- Хризофиты – золотистые водоросли

Содержание
- 2. Внешний вид под световым микроскопом Питание: псевдоподия захваты-вает пищевые частицы, формирует пищеварительную вакуоль и выбрасывает непереваренные
- 3. Другие хризофиты могут быть коккоидные, амебоидные, нитчатые, талломные, сифональные Характерная циста хризофитов - кувшинчик с пробкой
- 4. Синуровые часто рассматриваются как самостоятельный класс внутри отдела хризофитов Особенности: клетка покрыта кремнезёмными чешуйками, сцементированными в
- 5. Ксантофиты – жёлто-зелёные водоросли хризофиты без фукоксантина Помимо отсутствия фукоксантина ксантофиты имеют несколько мелких отличий от
- 6. Ксантофиты класса ксантофицеа – собственно жёлто-зелёные водоросли Амебоидные, монадные, пальмеллоидные, коккоидные или нитчатые формы с клеточной
- 7. Рафидофиты – отдельный отдел? не имеют переходной спирали запасают масло, а не углеводы не имеют клеточной
- 8. Вошеривые и ботридиофиты – странные многоклеточные из пресной лужи Плазмодиальные организмы с плотной наружной оболочкой из
- 9. Отдел Диктиохи растения с тенденцией вторичного перехода к голозойному питанию Радиальносимметричная клетка с центральным ядром. Миксотрофы
- 10. Отдел Диктиохи: Силикофлагелляты – хромофиты с радиальным скелетом из кремнезёмных трубок Силикофлагеллата Dictyocha fileura: А –
- 11. Отдел Диктиохи: Пединнелеи Пединнелалес – хромофиты на пути к воротничковым жгутиконосцам, Цилиофридные солнечники уже без хроматофоров
- 12. Отдел Диатомовые (=Бацилляриофиты) все с кремнезёмным двустворчатым панцирем Поперечное сечение клетки АГ – аппарат Гольджи М
- 13. Два класса диатомовей различия не только в симметрии Класс Центрические диатомеи 1 – Coscinodiscus convexus, 2
- 14. Жизненный цикл центрической диатомеи Melosira varians При каждом митотическом делении размеры некоторых дочерних клеток сокращаются. Периодически
- 15. Отдел Бурые водоросли, Феофиты – ближайшие родственники диатомей многоклеточная организация бурых водорослей варьирует от нитчатой до
- 16. Бурые водоросли – ближайшие родственники диатомей Спермий Laminaria angustata Клетка с хроматофором А – вид с
- 17. Подавляющее число бурых водоросдей – прикреплённые макрофиты Средообразующая роль бурых водорослей на литорали (побережье Калифорнии). За
- 18. Эустигматофиты – максимальные отличия от прочих хромофитов ОТЛИЧИЯ: фукоксантина нет, хлорофилла с нет; хлорофилл с замещён
- 19. Филогенетические связи хромофитов ПС С ПС С ПС С замена хлорофилла с виолоксантином в море –фукоксантин,
- 20. Гетероконты – это больше, чем хромофиты Неравные жгутики той же организации – передний с трубчатыми трёхраздельными
- 21. Другие гетероконты: Бикозоеки вроде хризофиты, но бесцветные Siluania monomastiga Adriamonas peritocrescens Собственно бикозоеки – «обитатели горшка»
- 22. Другие гетероконты: Лабиртнтулиды – сетчатые слизевики с эктоплазматической сетью Амебоидные организмы, имеющие уникальные органеллы – ботросомы
- 23. Другие гетероконты: Лабиртинтулиды класс Лабиринтулеи – обитатели эктоплазматических туннелей Веретеновидные клетки находятся внутри каналов, образованных эктоплазматической
- 24. Класс Траустохитридиевые – сетчатые слизевики с эктоплазматическими ризоидами Жизненный цикл Thraustochytrium proliferum – сапрофита на морских
- 25. Другие гетероконты: тип Псевдогрибы (Pseudofungi) Бесцветные, обычно разветвленные, плазмодии без ботросом и эктоплазма-тической сети. Размножаются с
- 26. Псевдогрибы: класс Оомицеты (=Сапролегниевые) - стенка целлюлозная, споры двужгутиковые Зооспоры двужгутиковые, клеточная стенка целлюлозная, не содержит
- 27. Псевдогрибы: класс Оомицеты (=Сапролегниевые) размножаются сначала как грибы, потом - дипланетизм Жизненный цикл сапролегнии А –
- 29. Скачать презентацию
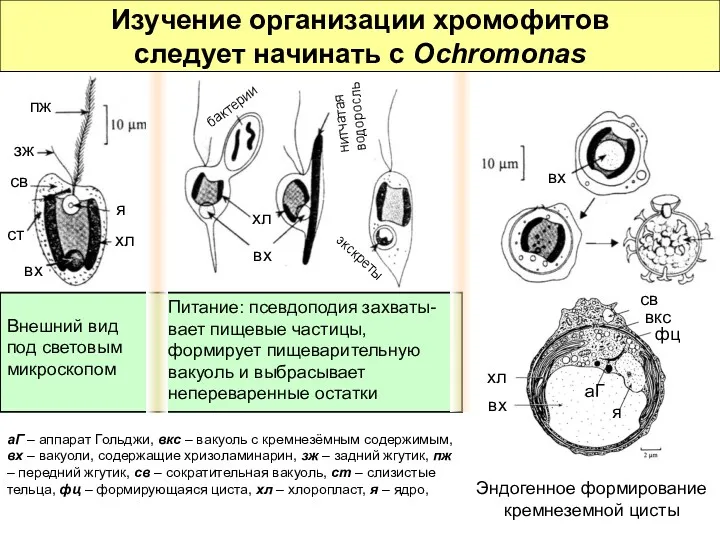
Внешний вид под световым микроскопом
Питание: псевдоподия захваты-вает пищевые частицы, формирует
Внешний вид под световым микроскопом
Питание: псевдоподия захваты-вает пищевые частицы, формирует
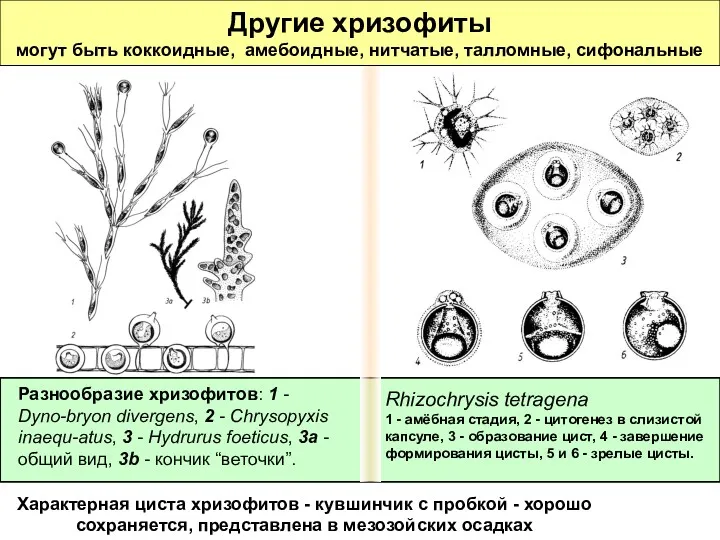
Другие хризофиты
могут быть коккоидные, амебоидные, нитчатые, талломные, сифональные
Характерная циста
Другие хризофиты
могут быть коккоидные, амебоидные, нитчатые, талломные, сифональные
Характерная циста
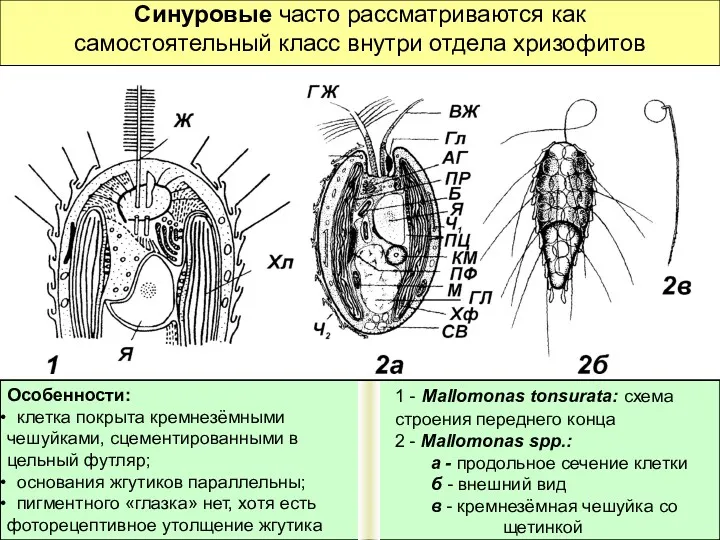
Синуровые часто рассматриваются как самостоятельный класс внутри отдела хризофитов
Особенности:
Синуровые часто рассматриваются как самостоятельный класс внутри отдела хризофитов
Особенности:
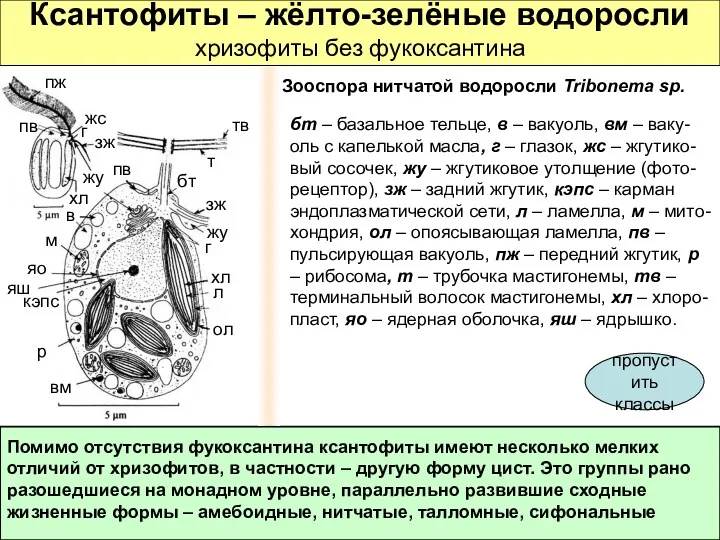
Ксантофиты – жёлто-зелёные водоросли
хризофиты без фукоксантина
Помимо отсутствия фукоксантина ксантофиты имеют
Ксантофиты – жёлто-зелёные водоросли
хризофиты без фукоксантина
Помимо отсутствия фукоксантина ксантофиты имеют
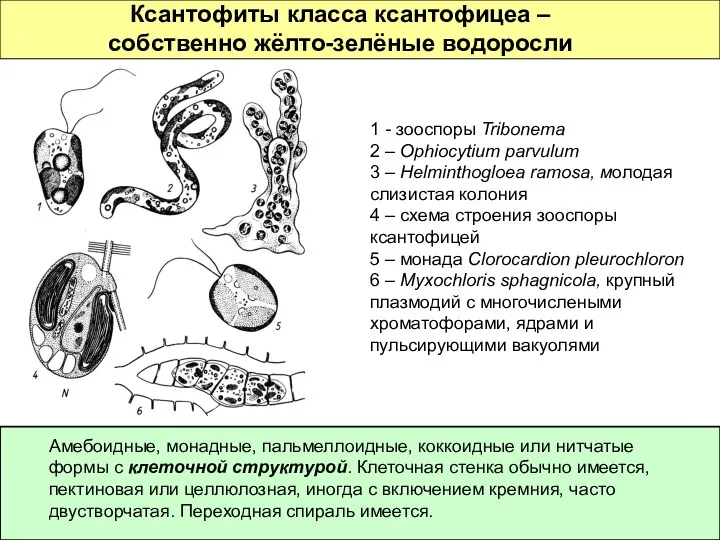
Ксантофиты класса ксантофицеа – собственно жёлто-зелёные водоросли
Амебоидные, монадные, пальмеллоидные, коккоидные
Ксантофиты класса ксантофицеа – собственно жёлто-зелёные водоросли
Амебоидные, монадные, пальмеллоидные, коккоидные
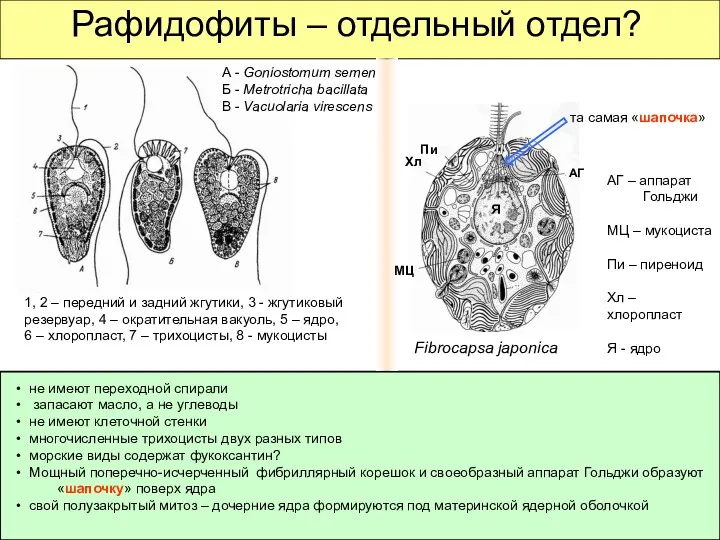
Рафидофиты – отдельный отдел?
не имеют переходной спирали
запасают масло,
Рафидофиты – отдельный отдел?
не имеют переходной спирали
запасают масло,
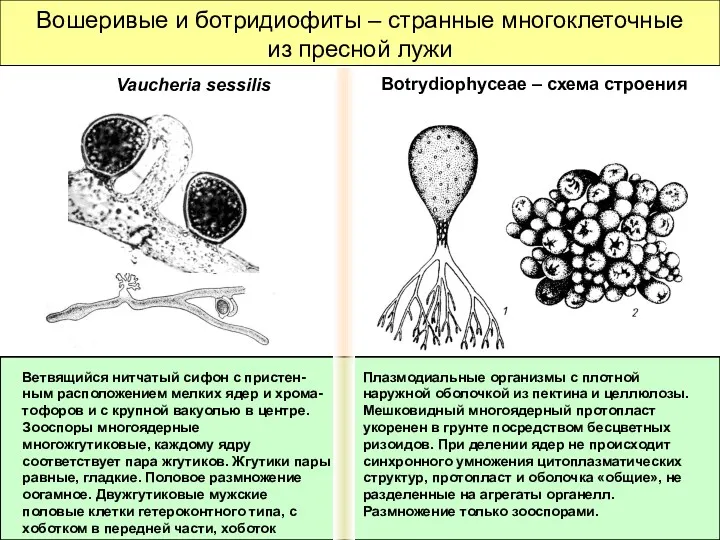
Вошеривые и ботридиофиты – странные многоклеточные из пресной лужи
Плазмодиальные организмы
Вошеривые и ботридиофиты – странные многоклеточные из пресной лужи
Плазмодиальные организмы

Отдел Диктиохи
растения с тенденцией вторичного перехода к голозойному питанию
Отдел Диктиохи
растения с тенденцией вторичного перехода к голозойному питанию
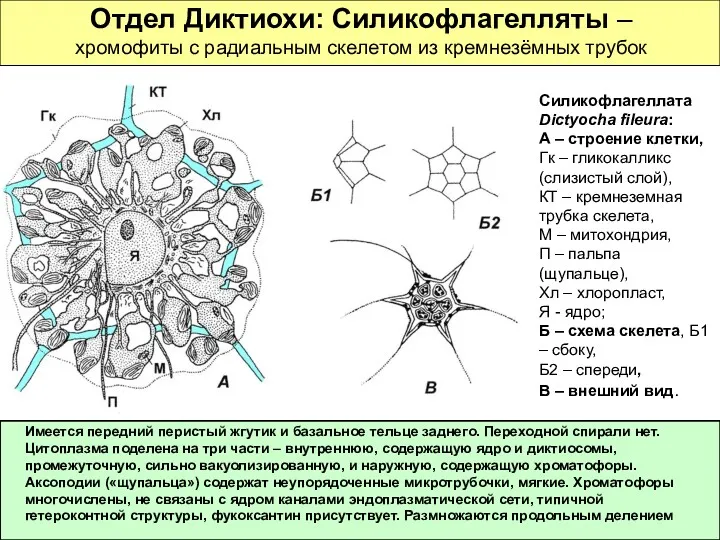
Отдел Диктиохи: Силикофлагелляты – хромофиты с радиальным скелетом из кремнезёмных
Отдел Диктиохи: Силикофлагелляты – хромофиты с радиальным скелетом из кремнезёмных
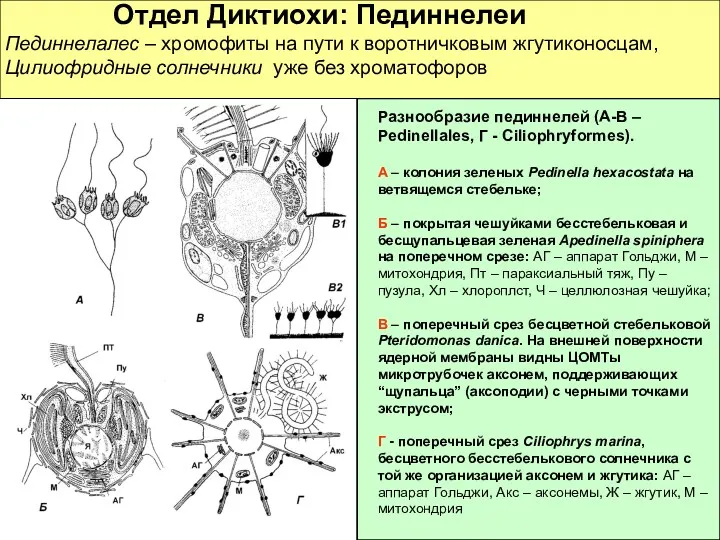
Отдел Диктиохи: Пединнелеи
Пединнелалес – хромофиты на пути к
Отдел Диктиохи: Пединнелеи Пединнелалес – хромофиты на пути к
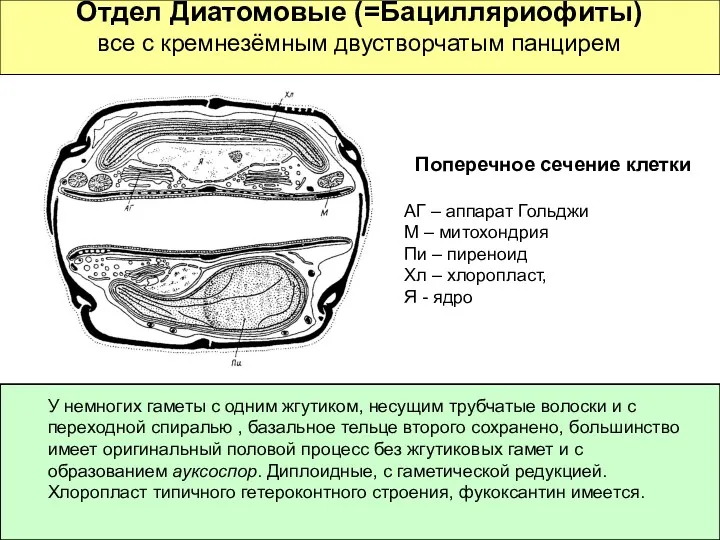
Отдел Диатомовые (=Бацилляриофиты)
все с кремнезёмным двустворчатым панцирем
Поперечное сечение
Отдел Диатомовые (=Бацилляриофиты)
все с кремнезёмным двустворчатым панцирем
Поперечное сечение
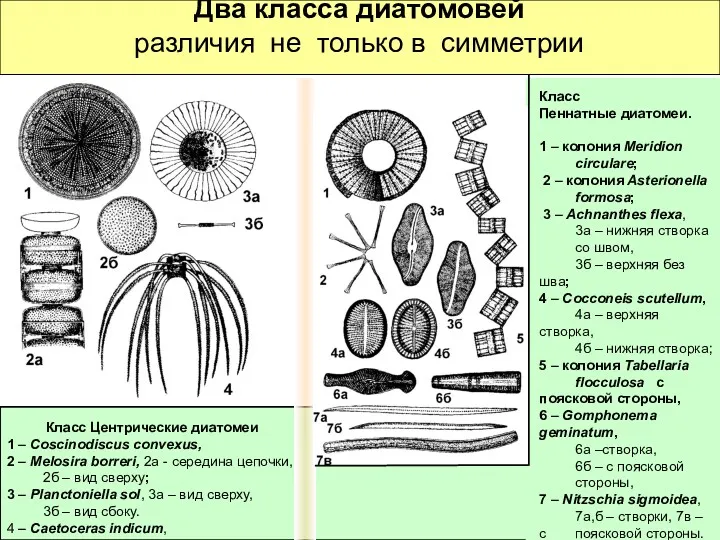
Два класса диатомовей
различия не только в симметрии
Класс Центрические
Два класса диатомовей
различия не только в симметрии
Класс Центрические
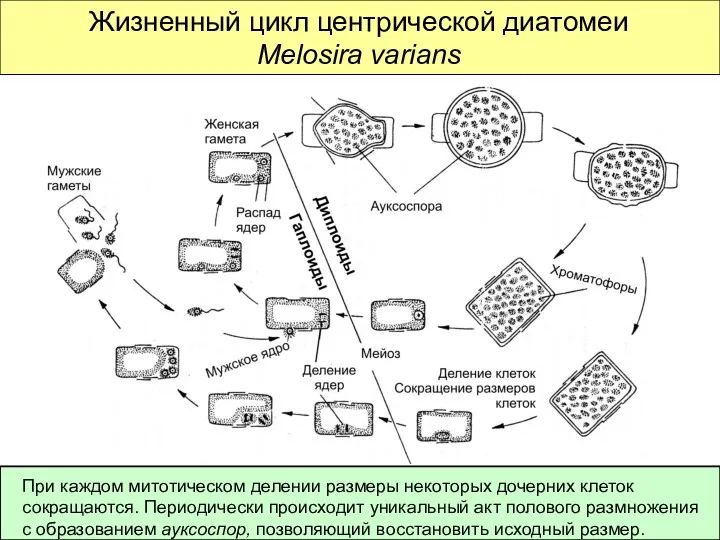
Жизненный цикл центрической диатомеи Melosira varians
При каждом митотическом делении размеры
Жизненный цикл центрической диатомеи Melosira varians
При каждом митотическом делении размеры

Отдел Бурые водоросли, Феофиты –
ближайшие родственники диатомей
многоклеточная организация
Отдел Бурые водоросли, Феофиты –
ближайшие родственники диатомей
многоклеточная организация
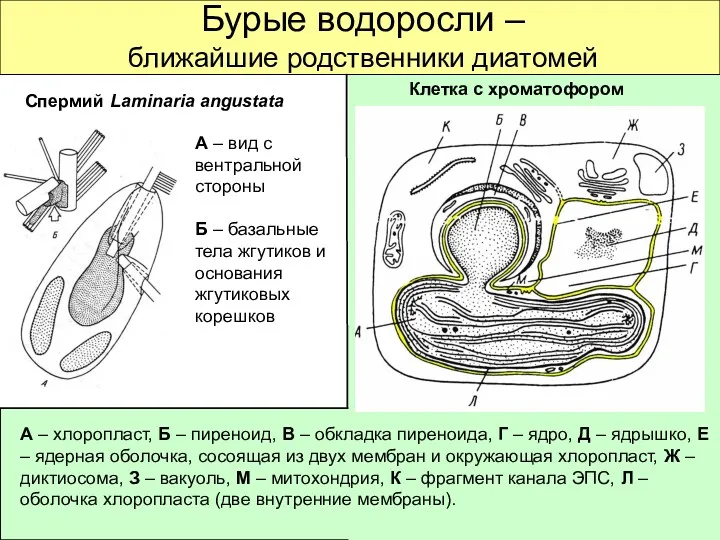
Бурые водоросли –
ближайшие родственники диатомей
Спермий Laminaria angustata
Клетка с хроматофором
А
Бурые водоросли –
ближайшие родственники диатомей
Спермий Laminaria angustata
Клетка с хроматофором
А
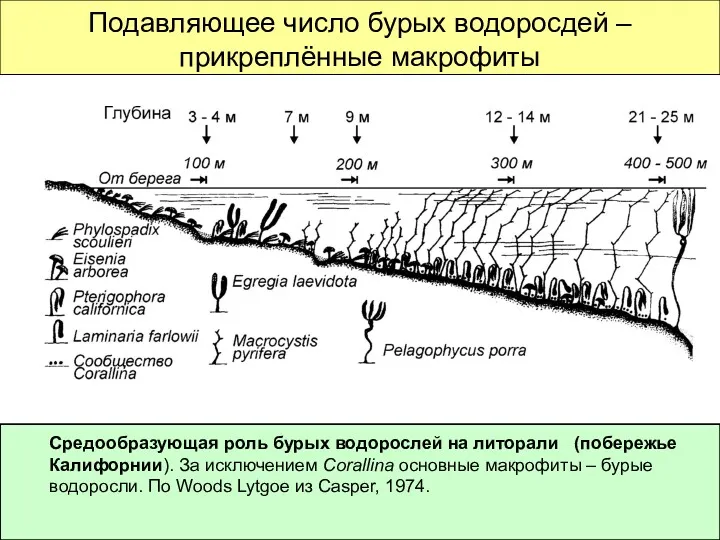
Подавляющее число бурых водоросдей – прикреплённые макрофиты
Средообразующая роль бурых водорослей
Подавляющее число бурых водоросдей – прикреплённые макрофиты
Средообразующая роль бурых водорослей
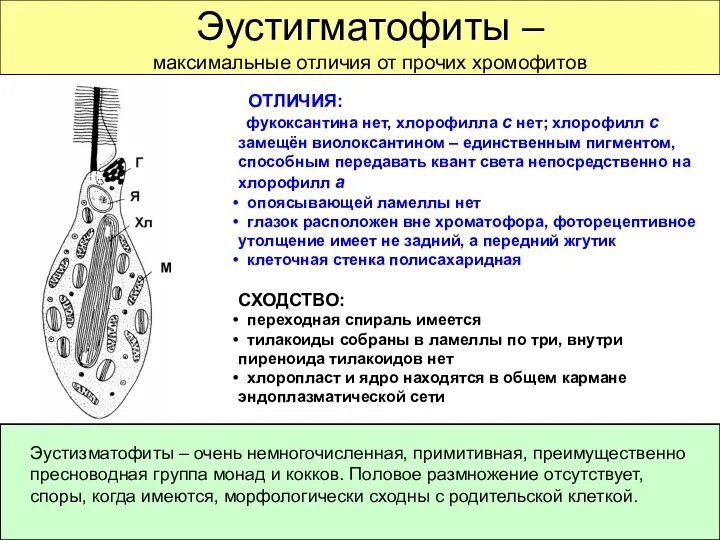
Эустигматофиты –
максимальные отличия от прочих хромофитов
ОТЛИЧИЯ:
фукоксантина
Эустигматофиты –
максимальные отличия от прочих хромофитов
ОТЛИЧИЯ:
фукоксантина
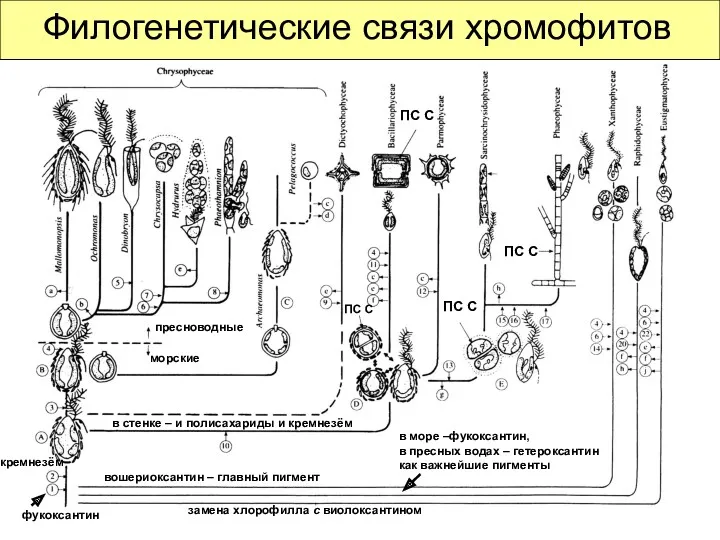
Филогенетические связи хромофитов
ПС С
ПС С
ПС С
замена хлорофилла с виолоксантином
в море –фукоксантин,
в
Филогенетические связи хромофитов
ПС С
ПС С
ПС С
замена хлорофилла с виолоксантином
в море –фукоксантин,
в

Гетероконты – это больше, чем хромофиты
Неравные жгутики той же организации
Гетероконты – это больше, чем хромофиты
Неравные жгутики той же организации
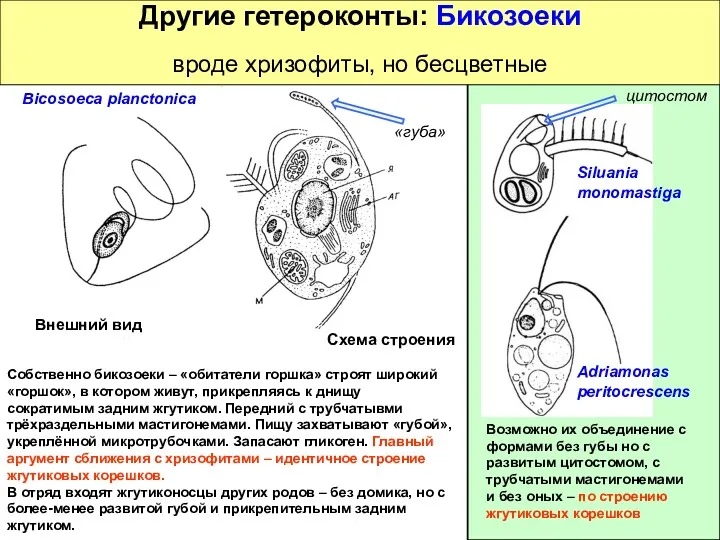
Другие гетероконты: Бикозоеки
вроде хризофиты, но бесцветные
Siluania
monomastiga
Adriamonas
peritocrescens
Собственно бикозоеки –
Другие гетероконты: Бикозоеки
вроде хризофиты, но бесцветные
Siluania
monomastiga
Adriamonas
peritocrescens
Собственно бикозоеки –
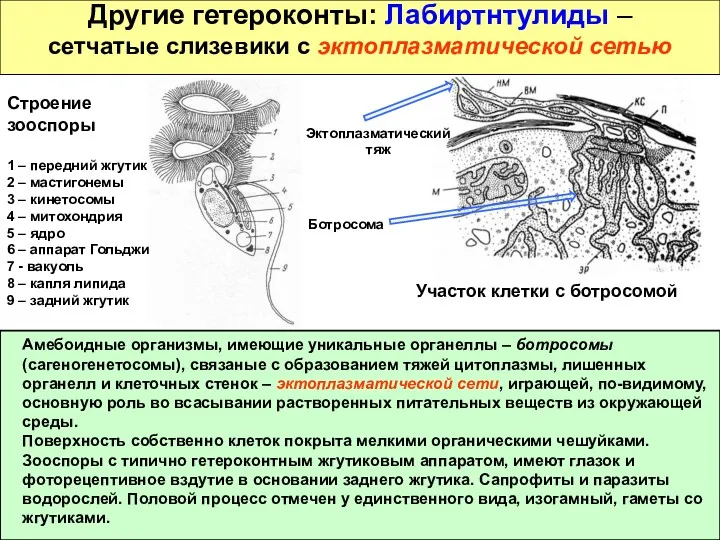
Другие гетероконты: Лабиртнтулиды –
сетчатые слизевики с эктоплазматической сетью
Амебоидные организмы, имеющие
Другие гетероконты: Лабиртнтулиды –
сетчатые слизевики с эктоплазматической сетью
Амебоидные организмы, имеющие
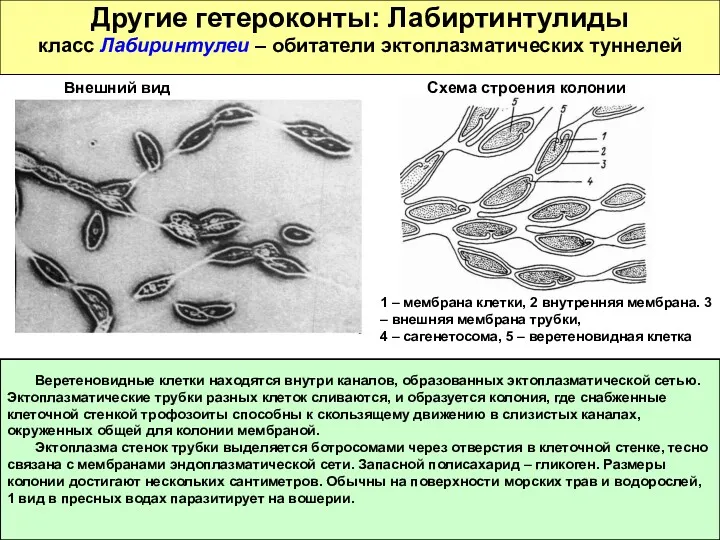
Другие гетероконты: Лабиртинтулиды
класс Лабиринтулеи – обитатели эктоплазматических туннелей
Веретеновидные клетки
Другие гетероконты: Лабиртинтулиды
класс Лабиринтулеи – обитатели эктоплазматических туннелей
Веретеновидные клетки
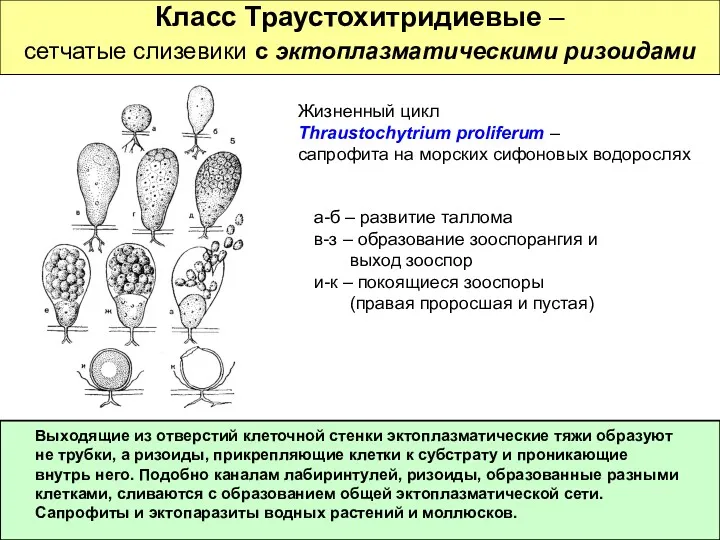
Класс Траустохитридиевые –
сетчатые слизевики с эктоплазматическими ризоидами
Жизненный цикл
Thraustochytrium
Класс Траустохитридиевые –
сетчатые слизевики с эктоплазматическими ризоидами
Жизненный цикл
Thraustochytrium

Другие гетероконты: тип Псевдогрибы (Pseudofungi)
Бесцветные, обычно разветвленные, плазмодии без
Другие гетероконты: тип Псевдогрибы (Pseudofungi)
Бесцветные, обычно разветвленные, плазмодии без
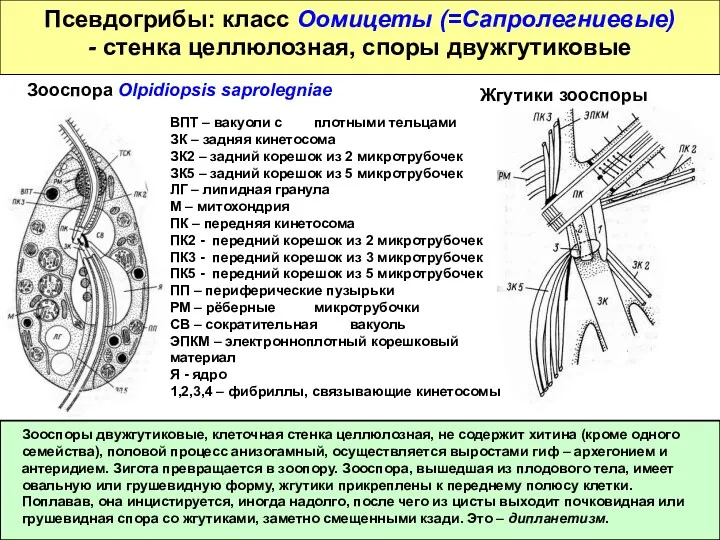
Псевдогрибы: класс Оомицеты (=Сапролегниевые)
- стенка целлюлозная, споры двужгутиковые
Зооспоры двужгутиковые, клеточная
Псевдогрибы: класс Оомицеты (=Сапролегниевые)
- стенка целлюлозная, споры двужгутиковые
Зооспоры двужгутиковые, клеточная
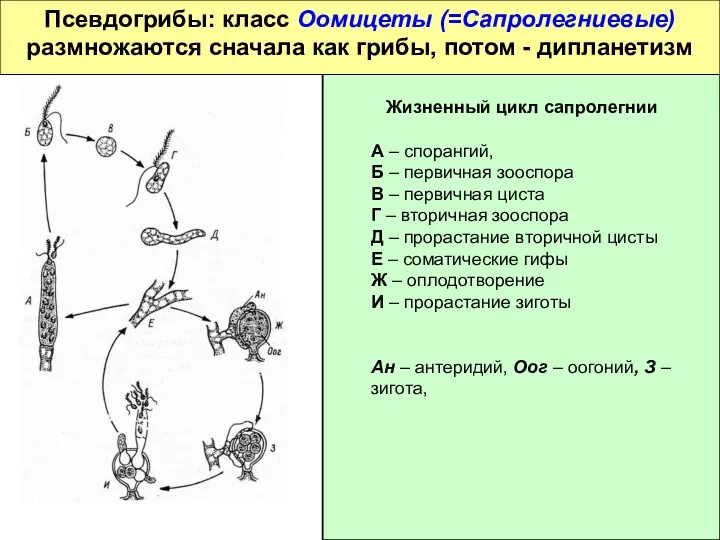
Псевдогрибы: класс Оомицеты (=Сапролегниевые)
размножаются сначала как грибы, потом - дипланетизм
Псевдогрибы: класс Оомицеты (=Сапролегниевые)
размножаются сначала как грибы, потом - дипланетизм
 В мире живой природы. Птицы. (2 класс)
В мире живой природы. Птицы. (2 класс) Декоративные устройства для оформления объектов. Устройство и содержание цветников, вертикальное озеленение, каменистые участки
Декоративные устройства для оформления объектов. Устройство и содержание цветников, вертикальное озеленение, каменистые участки Интересные факты о кожном покрове и его производных
Интересные факты о кожном покрове и его производных Адаптация организмов к условиям обитания
Адаптация организмов к условиям обитания Отряды пресмыкающихся: Черепахи и Крокодилы
Отряды пресмыкающихся: Черепахи и Крокодилы Городские зимующие птицы Сибири
Городские зимующие птицы Сибири Тип Моллюски. Класс Брюхоногие. Класс Двустворчатые. Класс Головоногие
Тип Моллюски. Класс Брюхоногие. Класс Двустворчатые. Класс Головоногие Расы человека
Расы человека Селекция и методы ее исследования
Селекция и методы ее исследования Презентация Энерготраты человека: основной и общий обмен.
Презентация Энерготраты человека: основной и общий обмен. Половое размножение у покрытосеменных растений
Половое размножение у покрытосеменных растений Прокариоты, бактерии, цианобактерии
Прокариоты, бактерии, цианобактерии Психологические особенности личности
Психологические особенности личности Виділення, його значення для організму. Органи виділення тварин
Виділення, його значення для організму. Органи виділення тварин Почему листья на деревьях осенью желтеют, а на комнатных растениях нет?
Почему листья на деревьях осенью желтеют, а на комнатных растениях нет? Введение 01 02 2016
Введение 01 02 2016 Презентация Кольчатые черви
Презентация Кольчатые черви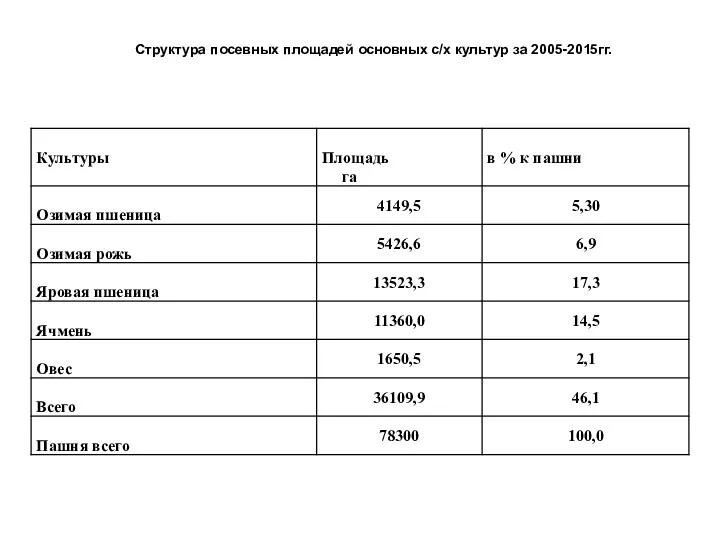 Структура посевных площадей основных с/х культур
Структура посевных площадей основных с/х культур Млекопитающие зайцы
Млекопитающие зайцы Внешнее строение и разнообразие листьев
Внешнее строение и разнообразие листьев Органи і системи органів тварин
Органи і системи органів тварин Развитие до имплантации
Развитие до имплантации Вегетативная нервная система. Лекция №6
Вегетативная нервная система. Лекция №6 Размножение организмов
Размножение организмов Морфология бактерий, грибов, спирохет, актиномицетов, микоплазм, риккетсий и хламидий
Морфология бактерий, грибов, спирохет, актиномицетов, микоплазм, риккетсий и хламидий Многолетние цветковые растения
Многолетние цветковые растения Углеводы, липиды
Углеводы, липиды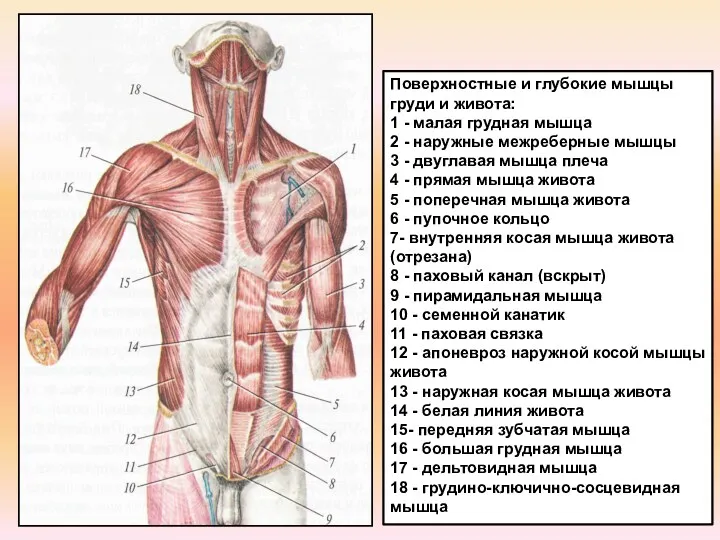 Поверхностные и глубокие мышцы груди и живота
Поверхностные и глубокие мышцы груди и живота