- Хромосомная теория наследственности. Сцепленное наследование

Содержание
- 2. Сцепленное наследование. Опыты Т. Моргана. Наследование при полном и неполном сцеплении. Образование гамет. Кроссинговер. Виды кроссинговера.
- 3. Вскоре после переоткрытия законов Менделя немецкий цитолог Теодор Бовери (1902) представил доказательства в пользу участия хромосом
- 4. Уильям Сэттон предположил, что в одной хромосоме может находиться несколько генов. В этом случае должно наблюдаться
- 5. Однако, в 1906 году английские генетики У. Бэтсон и Р. Пеннет, проводят скрещивание растений и анализируют
- 6. Но ученые не смогли вскрыть причины этого явления!!! Ученые изучали наследование окраски цветка (пурпурная или красная)
- 7. Стало ясно, что не для всех признаков характерно независимое распределение в потомстве и свободное комбинирование. Каждый
- 8. Морган выбрал для объекта основания - мушку дрозофилу. 1. Дает многочисленное потомство. 2. Самец и самка
- 9. Схема анализирующего скрещивания гомозиготных самок с гетерозиготными самцами По результатам этого скрещивания Морган сделал вывод, что
- 10. Схема анализирующего скрещивания гетерозиготных самок с гомозиготными самцами Однако в потомстве явно преобладали особи с признаками
- 11. Результаты опытов Моргана показывают, что существует сцепление генов и в определенном % случаев оно нарушается вследствие
- 12. В зависимости от особенностей образования гамет, различают: некроссоверные гаметы — гаметы с хромосомами, образованными без кроссинговера.
- 13. Величина перекреста и линейное расположение генов в хромосоме Величина кроссинговера измеряется отношением числа кроссоверных особей к
- 14. Т. Морган предположил, что частота кроссинговера показывает расстояние между генами: чем чаще происходит кроссинговер, тем далее
- 15. Гипотеза линейного расположения генов в хромосоме. Одним из генетических опытов Моргана, доказывающих линейное расположение генов, был
- 16. В потомстве было получено 1160 мух некроссоверных (нормальных и одновременно несущих все три рецессивных признака), 15
- 17. Морган сказал, что процент перекреста является функцией расстояния между генами и их последовательного, т. е. линейного,
- 18. Морган предположил, что перекрест между гомологичными хромосомами может происходить одновременно в нескольких точках.
- 21. Пусть , например в гомологичной паре хромосом содержатся три пары аллелей в гетерозиготном состоянии: АВС авс
- 22. По % кроссоверных классов устанавливают частоту произошедших одинарных обменов между генами А и В или В
- 23. Одинарные и двойные перекресты между гомологичными хромосомами доказываются генетическим анализом. Вероятность двойного перекреста всегда меньше вероятности
- 24. Для обозначения частоты кроссинговера была предложена мерная единица – морганида (в честь Т.Моргана), соответствующая 1% кроссинговера.
- 25. Морган заявил: чем дальше стоят друг от друга в хромосоме гены А и С , тем
- 26. Интерференция – это подавление кроссинговера на участках, непосредственно прилегающих к точке происшедшего обмена. Открыл 1816 г.
- 27. Ученик Моргана Г. Меллер предложил определять интенсивность интерференции количественно, путем деления фактически наблюдаемой частоты двойного кроссинговера
- 28. Определение положения гена на хромосоме Для определения локализации гена на хромосоме необходимо определить группу сцепления.
- 29. - Гены одной хромосомы образуют группу сцепления и происходит сцепленное наследование признаков. - Сила сцепления находиться
- 30. Неравный кроссинговер связан с дупликацией какого-либо участка в одном из гомологов и с утратой его в
- 34. Явление кроссинговера помогло ученым установить расположение каждого гена в хромосоме, создать генетические карты хромосом. Чем дальше
- 35. Следовательно, подсчитав частоту кроссинговера между какими-либо двумя генами одной хромосомы, отвечающими за различные признаки, можно точно
- 37. Скачать презентацию

Сцепленное наследование.
Опыты Т. Моргана.
Наследование при полном и неполном сцеплении.
Образование гамет.
Кроссинговер.
Сцепленное наследование.
Опыты Т. Моргана.
Наследование при полном и неполном сцеплении.
Образование гамет.
Кроссинговер.

Вскоре после переоткрытия законов Менделя немецкий цитолог Теодор Бовери (1902) представил
Вскоре после переоткрытия законов Менделя немецкий цитолог Теодор Бовери (1902) представил

Уильям Сэттон предположил, что в одной хромосоме может находиться несколько генов.
Уильям Сэттон предположил, что в одной хромосоме может находиться несколько генов.

Однако, в 1906 году английские генетики У. Бэтсон и Р.
Однако, в 1906 году английские генетики У. Бэтсон и Р.
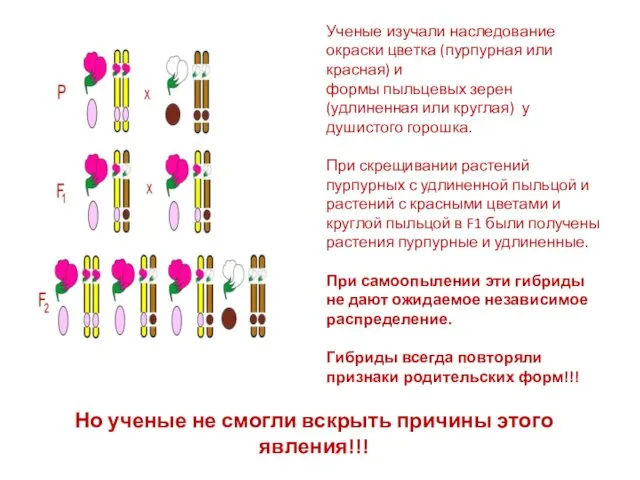
Но ученые не смогли вскрыть причины этого явления!!!
Ученые изучали наследование окраски
Но ученые не смогли вскрыть причины этого явления!!!
Ученые изучали наследование окраски

Стало ясно, что не для всех признаков характерно независимое распределение в
Стало ясно, что не для всех признаков характерно независимое распределение в

Морган выбрал для объекта основания - мушку дрозофилу.
1. Дает многочисленное
1. Дает многочисленное

Схема анализирующего скрещивания гомозиготных самок с гетерозиготными самцами
По результатам этого
Схема анализирующего скрещивания гомозиготных самок с гетерозиготными самцами
По результатам этого
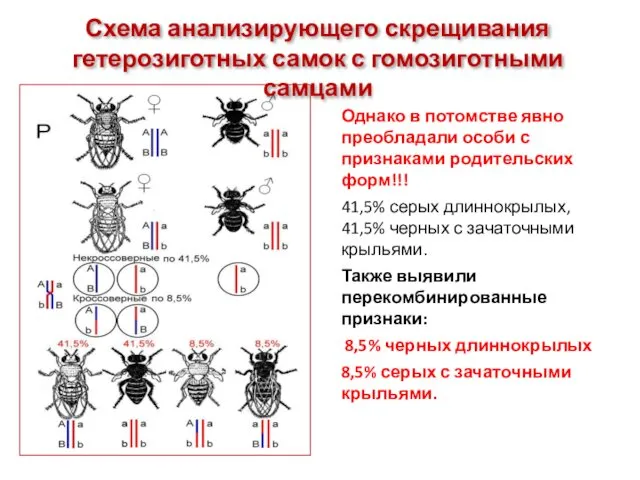
Схема анализирующего скрещивания гетерозиготных самок с гомозиготными самцами
Однако в потомстве явно
Схема анализирующего скрещивания гетерозиготных самок с гомозиготными самцами
Однако в потомстве явно
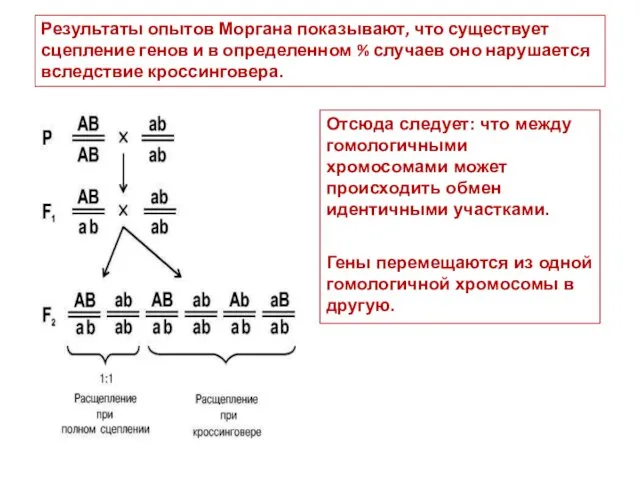
Результаты опытов Моргана показывают, что существует сцепление генов и в определенном
Результаты опытов Моргана показывают, что существует сцепление генов и в определенном
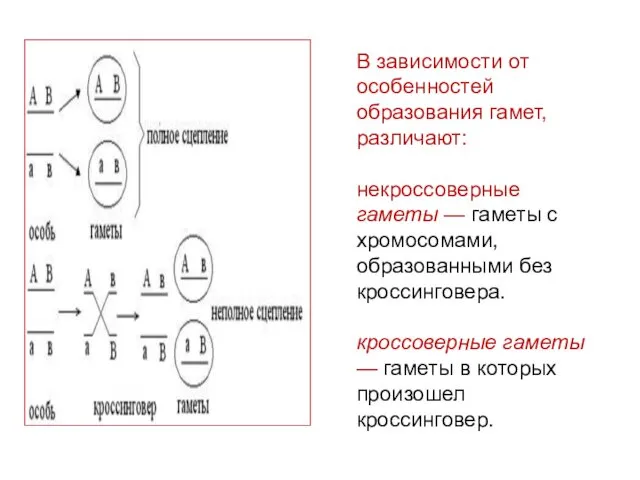
В зависимости от особенностей образования гамет, различают:
некроссоверные гаметы — гаметы с
В зависимости от особенностей образования гамет, различают:
некроссоверные гаметы — гаметы с
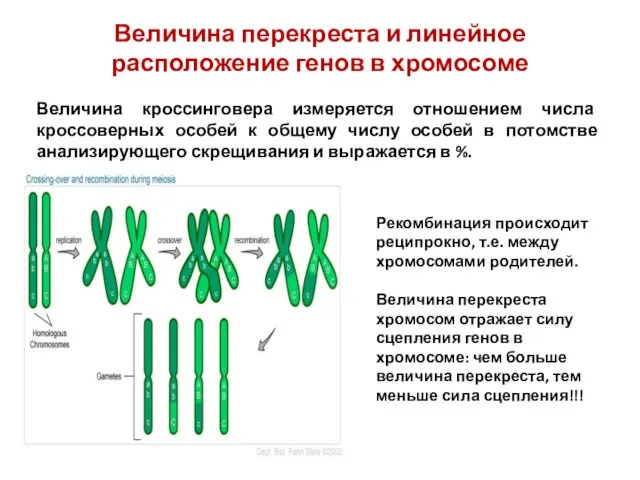
Величина перекреста и линейное расположение генов в хромосоме
Величина кроссинговера измеряется отношением
Величина перекреста и линейное расположение генов в хромосоме
Величина кроссинговера измеряется отношением
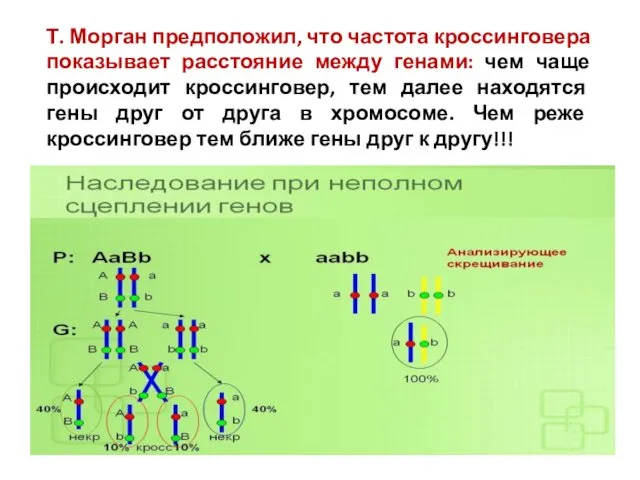
Т. Морган предположил, что частота кроссинговера показывает расстояние между генами: чем
Т. Морган предположил, что частота кроссинговера показывает расстояние между генами: чем

Гипотеза линейного расположения генов в хромосоме.
Одним из генетических опытов Моргана,
Гипотеза линейного расположения генов в хромосоме.
Одним из генетических опытов Моргана,
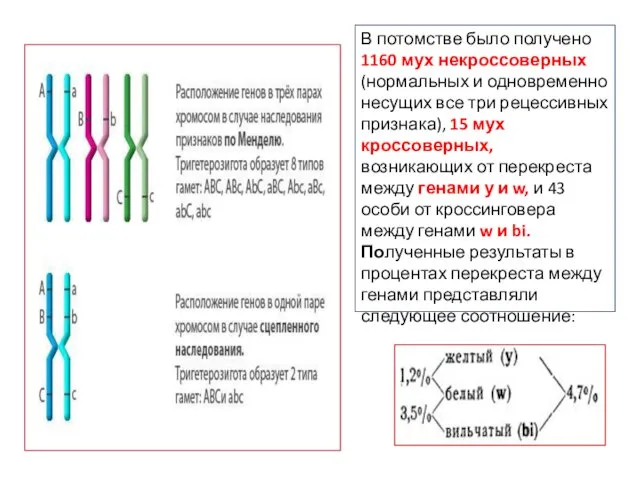
В потомстве было получено 1160 мух некроссоверных (нормальных и одновременно несущих
В потомстве было получено 1160 мух некроссоверных (нормальных и одновременно несущих

Морган сказал, что процент перекреста является функцией расстояния между генами и
Морган сказал, что процент перекреста является функцией расстояния между генами и
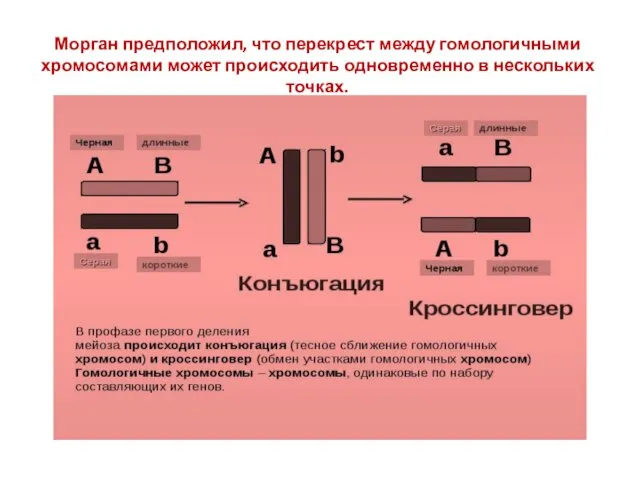
Морган предположил, что перекрест между гомологичными хромосомами может происходить одновременно в
Морган предположил, что перекрест между гомологичными хромосомами может происходить одновременно в



Пусть , например в гомологичной паре хромосом содержатся три пары аллелей
Пусть , например в гомологичной паре хромосом содержатся три пары аллелей
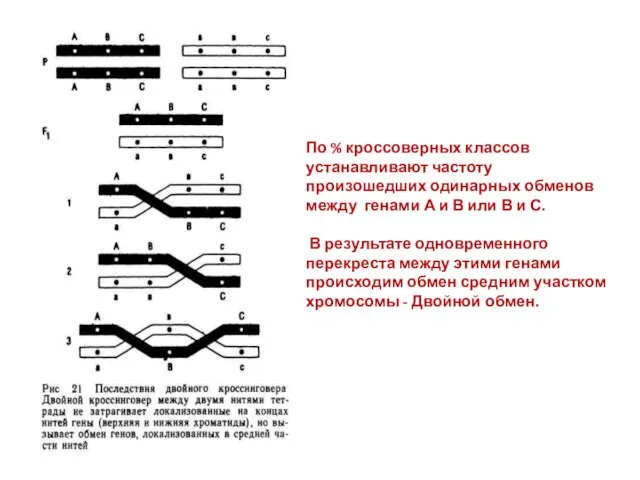
По % кроссоверных классов устанавливают частоту произошедших одинарных обменов между генами
По % кроссоверных классов устанавливают частоту произошедших одинарных обменов между генами

Одинарные и двойные перекресты между гомологичными хромосомами доказываются генетическим анализом.
Вероятность
Одинарные и двойные перекресты между гомологичными хромосомами доказываются генетическим анализом.
Вероятность

Для обозначения частоты кроссинговера была предложена мерная единица – морганида (в
Для обозначения частоты кроссинговера была предложена мерная единица – морганида (в

Морган заявил: чем дальше стоят друг от друга в хромосоме гены
Морган заявил: чем дальше стоят друг от друга в хромосоме гены

Интерференция – это подавление кроссинговера на участках, непосредственно прилегающих к точке
Интерференция – это подавление кроссинговера на участках, непосредственно прилегающих к точке

Ученик Моргана Г. Меллер предложил определять интенсивность интерференции количественно, путем деления
Ученик Моргана Г. Меллер предложил определять интенсивность интерференции количественно, путем деления

Определение положения гена на хромосоме
Для определения локализации гена на хромосоме
Определение положения гена на хромосоме
Для определения локализации гена на хромосоме

- Гены одной хромосомы образуют группу сцепления и происходит сцепленное наследование
- Гены одной хромосомы образуют группу сцепления и происходит сцепленное наследование
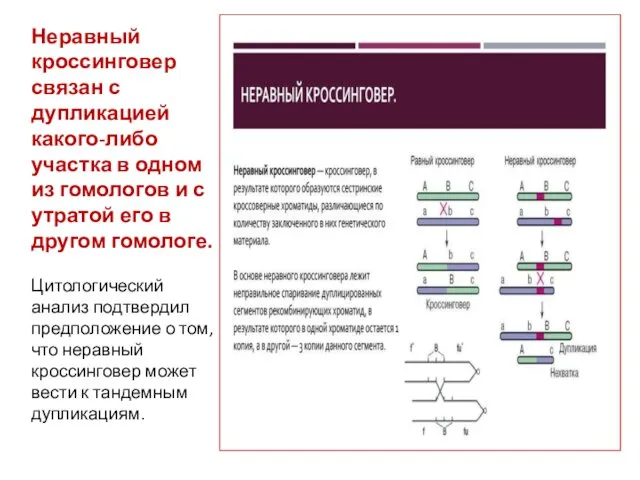
Неравный кроссинговер связан с дупликацией какого-либо участка в одном из гомологов
Неравный кроссинговер связан с дупликацией какого-либо участка в одном из гомологов

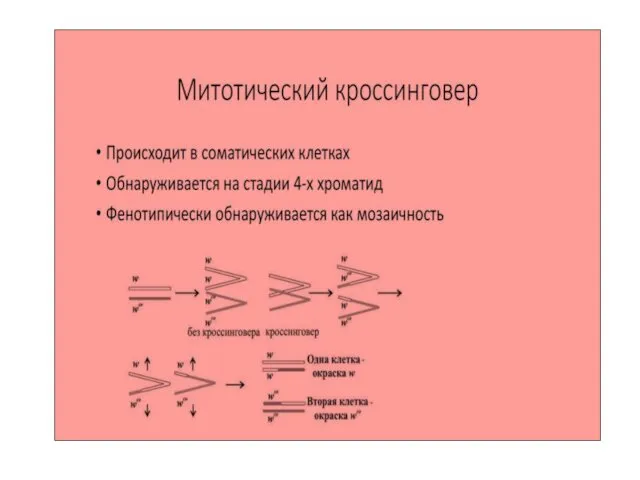


Явление кроссинговера помогло ученым установить расположение каждого гена в хромосоме, создать
Явление кроссинговера помогло ученым установить расположение каждого гена в хромосоме, создать

Следовательно, подсчитав частоту кроссинговера между какими-либо двумя генами одной хромосомы, отвечающими
Следовательно, подсчитав частоту кроссинговера между какими-либо двумя генами одной хромосомы, отвечающими
 Индивидуальное развитие организмов
Индивидуальное развитие организмов Хрящевые рыбы
Хрящевые рыбы Пищеварение в 12-перстной кишке
Пищеварение в 12-перстной кишке Внеклассное мероприятие Цветы
Внеклассное мероприятие Цветы Внутреннее строение млекопитающих
Внутреннее строение млекопитающих уроки и презентации по биологии 8 класс
уроки и презентации по биологии 8 класс Среда обитания первых форм жизни. Происхождение нуклеотидов и РНК
Среда обитания первых форм жизни. Происхождение нуклеотидов и РНК Бактерии. История открытия. Строение бактерий
Бактерии. История открытия. Строение бактерий Система органов размножения самок и самцов, кровеносная и лимфатическая системы
Система органов размножения самок и самцов, кровеносная и лимфатическая системы Предмет и задачи селекции
Предмет и задачи селекции Животноводство
Животноводство Экологические сукцеcсии
Экологические сукцеcсии Кожа. Строение и функции кожи
Кожа. Строение и функции кожи ПРЕЗЕНТАЦИЯ ДЛЯ ИНТЕРАКТИВНОЙ ДОСКИ. Введение в общую биологию. (В четырёх частях).
ПРЕЗЕНТАЦИЯ ДЛЯ ИНТЕРАКТИВНОЙ ДОСКИ. Введение в общую биологию. (В четырёх частях). чому скисає молоко
чому скисає молоко Мутационная изменчивость
Мутационная изменчивость Воспроизведение на молекулярном и клеточном уровнях
Воспроизведение на молекулярном и клеточном уровнях Cerphidae. Ден
Cerphidae. Ден Движение крови в организме. Органы кровообращения
Движение крови в организме. Органы кровообращения Цепи питания и пищевые сети
Цепи питания и пищевые сети Heart valves: functionality and treats
Heart valves: functionality and treats Первичные и вторичные метаболиты
Первичные и вторичные метаболиты Ботаника для ЕГЭ
Ботаника для ЕГЭ Организация научно-исследовательской деятельности учащихся с использованием материалов по краеведению
Организация научно-исследовательской деятельности учащихся с использованием материалов по краеведению Генотип як цілісна система
Генотип як цілісна система Витамины и их значение
Витамины и их значение Bacteria Cell Structure
Bacteria Cell Structure Төменгі сатыдағы өсімдіктер, балдырлар
Төменгі сатыдағы өсімдіктер, балдырлар