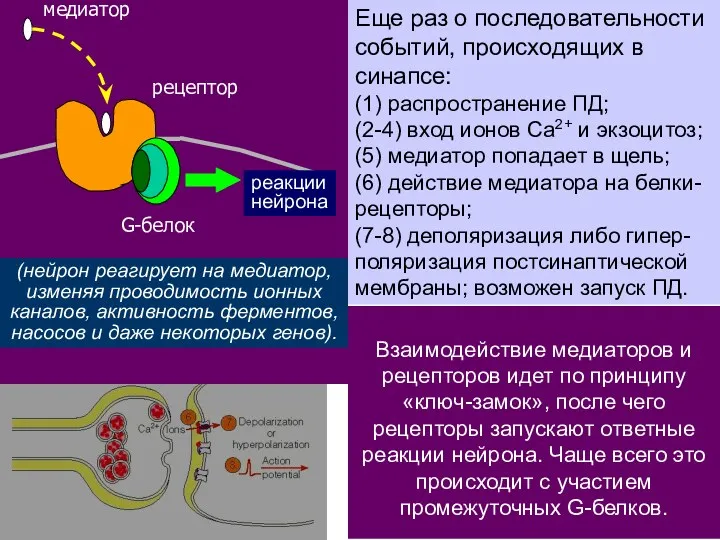
Еще раз о последовательности событий, происходящих в синапсе:
(1) распространение ПД;
(2-4) вход
Еще раз о последовательности событий, происходящих в синапсе:
(1) распространение ПД;
(2-4) вход
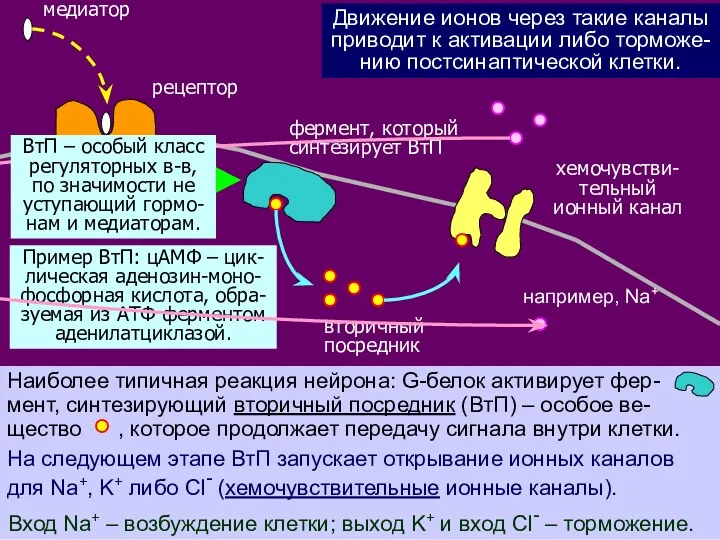
На следующем этапе ВтП запускает открывание ионных каналов
для Na+, K+
На следующем этапе ВтП запускает открывание ионных каналов
для Na+, K+
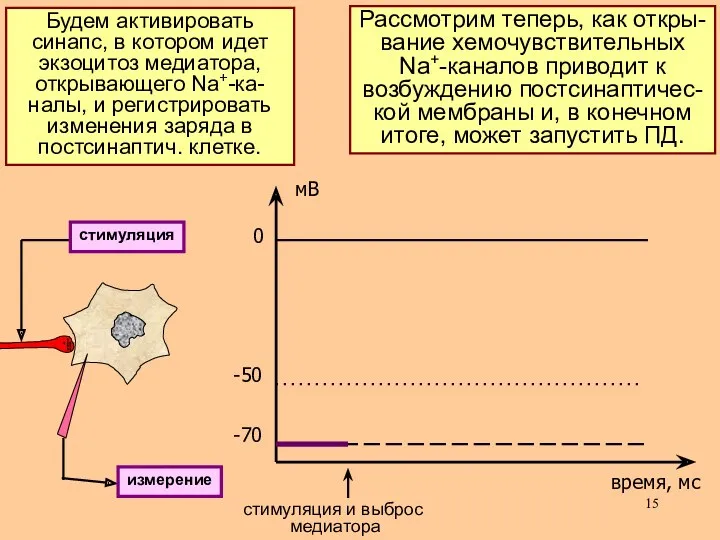
Рассмотрим теперь, как откры-
вание хемочувствительных
Na+-каналов приводит к
возбуждению постсинаптичес-
кой мембраны
Рассмотрим теперь, как откры-
вание хемочувствительных
Na+-каналов приводит к
возбуждению постсинаптичес-
кой мембраны
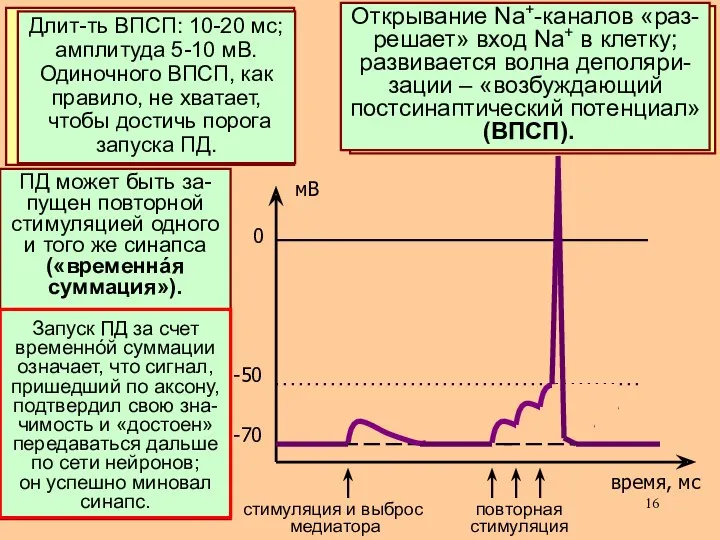
Рассмотрим теперь, как откры-
вание хемочувствительных
Na+-каналов приводит к
возбуждению постсинаптичес-
кой мембраны
Рассмотрим теперь, как откры-
вание хемочувствительных
Na+-каналов приводит к
возбуждению постсинаптичес-
кой мембраны
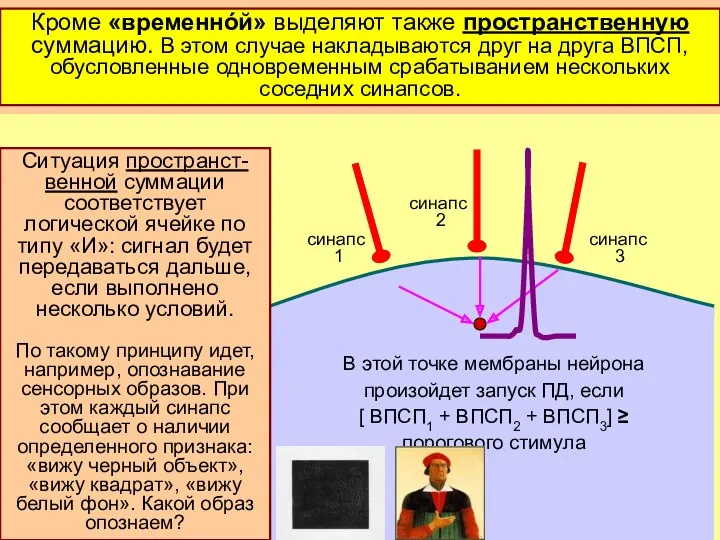
ПД может быть за-
пущен повторной
стимуляцией одного
и того же синапса
(«временнáя
ПД может быть за-
пущен повторной
стимуляцией одного
и того же синапса
(«временнáя
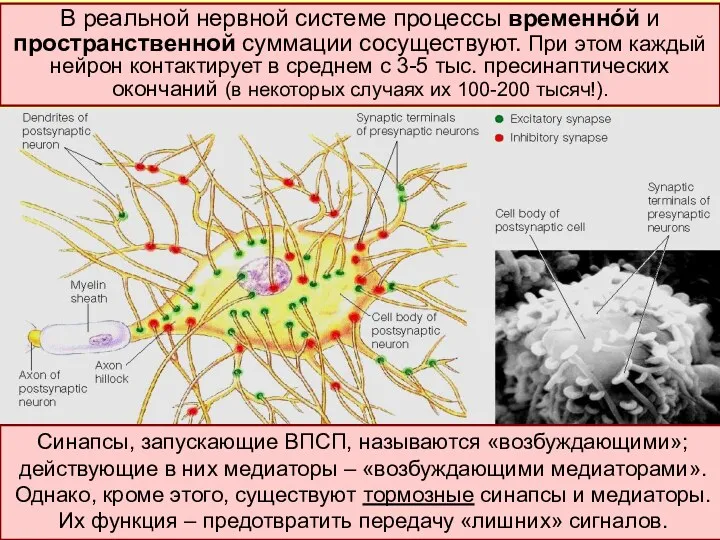
Кроме «временнóй» выделяют также пространственную суммацию. В этом случае накладываются друг
Кроме «временнóй» выделяют также пространственную суммацию. В этом случае накладываются друг
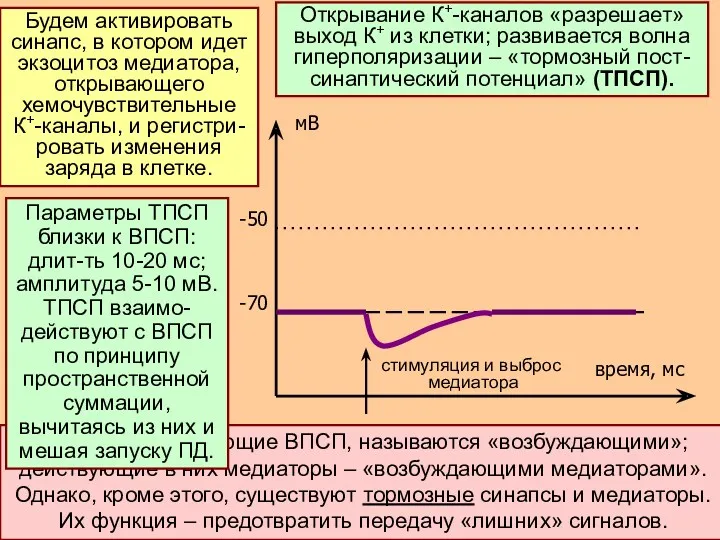
Синапсы, запускающие ВПСП, называются «возбуждающими»; действующие в них медиаторы – «возбуждающими
Синапсы, запускающие ВПСП, называются «возбуждающими»; действующие в них медиаторы – «возбуждающими
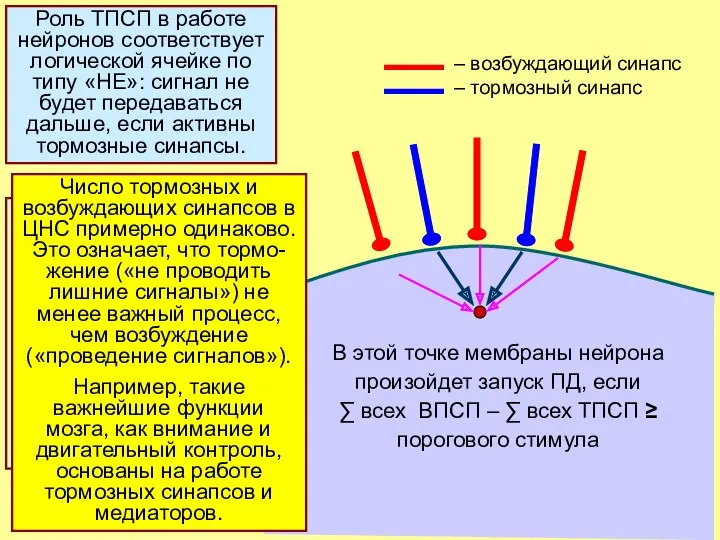
В этой точке мембраны нейрона
произойдет запуск ПД, если
∑ всех ВПСП –
В этой точке мембраны нейрона
произойдет запуск ПД, если
∑ всех ВПСП –
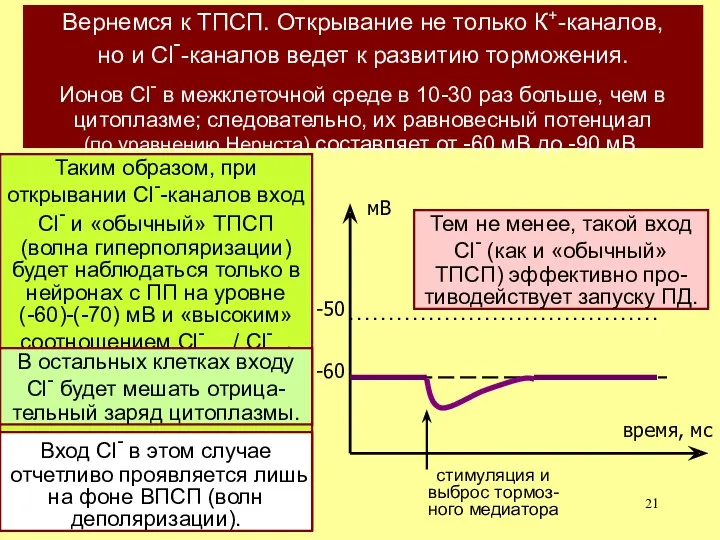
Вернемся к ТПСП. Открывание не только К+-каналов,
но и Cl--каналов ведет
Вернемся к ТПСП. Открывание не только К+-каналов,
но и Cl--каналов ведет

Тем не менее, такой вход Cl- (как и «обычный» ТПСП) эффективно
Тем не менее, такой вход Cl- (как и «обычный» ТПСП) эффективно
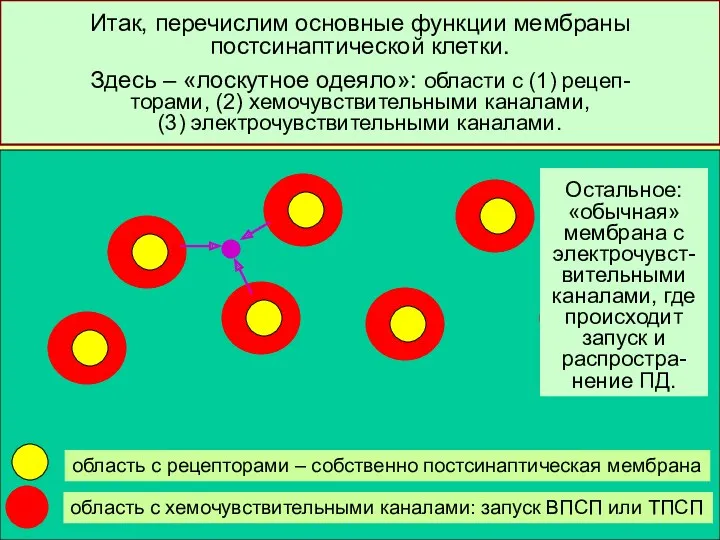
Итак, перечислим основные функции мембраны постсинаптической клетки.
Здесь – «лоскутное одеяло»:
Здесь – «лоскутное одеяло»:
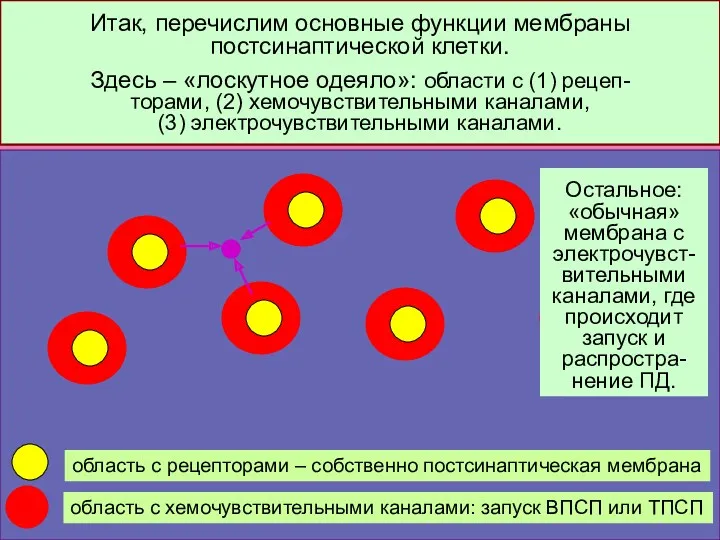
Итак, перечислим основные функции мембраны постсинаптической клетки.
Здесь – «лоскутное одеяло»:
Здесь – «лоскутное одеяло»:
Таким образом, в синапсе электрический сигнал (ПД аксона) сначала превраща-ется в
Таким образом, в синапсе электрический сигнал (ПД аксона) сначала превраща-ется в

Инактивация – это процесс удаления медиатора с рецептора для предотвращения слишком
Инактивация – это процесс удаления медиатора с рецептора для предотвращения слишком
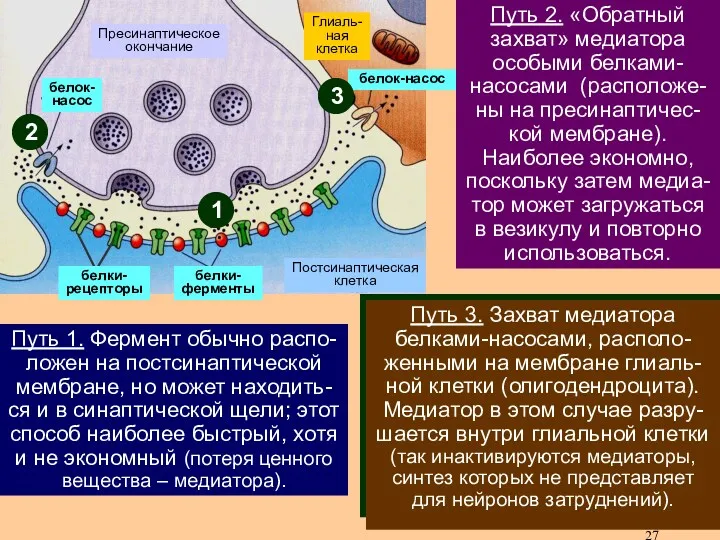
В каждом конкретном синапсе используется один трех путей инактивации:
1) разрушение медиатора
В каждом конкретном синапсе используется один трех путей инактивации:
1) разрушение медиатора
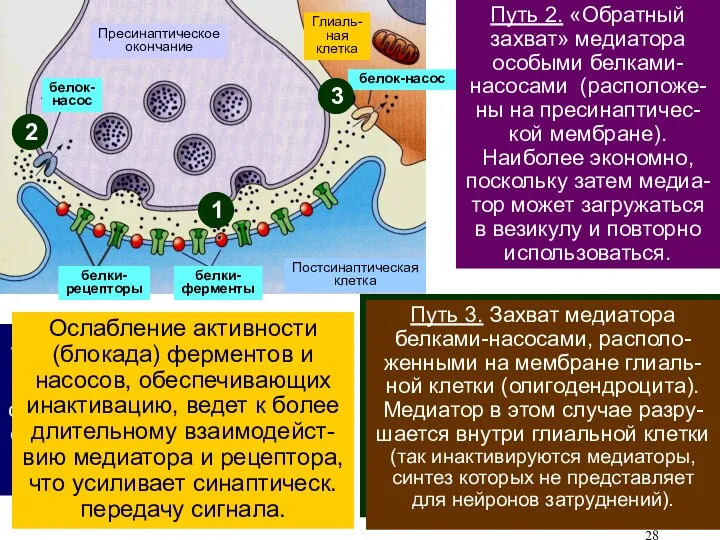
В каждом конкретном синапсе используется один трех путей инактивации:
1) разрушение медиатора
В каждом конкретном синапсе используется один трех путей инактивации:
1) разрушение медиатора
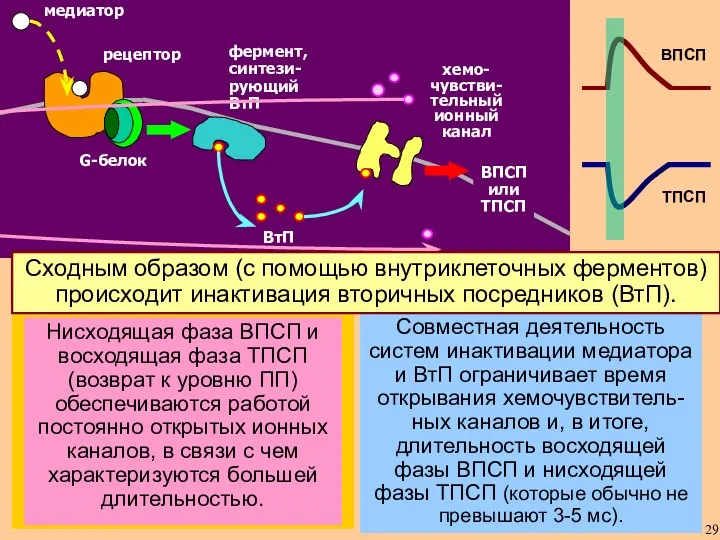
Совместная деятельность систем инактивации медиатора и ВтП ограничивает время открывания хемочувствитель-ных
Совместная деятельность систем инактивации медиатора и ВтП ограничивает время открывания хемочувствитель-ных
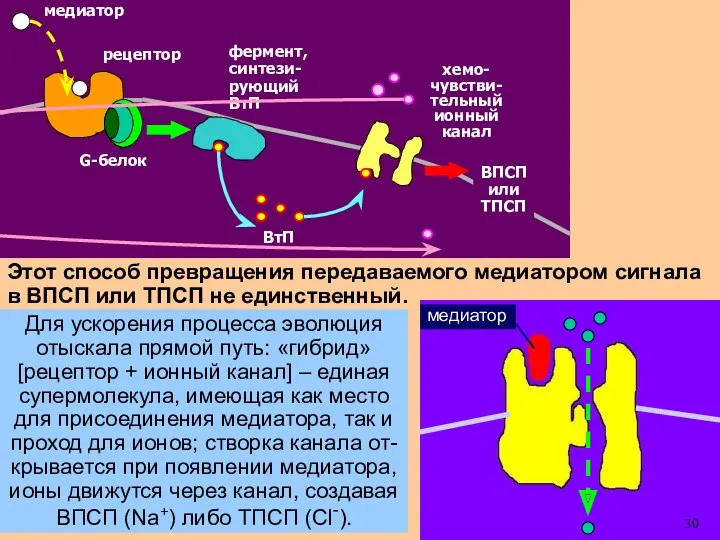
Этот способ превращения передаваемого медиатором сигнала в ВПСП или ТПСП не
Этот способ превращения передаваемого медиатором сигнала в ВПСП или ТПСП не
Для ускорения процесса эволюция отыскала прямой путь: «гибрид» [рецептор + ионный
Для ускорения процесса эволюция отыскала прямой путь: «гибрид» [рецептор + ионный
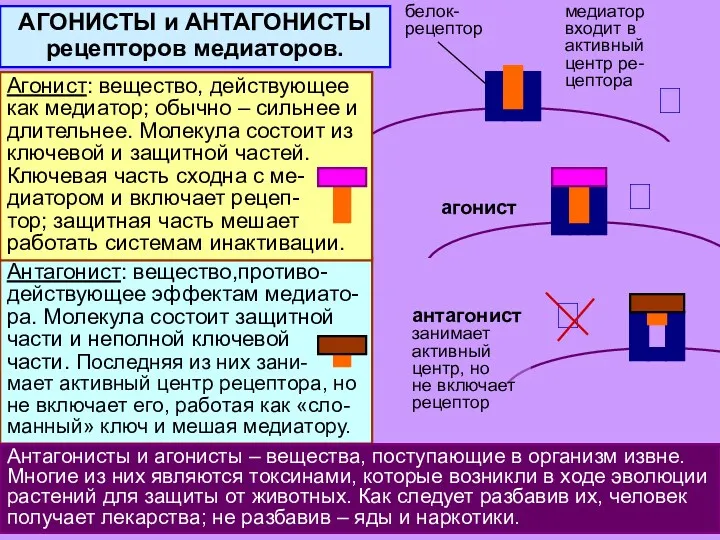
Антагонист: вещество,противо- действующее эффектам медиато-ра. Молекула состоит защитной части и неполной
Антагонист: вещество,противо- действующее эффектам медиато-ра. Молекула состоит защитной части и неполной
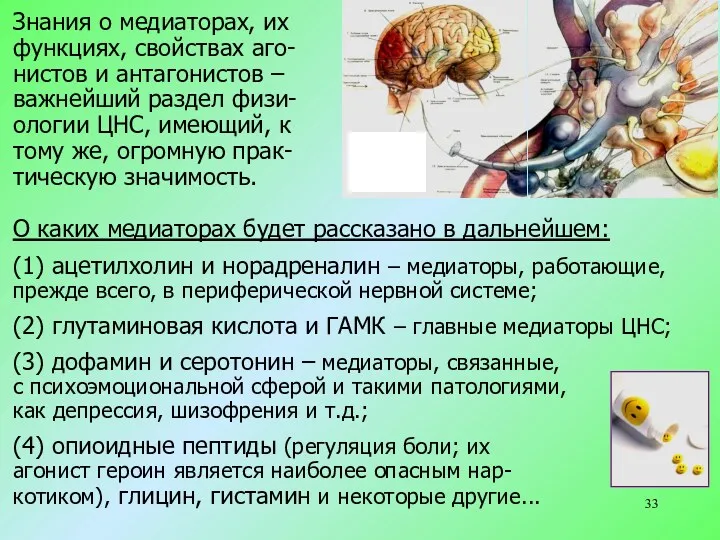
Знания о медиаторах, их
функциях, свойствах аго-
нистов и антагонистов –
важнейший
Знания о медиаторах, их
функциях, свойствах аго-
нистов и антагонистов –
важнейший

И наконец…
Еще раз подчеркну, что именно синапс (а не
нейрон) является элементарной
И наконец…
Еще раз подчеркну, что именно синапс (а не
нейрон) является элементарной
 Влияние физических факторов на микроорганизмы
Влияние физических факторов на микроорганизмы Артерии большого и малого круга кровообращения
Артерии большого и малого круга кровообращения презентация Цветок
презентация Цветок Организменный уровень жизни, его роль в природе
Организменный уровень жизни, его роль в природе Термин популяция
Термин популяция Кишечнополостные. Губки
Кишечнополостные. Губки Видоизменения листьев
Видоизменения листьев МАТЕМАТИКА и БИОЛОГИЯ ... НА КУХНЕ?
МАТЕМАТИКА и БИОЛОГИЯ ... НА КУХНЕ? Класс Паукообразные. Паукообразные, как и насекомые
Класс Паукообразные. Паукообразные, как и насекомые Способы размножения животных
Способы размножения животных Механизмы влияния гуминовых препаратов на физиолого-биохимические процессы, происходящие в растениях
Механизмы влияния гуминовых препаратов на физиолого-биохимические процессы, происходящие в растениях Комнатные растения (часть 2)
Комнатные растения (часть 2) Рослини і тварини Сумщини занесені до Червоної книги України
Рослини і тварини Сумщини занесені до Червоної книги України 20231130_8_klass_ptitsy._obshchie_priznaki._osobennosti_stroeniya._1
20231130_8_klass_ptitsy._obshchie_priznaki._osobennosti_stroeniya._1 Световая микроскопия. Занятие 1
Световая микроскопия. Занятие 1 Пищевые связи в сообществе
Пищевые связи в сообществе Митохондрия
Митохондрия Женская половая система. (Часть 3)
Женская половая система. (Часть 3) Группы крови. Переливание крови.
Группы крови. Переливание крови. Жабы и лягушки. 2 класс
Жабы и лягушки. 2 класс Законы зависимости организмов от факторов среды
Законы зависимости организмов от факторов среды Периферическая нервная система животных
Периферическая нервная система животных Семейство растений яснотковые
Семейство растений яснотковые В гости к весне (окружающий мир, 2 класс)
В гости к весне (окружающий мир, 2 класс) Лесные жители
Лесные жители Мышечная ткань
Мышечная ткань Размножение многоклеточных животных
Размножение многоклеточных животных Модификационная изменчивость
Модификационная изменчивость