- Среда обитания первых форм жизни. Происхождение нуклеотидов и РНК

Содержание
- 2. «ПЕРВИЧНЫЙ БУЛЬОН» ИЛИ «ПЕРВИЧНАЯ ПИЦЦА» ? Для синтеза полимеров хороши условия периодического высыхания (приливная зона моря
- 3. ОТБОР РНК-ПОДОБНЫХ ПОЛИМЕРОВ НА УСТОЙЧИВОСТЬ К УЛЬТРАФИОЛЕТУ А, Г, Т, У исключительно устойчивы к ультрафиолету (в
- 4. МИНЕРАЛЬНЫЙ СОСТАВ ЖИВЫХ КЛЕТОК Кровь похожа на морскую воду. А цитоплазма – на место, где зародились
- 5. МИНЕРАЛЬНЫЙ СОСТАВ ЖИВЫХ КЛЕТОК И МЕСТО ИХ ПРОИСХОЖДЕНИЯ Высокое содержание К и низкое – Na встречается
- 6. ВОССТАНОВЛЕНИЕ СО2 В ГЕОТЕРМАЛЬНЫХ СИСТЕМАХ: ЖЕЛЕЗО-СЕРНЫЙ МИР FeS + H2S → FeS2 + 2 [H] (150-200
- 7. СШИВАНИЕ АМИНОКИСЛОТ В ПЕПТИДЫ ПРИ ПОМОЩИ COS Уходящие группы – H2S и CO2
- 8. ВОССТАНОВЛЕНИЕ СО2 В ГЕОТЕРМАЛЬНЫХ СИСТЕМАХ: ЩЕЛОЧНЫЕ ИСТОЧНИКИ Температуры 40-100 'C, pH 10-11 Осаждение Mg(OH)2 и CaCO3
- 9. ВОССТАНОВЛЕНИЕ СО2 : ЦИНКОВЫЙ МИР И АБИОГЕННЫЙ ФОТОСИНТЕЗ Кристаллы ZnS постепенно разрушаются, цинк выходит в раствор
- 10. ЦИНКОВЫЙ МИР ИЛИ ЖЕЛЕЗНЫЙ МИР? В рибозимах и древнейших белках встречается Zn, Mn и Co, но
- 11. ДОСТУПНОСТЬ ФОСФОРА Обычно находится в виде Ca3(PO4)2 - нерастворим В вулканических газах летят оксиды фосфора P4O10,
- 12. РАЗДЕЛЕНИЕ ВЕЩЕСТВ ПРИ КИПЕНИИ ВОДЫ В ГЕОТЕРМАЛЬНЫХ ИСТОЧНИКАХ K, Mg, Zn, Mn, Mo, P, B переходят
- 13. АВТОКАТАЛИТИЧЕСКИЕ РЕАКЦИИ Автокаталитические реакции в биохимии: Цикл Кальвина Биосинтез большинства витаминов Жизнь в целом с точки
- 14. СИНТЕЗ НУКЛЕОТИДОВ ПРОИСХОЖДЕНИЕ ХИРАЛЬНОЙ ЧИСТОТЫ Лекция 5
- 15. ЧТО НАМ СТОИТ РНК ПОСТРОИТЬ ? Азотистые основания (аденин, гуанин, цитозин, урацил) Рибоза Фосфат Соединить их
- 16. РЕАКЦИЯ БУТЛЕРОВА: СИНТЕЗ САХАРОВ Водный раствор формальдегида (СН2О), Са(ОН)2, небольшое нагревание Образуются все возможные сахара одновременно
- 17. СИНТЕЗ АЗОТИСТЫХ ОСНОВАНИЙ Аденин и гуанин - из HCN при замерзании водного раствора, УФ-облучении, нагревании. При
- 18. ИСТОЧНИКИ ФОРМАЛЬДЕГИДА И ЦИАНИДА
- 19. СИНТЕЗ НУКЛЕОТИДОВ Аденин при УФ-облучении водного раствора присоединяет рибозу и до 3 фосфатных групп Гуанин присоединяет
- 20. ОБХОДНОЙ ПУТЬ СИНТЕЗА ЦИТИДИНОВЫХ И ТИМИДИНОВЫХ НУКЛЕОТИДОВ Фосфат катализирует нужные реакции Побочные продукты первых реакций катализируют
- 21. СИНТЕЗ АКТИВИРОВАННЫХ АДЕНИНОВЫХ И ГУАНИНОВЫХ НУКЛЕОТИДОВ Для получения пуринов в той же системе достаточно добавить синильную
- 22. СИНТЕЗ ГЛИКОАЛЬДЕГИДА И ГЛИЦЕРАЛЬДЕГИДА На древней Земле не было Pd / BaSO4 Подходящий природный восстановитель —
- 23. ЦИАНОСУЛЬФИДНЫЙ ПРОТОМЕТАБОЛИЗМ Фосфат вызывает побочные реакции: Глицеральдегид → диоксиацетон → Val, Leu Цианоацетилен → малеонитрил →
- 24. МЕХАНИЗМЫ ПОЯВЛЕНИЯ ХИРАЛЬНОЙ ЧИСТОТЫ В метеоритной органике есть преобладание левых (L) аминокислот над правыми (D) Избирательное
- 25. МЕХАНИЗМЫ УСИЛЕНИЯ ХИРАЛЬНОЙ ЧИСТОТЫ Аминокислоты обычно выпадают в осадок в соотношении L:D 1:1 – раствор обогащается
- 26. СИНТЕЗ ХИРАЛЬНО ЧИСТЫХ НУКЛЕОТИДОВ L-аминокислоты связывают L-глицеральдегид в побочную реакцию Из D-глицеральдегида образуются нуклеотиды с D-рибозой
- 27. КОПИРОВАНИЕ РНК НАЧАЛО РНК-МИРА Лекция 6
- 28. БЕЗМАТРИЧНАЯ ПОЛИМЕРИЗАЦИЯ НУКЛЕОТИДОВ В РНК На поверхности минералов (глины, сульфиды металлов) из имидазол-активированных нуклеотидов — до
- 29. РИБОЗИМЫ — РНК-ПОЛИМЕРАЗЫ Активные рибозимы-полимеразы состоят из 150-200 нуклеотидов Лучшая полимераза tC19Z во льду удлиняет праймер
- 30. РИБОЗИМЫ — РНК-ЛИГАЗА Гораздо короче полимераз, от 29 нуклеотидов → вероятнее случайное возникновение из абиогенных шпилек
- 31. АБИОГЕННАЯ РЕПЛИКАЦИЯ РНК При высокой концентрации Mg++ короткие РНК реплицируются сами Копируется до 15 нуклеотидов 10%
- 32. НАКОПЛЕНИЕ РНК В ТЕПЛОВОЙ ЛОВУШКЕ Конвекция и термофорез в капиллярах концентрируют РНК и нуклеотиды на много
- 33. МИР ПАЛИНДРОМОВ Дарвиновская эволюция начинается до первого рибозима — отбор идет на самопрайминг Палиндромные РНК богаты
- 35. Скачать презентацию

«ПЕРВИЧНЫЙ БУЛЬОН» ИЛИ «ПЕРВИЧНАЯ ПИЦЦА» ?
Для синтеза полимеров хороши условия периодического
«ПЕРВИЧНЫЙ БУЛЬОН» ИЛИ «ПЕРВИЧНАЯ ПИЦЦА» ?
Для синтеза полимеров хороши условия периодического

ОТБОР РНК-ПОДОБНЫХ ПОЛИМЕРОВ НА УСТОЙЧИВОСТЬ К УЛЬТРАФИОЛЕТУ
А, Г, Т, У исключительно
ОТБОР РНК-ПОДОБНЫХ ПОЛИМЕРОВ НА УСТОЙЧИВОСТЬ К УЛЬТРАФИОЛЕТУ
А, Г, Т, У исключительно
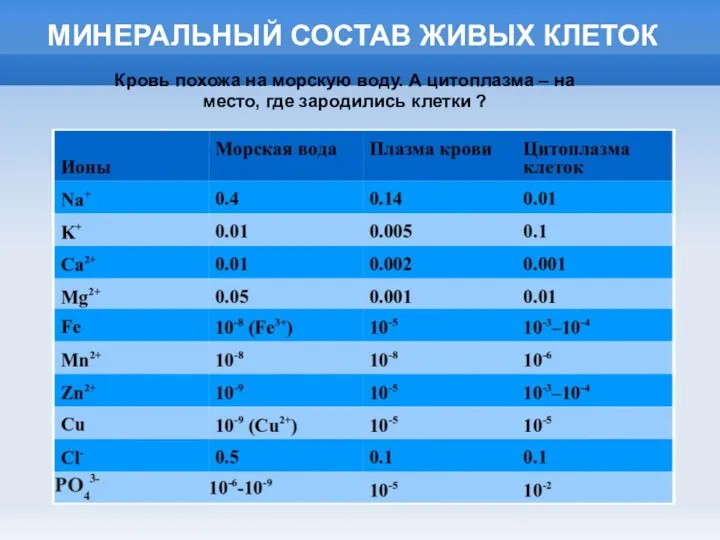
МИНЕРАЛЬНЫЙ СОСТАВ ЖИВЫХ КЛЕТОК
Кровь похожа на морскую воду. А цитоплазма –
МИНЕРАЛЬНЫЙ СОСТАВ ЖИВЫХ КЛЕТОК
Кровь похожа на морскую воду. А цитоплазма –

МИНЕРАЛЬНЫЙ СОСТАВ ЖИВЫХ КЛЕТОК И МЕСТО ИХ ПРОИСХОЖДЕНИЯ
Высокое содержание К и
МИНЕРАЛЬНЫЙ СОСТАВ ЖИВЫХ КЛЕТОК И МЕСТО ИХ ПРОИСХОЖДЕНИЯ
Высокое содержание К и
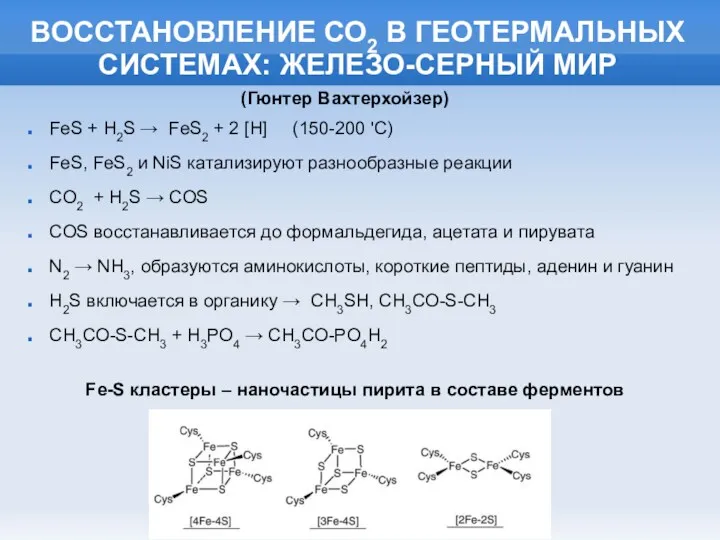
ВОССТАНОВЛЕНИЕ СО2 В ГЕОТЕРМАЛЬНЫХ СИСТЕМАХ: ЖЕЛЕЗО-СЕРНЫЙ МИР
FeS + H2S → FeS2
ВОССТАНОВЛЕНИЕ СО2 В ГЕОТЕРМАЛЬНЫХ СИСТЕМАХ: ЖЕЛЕЗО-СЕРНЫЙ МИР
FeS + H2S → FeS2
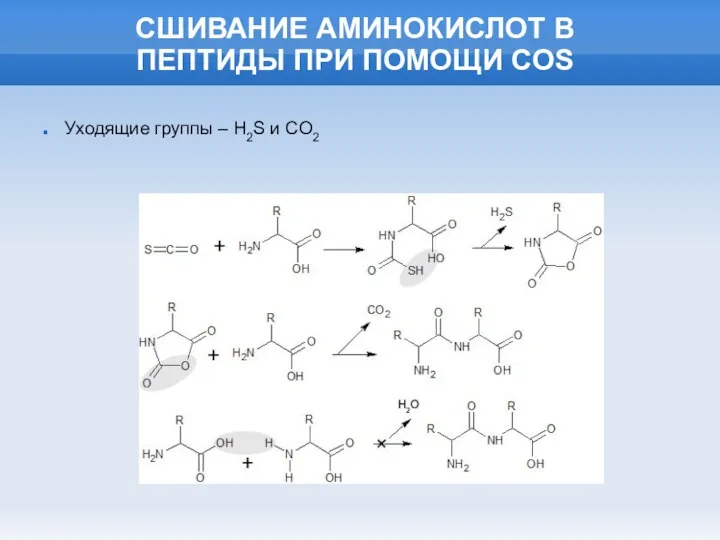
СШИВАНИЕ АМИНОКИСЛОТ В
ПЕПТИДЫ ПРИ ПОМОЩИ COS
Уходящие группы – H2S и
СШИВАНИЕ АМИНОКИСЛОТ В
ПЕПТИДЫ ПРИ ПОМОЩИ COS
Уходящие группы – H2S и
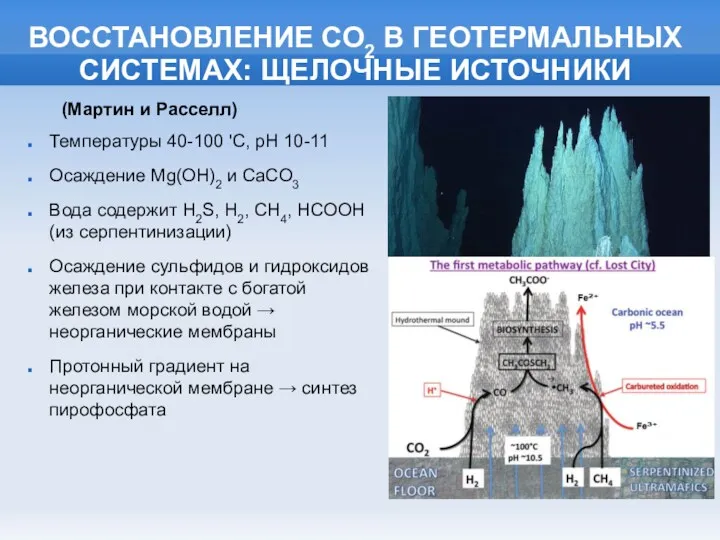
ВОССТАНОВЛЕНИЕ СО2 В ГЕОТЕРМАЛЬНЫХ СИСТЕМАХ: ЩЕЛОЧНЫЕ ИСТОЧНИКИ
Температуры 40-100 'C, pH 10-11
Осаждение
ВОССТАНОВЛЕНИЕ СО2 В ГЕОТЕРМАЛЬНЫХ СИСТЕМАХ: ЩЕЛОЧНЫЕ ИСТОЧНИКИ
Температуры 40-100 'C, pH 10-11
Осаждение
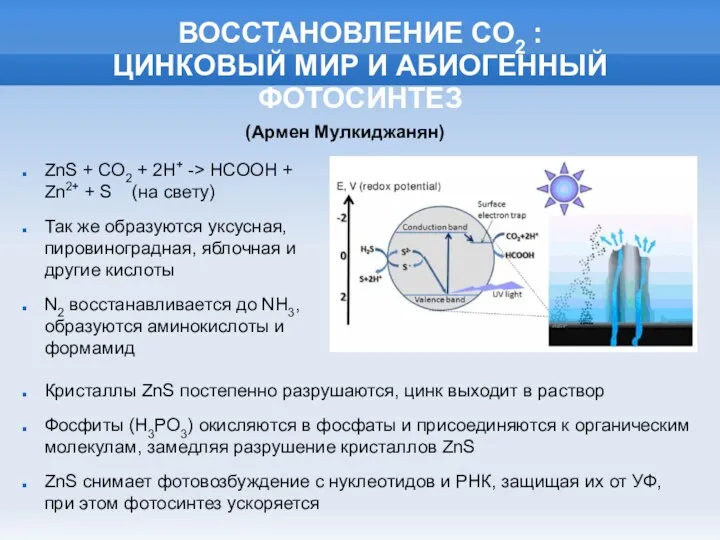
ВОССТАНОВЛЕНИЕ СО2 :
ЦИНКОВЫЙ МИР И АБИОГЕННЫЙ ФОТОСИНТЕЗ
Кристаллы ZnS постепенно разрушаются, цинк
ВОССТАНОВЛЕНИЕ СО2 :
ЦИНКОВЫЙ МИР И АБИОГЕННЫЙ ФОТОСИНТЕЗ
Кристаллы ZnS постепенно разрушаются, цинк

ЦИНКОВЫЙ МИР ИЛИ ЖЕЛЕЗНЫЙ МИР?
В рибозимах и древнейших белках встречается Zn,
ЦИНКОВЫЙ МИР ИЛИ ЖЕЛЕЗНЫЙ МИР?
В рибозимах и древнейших белках встречается Zn,
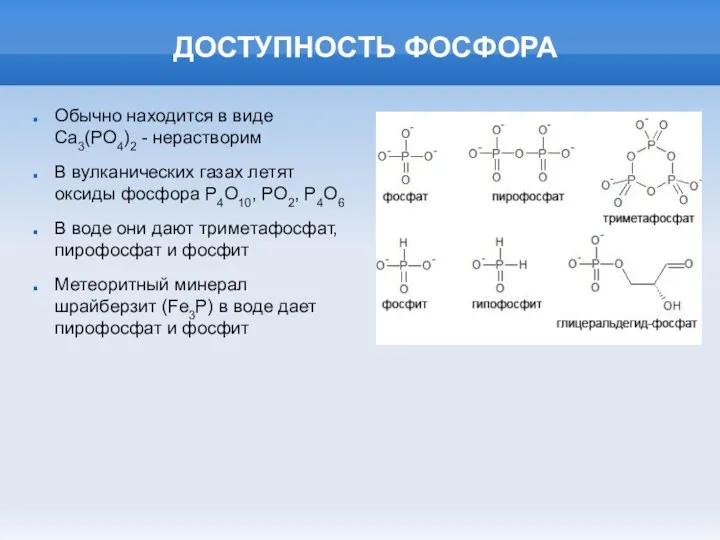
ДОСТУПНОСТЬ ФОСФОРА
Обычно находится в виде Ca3(PO4)2 - нерастворим
В вулканических газах летят
ДОСТУПНОСТЬ ФОСФОРА
Обычно находится в виде Ca3(PO4)2 - нерастворим
В вулканических газах летят

РАЗДЕЛЕНИЕ ВЕЩЕСТВ ПРИ КИПЕНИИ ВОДЫ В ГЕОТЕРМАЛЬНЫХ ИСТОЧНИКАХ
K, Mg, Zn, Mn,
РАЗДЕЛЕНИЕ ВЕЩЕСТВ ПРИ КИПЕНИИ ВОДЫ В ГЕОТЕРМАЛЬНЫХ ИСТОЧНИКАХ
K, Mg, Zn, Mn,

АВТОКАТАЛИТИЧЕСКИЕ РЕАКЦИИ
Автокаталитические реакции в биохимии:
Цикл Кальвина
Биосинтез большинства витаминов
Жизнь в целом с
АВТОКАТАЛИТИЧЕСКИЕ РЕАКЦИИ
Автокаталитические реакции в биохимии:
Цикл Кальвина
Биосинтез большинства витаминов
Жизнь в целом с

СИНТЕЗ НУКЛЕОТИДОВ
ПРОИСХОЖДЕНИЕ ХИРАЛЬНОЙ ЧИСТОТЫ
Лекция 5
СИНТЕЗ НУКЛЕОТИДОВ
ПРОИСХОЖДЕНИЕ ХИРАЛЬНОЙ ЧИСТОТЫ
Лекция 5
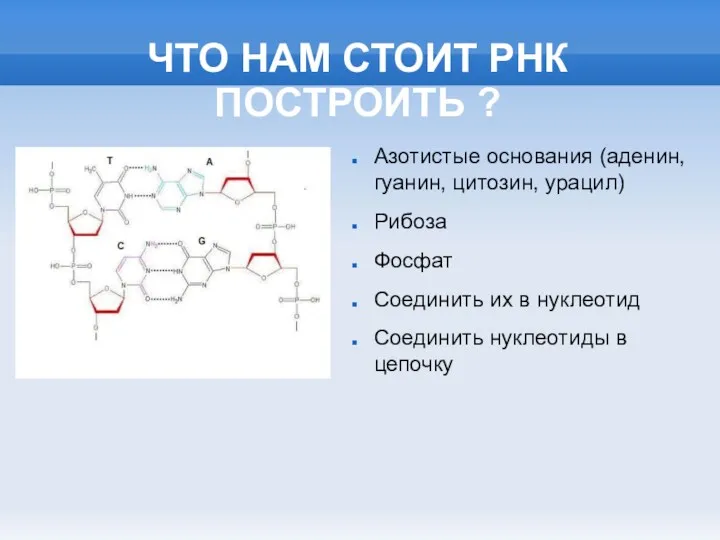
ЧТО НАМ СТОИТ РНК ПОСТРОИТЬ ?
Азотистые основания (аденин, гуанин, цитозин, урацил)
Рибоза
Фосфат
Соединить
ЧТО НАМ СТОИТ РНК ПОСТРОИТЬ ?
Азотистые основания (аденин, гуанин, цитозин, урацил)
Рибоза
Фосфат
Соединить
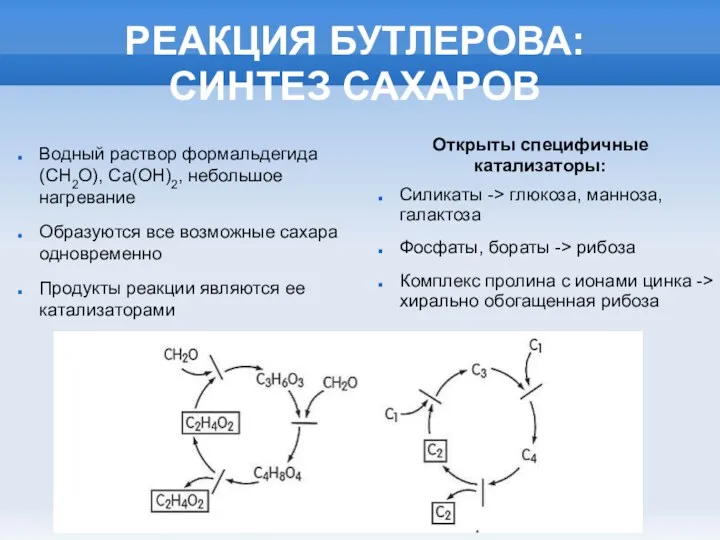
РЕАКЦИЯ БУТЛЕРОВА: СИНТЕЗ САХАРОВ
Водный раствор формальдегида (СН2О), Са(ОН)2, небольшое нагревание
Образуются все
РЕАКЦИЯ БУТЛЕРОВА: СИНТЕЗ САХАРОВ
Водный раствор формальдегида (СН2О), Са(ОН)2, небольшое нагревание
Образуются все

СИНТЕЗ АЗОТИСТЫХ ОСНОВАНИЙ
Аденин и гуанин - из HCN при замерзании водного
СИНТЕЗ АЗОТИСТЫХ ОСНОВАНИЙ
Аденин и гуанин - из HCN при замерзании водного
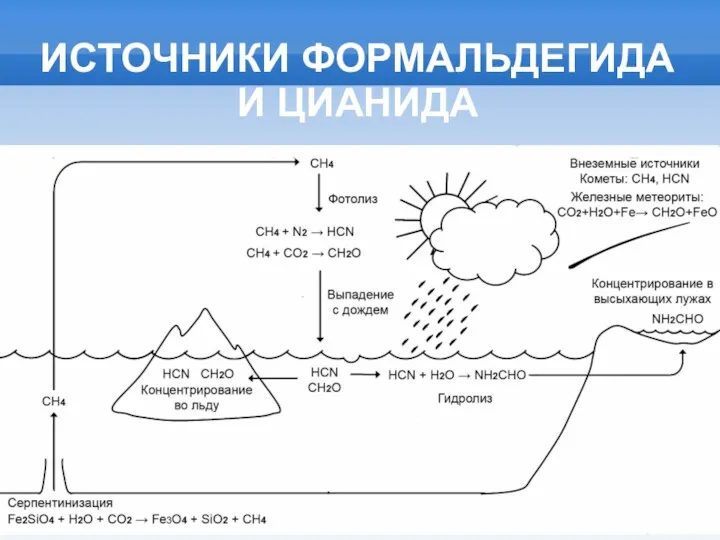
ИСТОЧНИКИ ФОРМАЛЬДЕГИДА И ЦИАНИДА
ИСТОЧНИКИ ФОРМАЛЬДЕГИДА И ЦИАНИДА

СИНТЕЗ НУКЛЕОТИДОВ
Аденин при УФ-облучении водного раствора присоединяет рибозу и до 3
СИНТЕЗ НУКЛЕОТИДОВ
Аденин при УФ-облучении водного раствора присоединяет рибозу и до 3
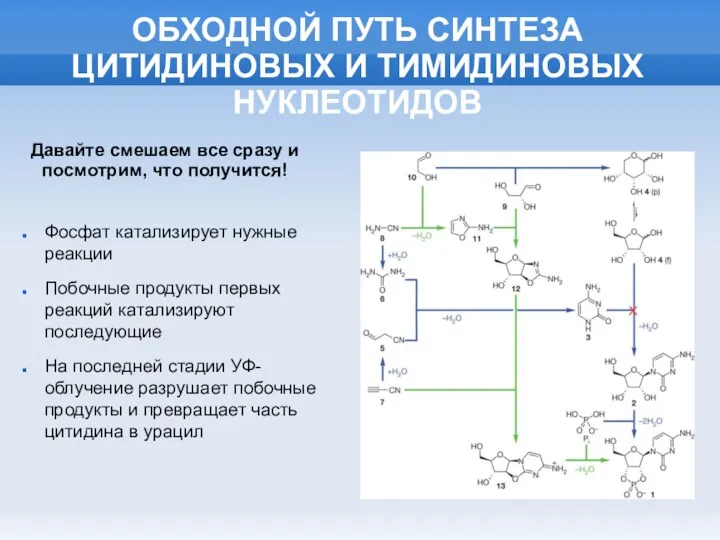
ОБХОДНОЙ ПУТЬ СИНТЕЗА ЦИТИДИНОВЫХ И ТИМИДИНОВЫХ НУКЛЕОТИДОВ
Фосфат катализирует нужные реакции
Побочные
ОБХОДНОЙ ПУТЬ СИНТЕЗА ЦИТИДИНОВЫХ И ТИМИДИНОВЫХ НУКЛЕОТИДОВ
Фосфат катализирует нужные реакции
Побочные
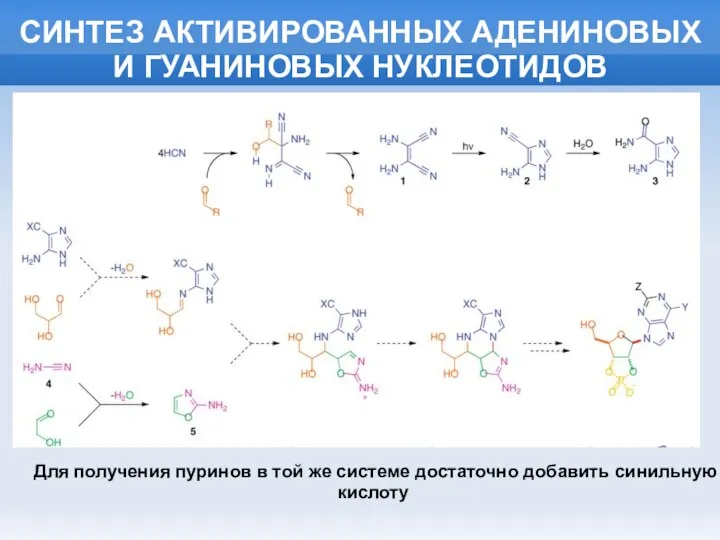
СИНТЕЗ АКТИВИРОВАННЫХ АДЕНИНОВЫХ И ГУАНИНОВЫХ НУКЛЕОТИДОВ
Для получения пуринов в той же
СИНТЕЗ АКТИВИРОВАННЫХ АДЕНИНОВЫХ И ГУАНИНОВЫХ НУКЛЕОТИДОВ
Для получения пуринов в той же
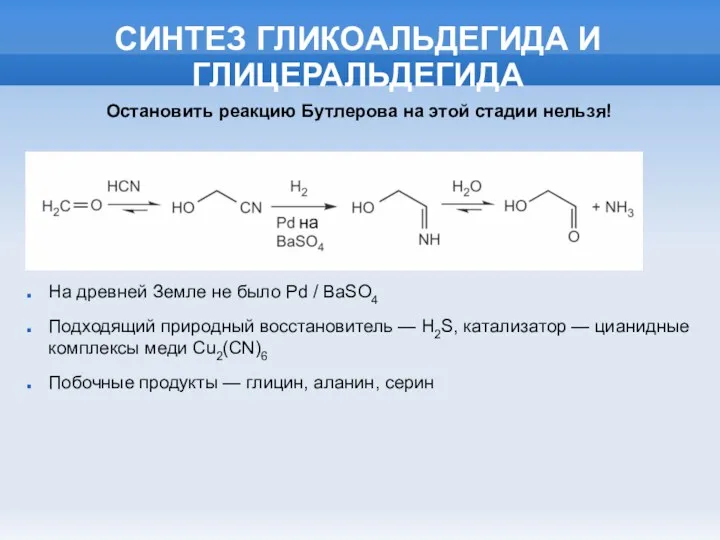
СИНТЕЗ ГЛИКОАЛЬДЕГИДА И ГЛИЦЕРАЛЬДЕГИДА
На древней Земле не было Pd / BaSO4
СИНТЕЗ ГЛИКОАЛЬДЕГИДА И ГЛИЦЕРАЛЬДЕГИДА
На древней Земле не было Pd / BaSO4
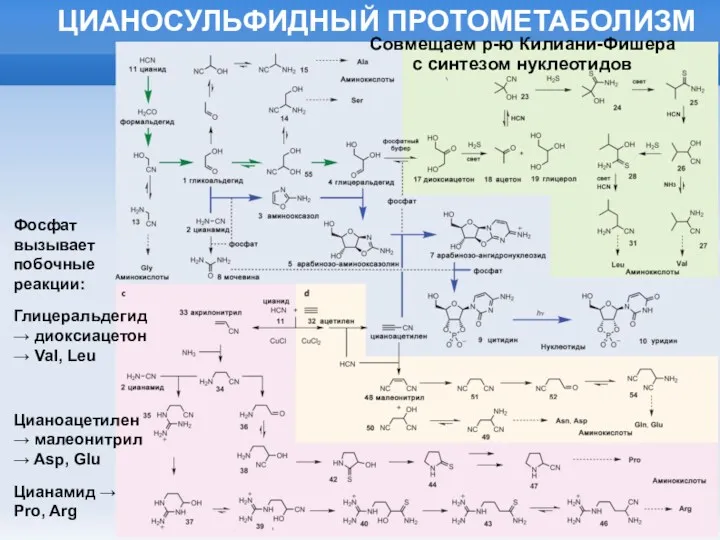
ЦИАНОСУЛЬФИДНЫЙ ПРОТОМЕТАБОЛИЗМ
Фосфат вызывает побочные реакции:
Глицеральдегид → диоксиацетон → Val, Leu
Цианоацетилен
ЦИАНОСУЛЬФИДНЫЙ ПРОТОМЕТАБОЛИЗМ
Фосфат вызывает побочные реакции:
Глицеральдегид → диоксиацетон → Val, Leu
Цианоацетилен

МЕХАНИЗМЫ ПОЯВЛЕНИЯ ХИРАЛЬНОЙ ЧИСТОТЫ
В метеоритной органике есть преобладание левых (L) аминокислот
МЕХАНИЗМЫ ПОЯВЛЕНИЯ ХИРАЛЬНОЙ ЧИСТОТЫ
В метеоритной органике есть преобладание левых (L) аминокислот
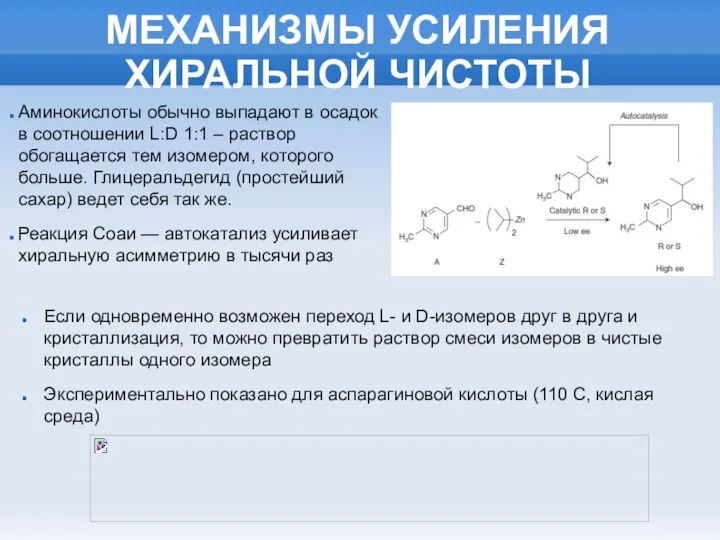
МЕХАНИЗМЫ УСИЛЕНИЯ ХИРАЛЬНОЙ ЧИСТОТЫ
Аминокислоты обычно выпадают в осадок в соотношении L:D
МЕХАНИЗМЫ УСИЛЕНИЯ ХИРАЛЬНОЙ ЧИСТОТЫ
Аминокислоты обычно выпадают в осадок в соотношении L:D
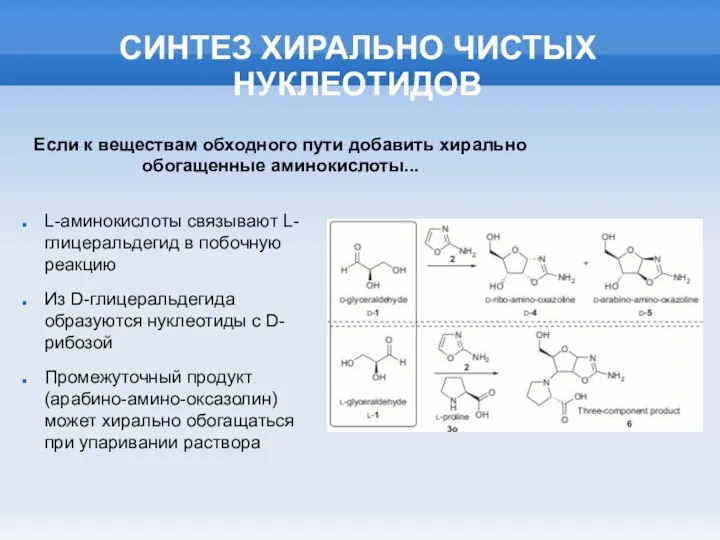
СИНТЕЗ ХИРАЛЬНО ЧИСТЫХ НУКЛЕОТИДОВ
L-аминокислоты связывают L-глицеральдегид в побочную реакцию
Из D-глицеральдегида
СИНТЕЗ ХИРАЛЬНО ЧИСТЫХ НУКЛЕОТИДОВ
L-аминокислоты связывают L-глицеральдегид в побочную реакцию
Из D-глицеральдегида

КОПИРОВАНИЕ РНК
НАЧАЛО РНК-МИРА
Лекция 6
КОПИРОВАНИЕ РНК
НАЧАЛО РНК-МИРА
Лекция 6

БЕЗМАТРИЧНАЯ ПОЛИМЕРИЗАЦИЯ НУКЛЕОТИДОВ В РНК
На поверхности минералов (глины, сульфиды металлов) из
БЕЗМАТРИЧНАЯ ПОЛИМЕРИЗАЦИЯ НУКЛЕОТИДОВ В РНК
На поверхности минералов (глины, сульфиды металлов) из

РИБОЗИМЫ — РНК-ПОЛИМЕРАЗЫ
Активные рибозимы-полимеразы состоят из 150-200 нуклеотидов
Лучшая полимераза tC19Z во
РИБОЗИМЫ — РНК-ПОЛИМЕРАЗЫ
Активные рибозимы-полимеразы состоят из 150-200 нуклеотидов
Лучшая полимераза tC19Z во
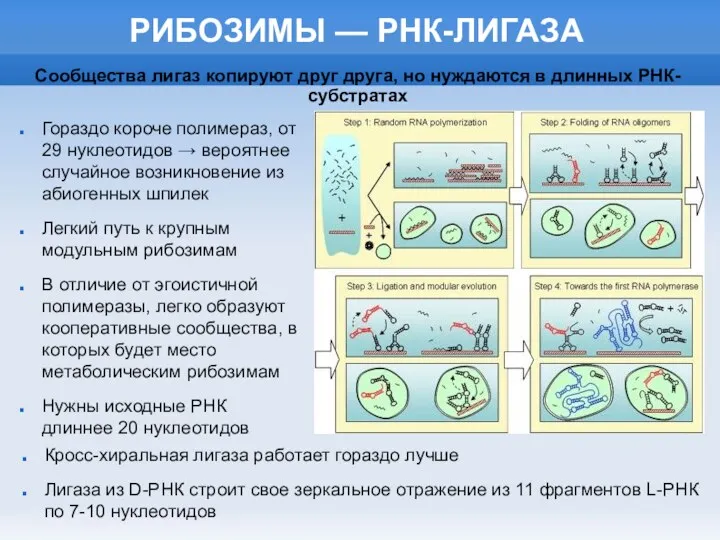
РИБОЗИМЫ — РНК-ЛИГАЗА
Гораздо короче полимераз, от 29 нуклеотидов → вероятнее случайное
РИБОЗИМЫ — РНК-ЛИГАЗА
Гораздо короче полимераз, от 29 нуклеотидов → вероятнее случайное

АБИОГЕННАЯ РЕПЛИКАЦИЯ РНК
При высокой концентрации Mg++ короткие РНК реплицируются сами
Копируется до
АБИОГЕННАЯ РЕПЛИКАЦИЯ РНК
При высокой концентрации Mg++ короткие РНК реплицируются сами
Копируется до

НАКОПЛЕНИЕ РНК В ТЕПЛОВОЙ ЛОВУШКЕ
Конвекция и термофорез в капиллярах концентрируют РНК
НАКОПЛЕНИЕ РНК В ТЕПЛОВОЙ ЛОВУШКЕ
Конвекция и термофорез в капиллярах концентрируют РНК
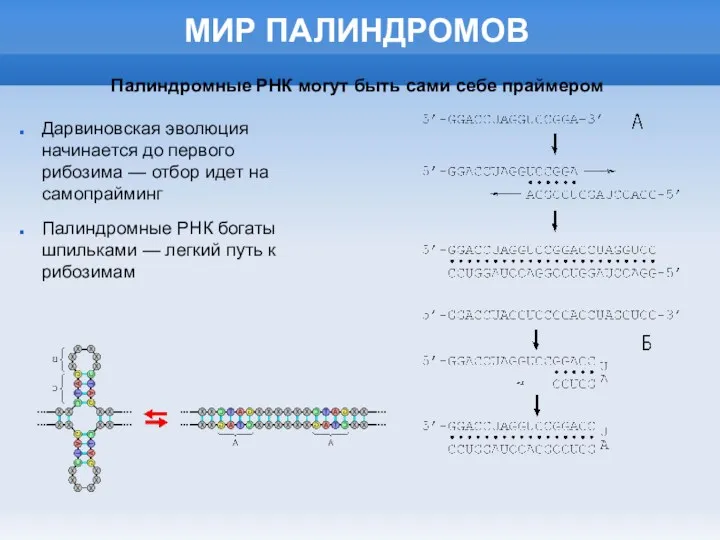
МИР ПАЛИНДРОМОВ
Дарвиновская эволюция начинается до первого рибозима — отбор идет на
МИР ПАЛИНДРОМОВ
Дарвиновская эволюция начинается до первого рибозима — отбор идет на
 Вегетативті көбею
Вегетативті көбею Девонский период палеозойской эры
Девонский период палеозойской эры Многообразие цветковых растений. Классы покрытосеменных. Семейства. Роль растений в природе и жизни человека
Многообразие цветковых растений. Классы покрытосеменных. Семейства. Роль растений в природе и жизни человека Обмен веществ и энергией
Обмен веществ и энергией Органы цветковых растений
Органы цветковых растений Витамины. Классификация витаминов
Витамины. Классификация витаминов Особенности желудочного пищеварения у жвачных
Особенности желудочного пищеварения у жвачных Тип кишечнополостные (пресноводная гидра)
Тип кишечнополостные (пресноводная гидра) Экспертиза качества молока
Экспертиза качества молока Утворення торфа та кам’яного вугілля
Утворення торфа та кам’яного вугілля Пестрая ситовая (ямчато-волокнистая) гниль корней. Корневая губка
Пестрая ситовая (ямчато-волокнистая) гниль корней. Корневая губка Функциональная структура биосферы
Функциональная структура биосферы Тұқым қуалаушылықтың молекулярлық негіздері
Тұқым қуалаушылықтың молекулярлық негіздері Рефлекс. Рефлекторная дуга. Спинной мозг
Рефлекс. Рефлекторная дуга. Спинной мозг Организация внеклассной работы по биологии. Разновозрастной биологический кружок
Организация внеклассной работы по биологии. Разновозрастной биологический кружок Анатомо-физиологические особенности человека в подростковом возрасте
Анатомо-физиологические особенности человека в подростковом возрасте Здоровое питание
Здоровое питание Круговорот веществ в природе
Круговорот веществ в природе Овчарки со всего мира. Фотографии
Овчарки со всего мира. Фотографии Історія виноробства. Класифікація вин
Історія виноробства. Класифікація вин Unusual animals
Unusual animals Белки
Белки Кровообращение как функциональная система. Анатомия сердца и сосудов. Характерные черты строения и функционирования сердечной мышцы
Кровообращение как функциональная система. Анатомия сердца и сосудов. Характерные черты строения и функционирования сердечной мышцы Публичная презентация 2014 год
Публичная презентация 2014 год Бактериофаги. Вирусы бактерий
Бактериофаги. Вирусы бактерий Фотосинтез. 9 класс
Фотосинтез. 9 класс Тварина - живий організм
Тварина - живий організм Доза і основні одиниці вимірювання іонізуючого випромінювання. Основні типи дозових залежностей в радіобіології
Доза і основні одиниці вимірювання іонізуючого випромінювання. Основні типи дозових залежностей в радіобіології