- Информационные макромолекулы

Содержание
- 2. Белки и нуклеиновые кислоты являются информаци-онными макромолекулами; играют огромную роль в обес-печении способности организма существовать во
- 3. Белки и нуклеиновые кислоты Белки и нуклеиновые кислоты – это органические ве-щества - биополимеры, высокомолекулярные соединения
- 4. Белки называют протеинами Состав белков Углерод азот 50-54% кислород 15-18% водород 20-23% сера 6-8% 2.5%
- 5. Аминокислоты Общее число аминокислот - 300; из них в состав белков входит только 20. Их называют
- 6. Образование пептидной связи Н О O │ ║ ║ N —CH— C — OH + H
- 7. Пептидная связь характеризуется следующими свойствами: 1. 4 атома пептидной связи лежат в одной плоскости (рис.7а); 2.
- 8. Пептидная связь характеризуется следующими свойствами: 1. 4 атома пептидной связи лежат в одной плоскости (рис.7а); 2.
- 9. Стереометрическая модель пептидной связи
- 10. Пространственная модель вращения аминокислотных остатков
- 11. Пептиды Соединения, в которых аминокислоты соединены пептидными связями, называется пептидами: олигопептиды (содержат до 10 аминокислот), полипептиды
- 12. Первичная структура полипептида
- 13. Α-спираль вторичной структуры полипептида В α-спирали NH-группа одного остатка аминокислоты соединяется водородной связью с СО-группой пятого
- 14. β-складчатый слой вторичной структуры полипептида β-структура (β-складчатый слой)) формируется за счет образования множества водородных связей между
- 15. Третичная структура белка Третичная структура белка – это трехмерная прост -ранственная структура, образующаяся за счет взаимо-
- 16. Третичная структура белка – это трехмер-ная пространственная структура, образую-щаяся за счет взаимодействия между ра-дикалами аминокислот, находящимися
- 18. Белковая глобула
- 19. Супервторичная структура белка Белок с мотивом «спираль-виток-спираль»: а - α-спираль белка, содержащая 3 домена; b -
- 20. . А – мотив суперструктуры белка «лейциновая застежка-молния» - между α-спиральными участками двух белков; Б –
- 21. Четвертичная структура белка Четвертичная структура гемоглобина Субъединичная структура глутаматсинтетазы: а – протомер, б – мультимер белка
- 22. Фолдинг белка Первичная структура белка формируется в результате трансляции белка. Пептидная цепь претерпевает пространственные изменения, приводящие
- 23. Первичная структура белка формируется в результате трансляции белка. Пептидная цепь претерпевает пространственные изменения, приводящие к ее
- 24. Классификация белков Белки можно классифицировать: по форме (глобулярные и фибриллярные), по молекулярной массе (низко-, высокомолекулярные и
- 25. Некоторые сложные белки
- 26. Виды азотистых оснований
- 27. Пуриновые и пиримидиновые основания Строение пентоз
- 28. Нуклеиновые кислоты – высокомолекулярные органические соединения - биополимеры, мономерами которых является нуклеотиды. Нуклеотиды являются нуклеозидфосфатами и
- 29. В зависимости от характера пентозы (рибоза или дезоксирибоза) в нуклеотиде различают рибонуклеозидфосфаты и дезоксинуклеозидмонофос - фаты.
- 30. 5′С О 5′С О │ 4′С С1′― аз.осн.1 4′С С1′ 3′С С2′ 3′С С2′ │ ОН
- 31. Схема полинуклеотидной цепи с указанием направления роста
- 32. Виды нуклеиновых кислот ДНК РНК ( дезоксиробонуклеиновая) (рибонуклеиновая) А-форма Z-форма мРНК тРНК рРНК В-форма мя-РНК рибозимы
- 33. Вторичная структура ДНК Молекула ДНК построена из двух полинуклеотидных цепей, ориентированных антипараллельно, Цепи соединены водородными связями
- 34. Пространственная структура молекулы ДНК
- 35. А – модели Z-формы (1) и В-формы (2) молекулы ДНК ; Б - схемы двойной спирали
- 36. . Схема расположения функциональных участков на молекуле мРНК Вторичная структура м-РНК, Схема третичной структуры мРНК (А)
- 37. Р-Г-Г-Г-Ц-Г-У-Г-У-МетГ-Г-Ц-Г-Ц-Г-У-А-Г-ДиГУ-А-Г-Ц-Г-Ц-ДиМеГ-Ц-У-Ц-Ц-Ц- ДиМетГ-Ц-У-Ц-Ц-Ц-У-У-Г-Ц-МетИ-ψ-Г-Г-Г-А-А-Г-У-Ц-Ц-Г-Г-Т-ψ-Ц-Г-А-У-У-Ц-Ц- Г-Г-А-Ц-У-Ц-Г-У-Ц-Ц-А-Ц-Ц-А-ОН Первичная структура аланиновой тРНК (по Р.Холли): минорные основания - ДиГУ(дигидроуридин), ДиМетГ (диметилгуанин), МетГ(
- 38. Первичная структура молекулы 5.8 рРНК Вторичная структура молекулы рРНК Helicobaster pylori Третичная структура молекулы рРНК в
- 40. Скачать презентацию
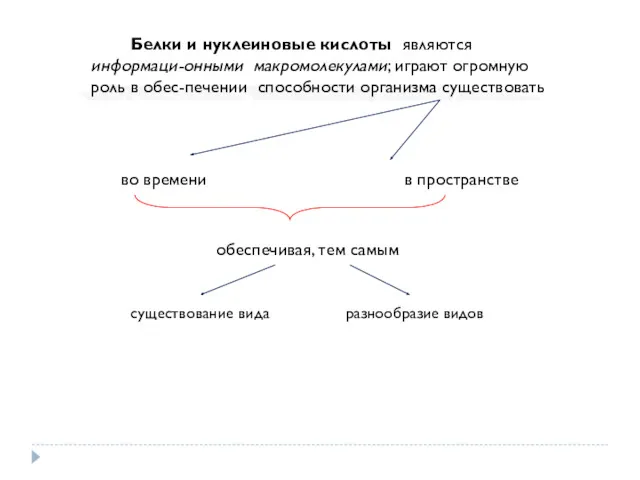
Белки и нуклеиновые кислоты являются информаци-онными макромолекулами; играют огромную роль
Белки и нуклеиновые кислоты являются информаци-онными макромолекулами; играют огромную роль

Белки и нуклеиновые кислоты
Белки и нуклеиновые кислоты – это органические
Белки и нуклеиновые кислоты
Белки и нуклеиновые кислоты – это органические
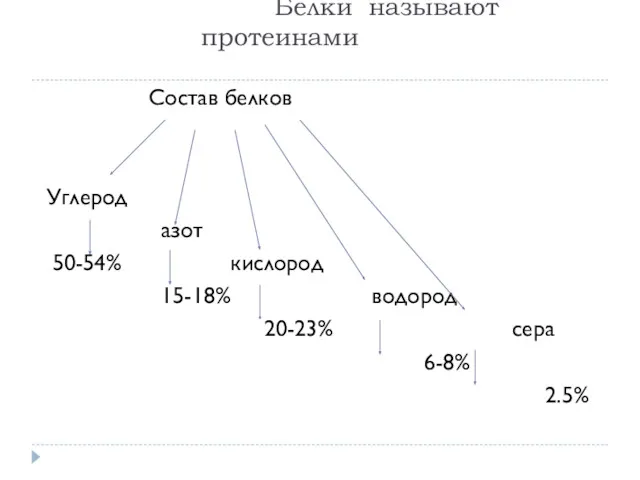
Белки называют протеинами
Состав белков
Углерод
азот
50-54% кислород
Белки называют протеинами
Состав белков
Углерод
азот
50-54% кислород
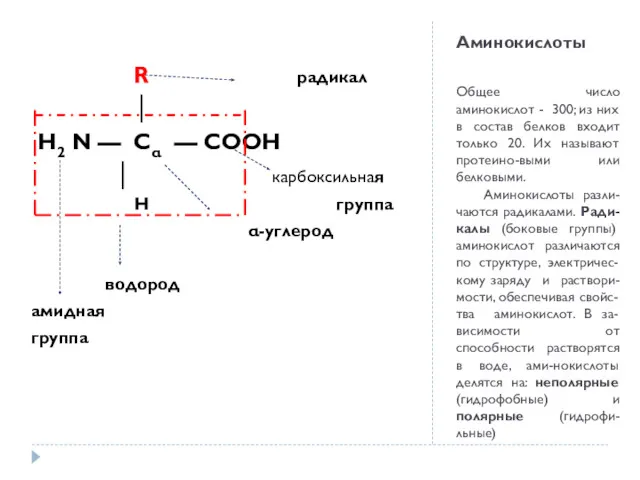
Аминокислоты
Общее число аминокислот - 300; из них в состав белков входит
Аминокислоты
Общее число аминокислот - 300; из них в состав белков входит
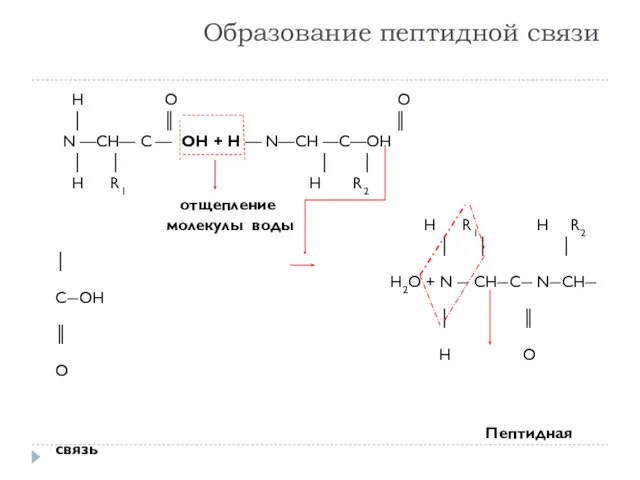
Образование пептидной связи
Н О O
│ ║ ║
Образование пептидной связи
Н О O
│ ║ ║

Пептидная связь характеризуется следующими свойствами:
1. 4 атома пептидной связи лежат
Пептидная связь характеризуется следующими свойствами:
1. 4 атома пептидной связи лежат

Пептидная связь характеризуется следующими свойствами:
1. 4 атома пептидной связи лежат
Пептидная связь характеризуется следующими свойствами:
1. 4 атома пептидной связи лежат
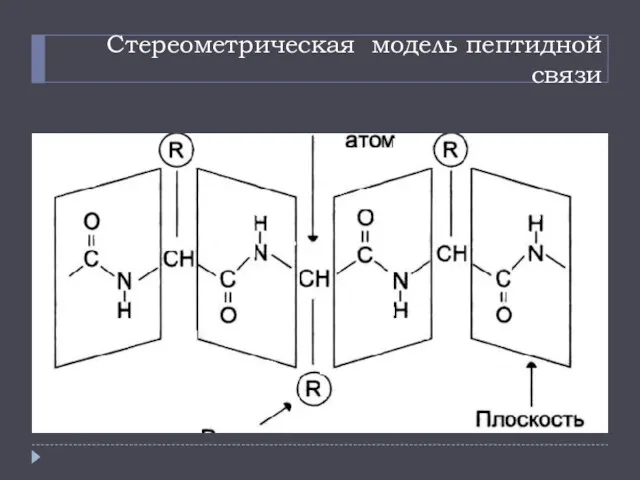
Стереометрическая модель пептидной связи
Стереометрическая модель пептидной связи
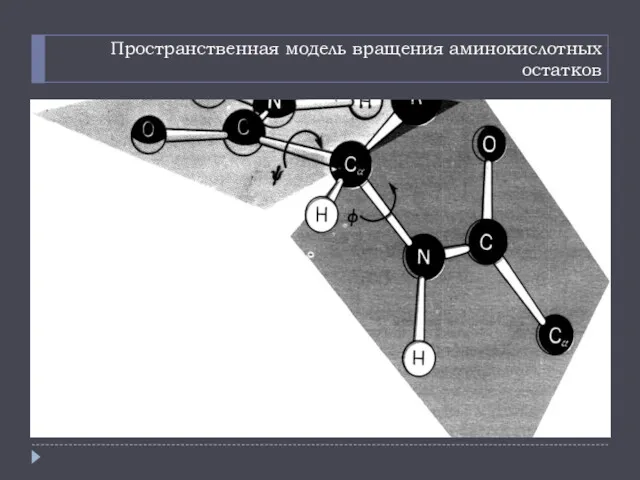
Пространственная модель вращения аминокислотных остатков
Пространственная модель вращения аминокислотных остатков

Пептиды
Соединения, в которых аминокислоты соединены пептидными связями, называется пептидами:
олигопептиды (содержат
Пептиды
Соединения, в которых аминокислоты соединены пептидными связями, называется пептидами:
олигопептиды (содержат
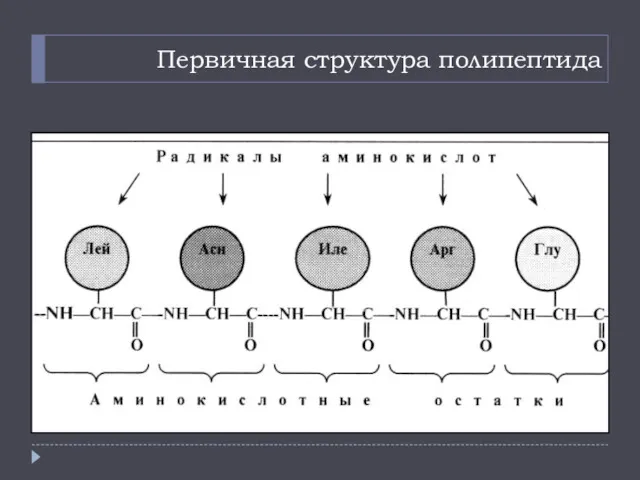
Первичная структура полипептида
Первичная структура полипептида
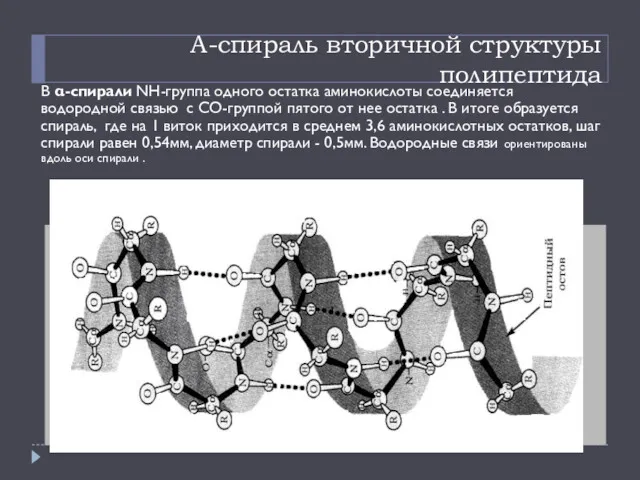
Α-спираль вторичной структуры полипептида
В α-спирали NH-группа одного остатка аминокислоты соединяется водородной
Α-спираль вторичной структуры полипептида
В α-спирали NH-группа одного остатка аминокислоты соединяется водородной
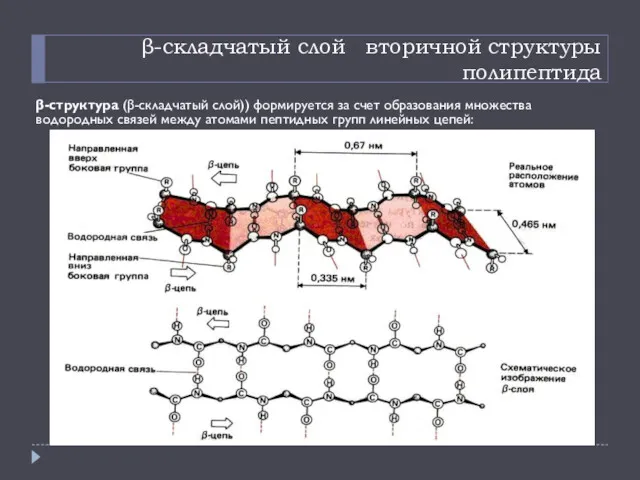
β-складчатый слой вторичной структуры полипептида
β-структура (β-складчатый слой)) формируется за счет образования
β-складчатый слой вторичной структуры полипептида
β-структура (β-складчатый слой)) формируется за счет образования

Третичная структура белка
Третичная структура белка – это трехмерная прост
Третичная структура белка
Третичная структура белка – это трехмерная прост
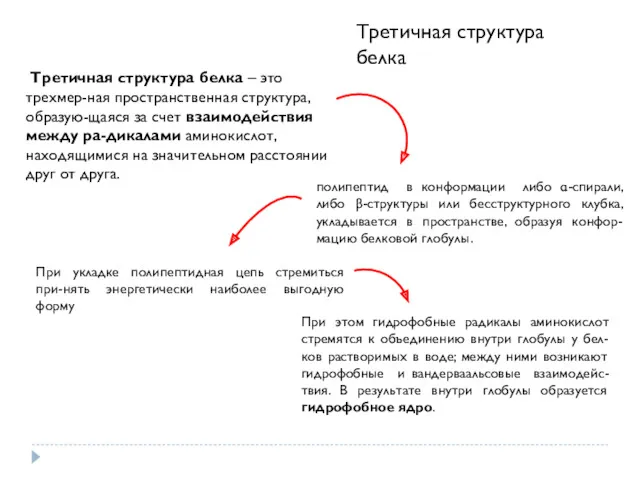
Третичная структура белка – это трехмер-ная пространственная структура, образую-щаяся за
Третичная структура белка – это трехмер-ная пространственная структура, образую-щаяся за
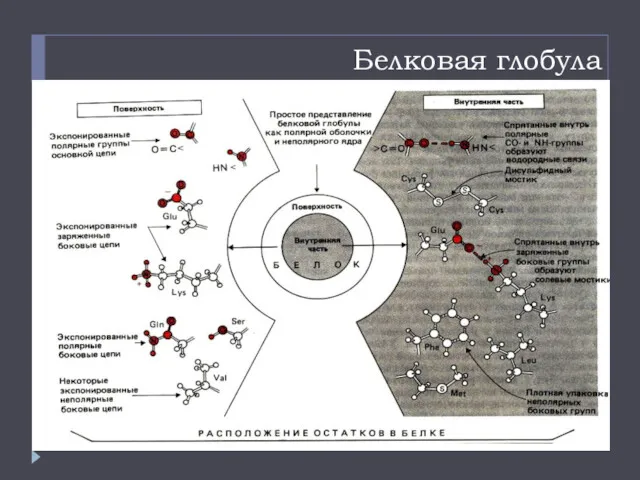
Белковая глобула
Белковая глобула
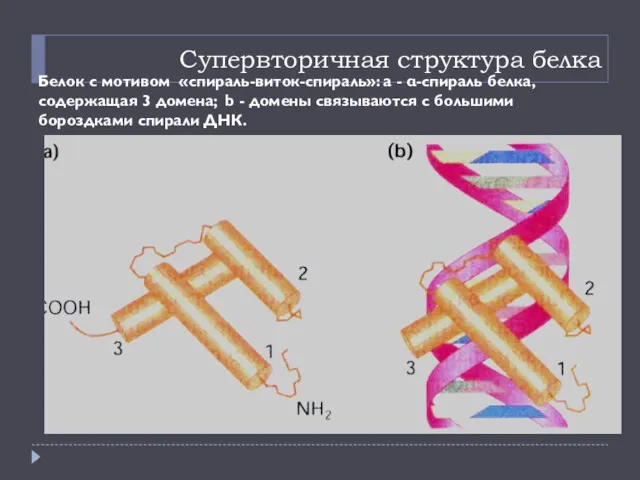
Супервторичная структура белка
Белок с мотивом «спираль-виток-спираль»: а - α-спираль белка, содержащая
Супервторичная структура белка
Белок с мотивом «спираль-виток-спираль»: а - α-спираль белка, содержащая
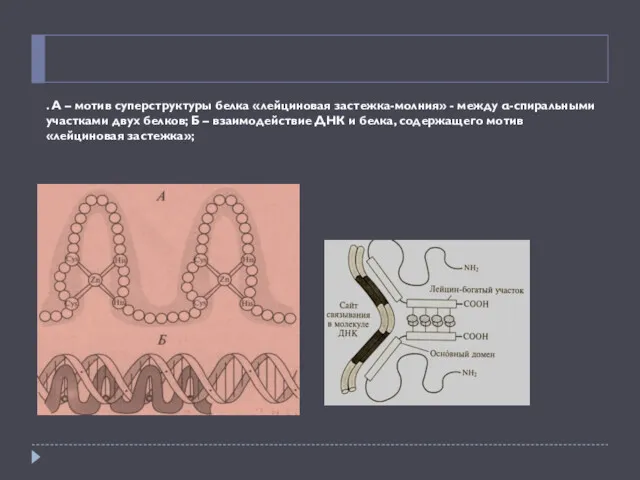
. А – мотив суперструктуры белка «лейциновая застежка-молния» - между
. А – мотив суперструктуры белка «лейциновая застежка-молния» - между
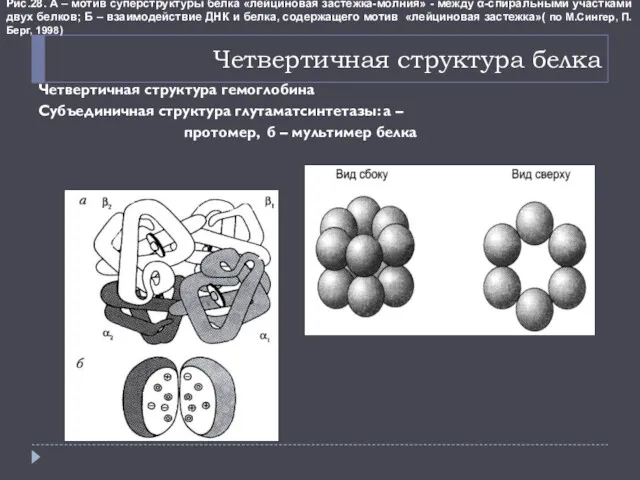
Четвертичная структура белка
Четвертичная структура гемоглобина
Субъединичная структура глутаматсинтетазы: а –
протомер,
Четвертичная структура белка
Четвертичная структура гемоглобина
Субъединичная структура глутаматсинтетазы: а –
протомер,

Фолдинг белка
Первичная структура белка формируется в результате трансляции белка.
Фолдинг белка
Первичная структура белка формируется в результате трансляции белка.
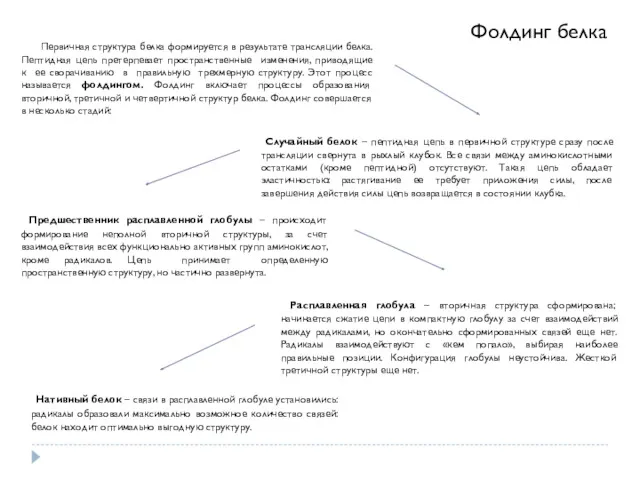
Первичная структура белка формируется в результате трансляции белка. Пептидная цепь
Первичная структура белка формируется в результате трансляции белка. Пептидная цепь

Классификация белков
Белки можно классифицировать:
по форме (глобулярные и фибриллярные),
по
Классификация белков
Белки можно классифицировать:
по форме (глобулярные и фибриллярные),
по

Некоторые сложные белки
Некоторые сложные белки
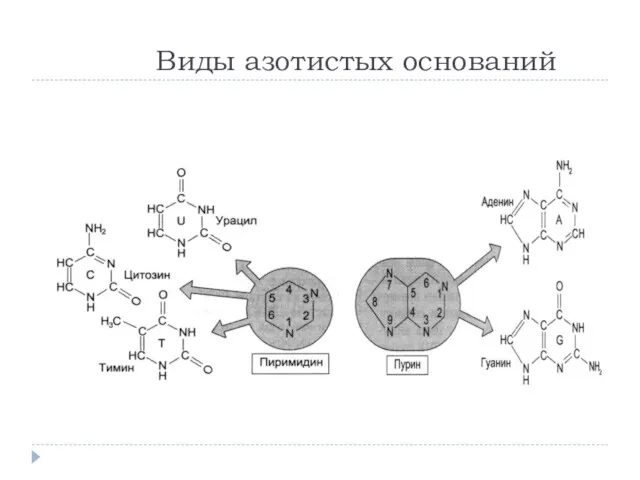
Виды азотистых оснований
Виды азотистых оснований
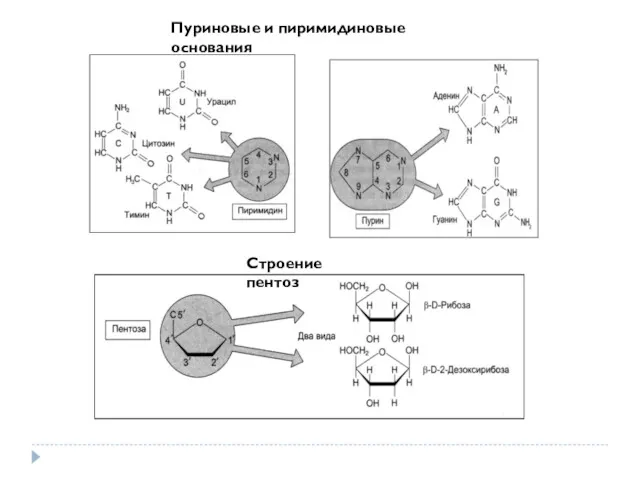
Пуриновые и пиримидиновые основания
Строение пентоз
Пуриновые и пиримидиновые основания
Строение пентоз
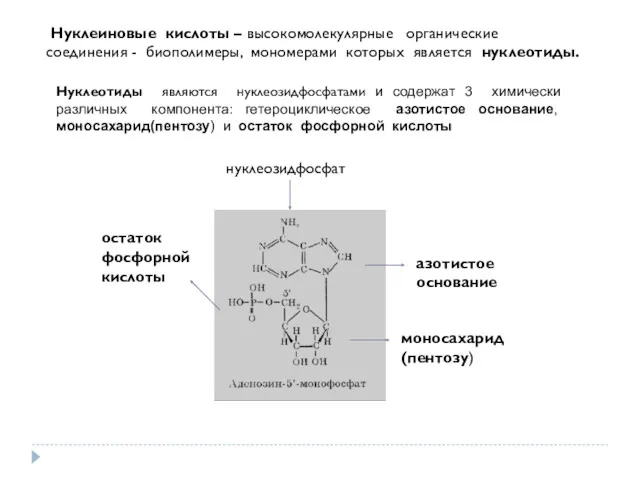
Нуклеиновые кислоты – высокомолекулярные органические соединения - биополимеры, мономерами которых
Нуклеиновые кислоты – высокомолекулярные органические соединения - биополимеры, мономерами которых
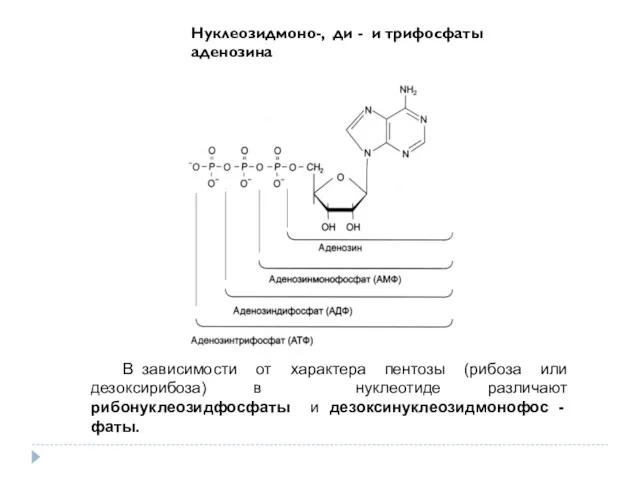
В зависимости от характера пентозы (рибоза или дезоксирибоза) в нуклеотиде различают
В зависимости от характера пентозы (рибоза или дезоксирибоза) в нуклеотиде различают
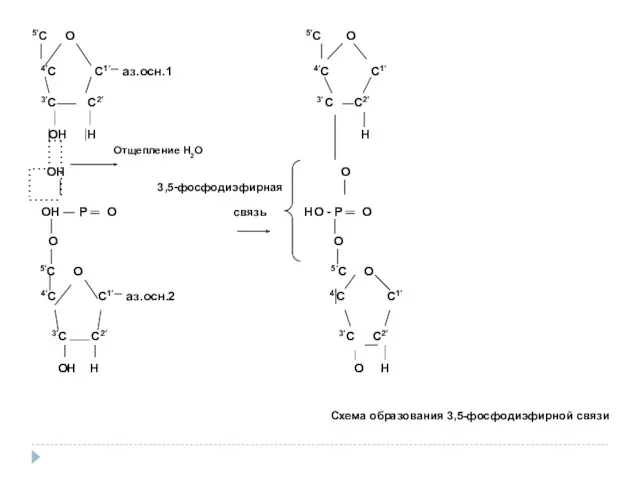
5′С О 5′С О
│
4′С С1′― аз.осн.1 4′С С1′
5′С О 5′С О
│
4′С С1′― аз.осн.1 4′С С1′
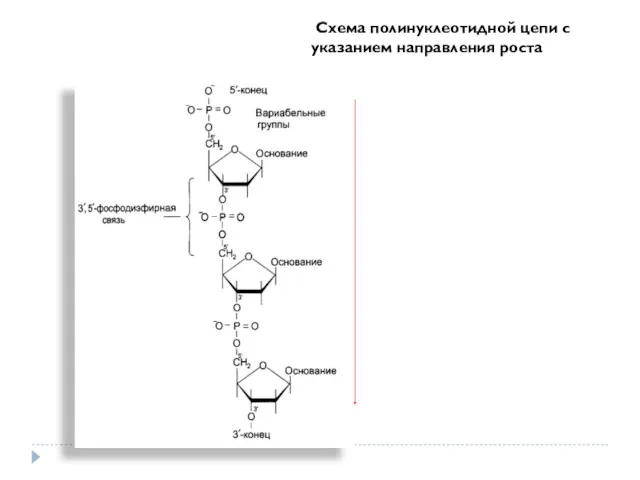
Схема полинуклеотидной цепи с указанием направления роста
Схема полинуклеотидной цепи с указанием направления роста
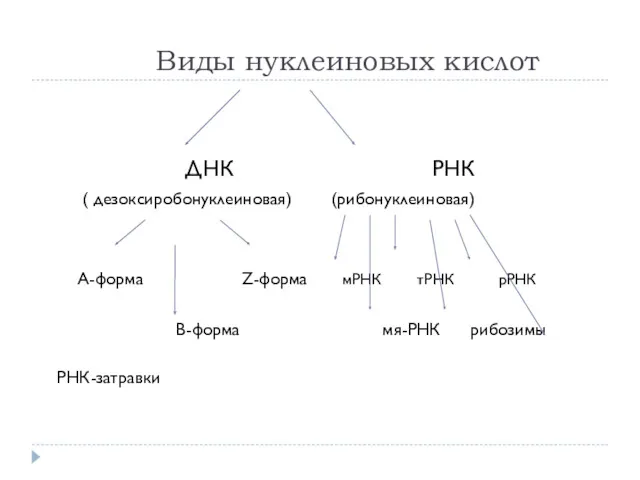
Виды нуклеиновых кислот
ДНК РНК
( дезоксиробонуклеиновая) (рибонуклеиновая)
А-форма
Виды нуклеиновых кислот
ДНК РНК
( дезоксиробонуклеиновая) (рибонуклеиновая)
А-форма
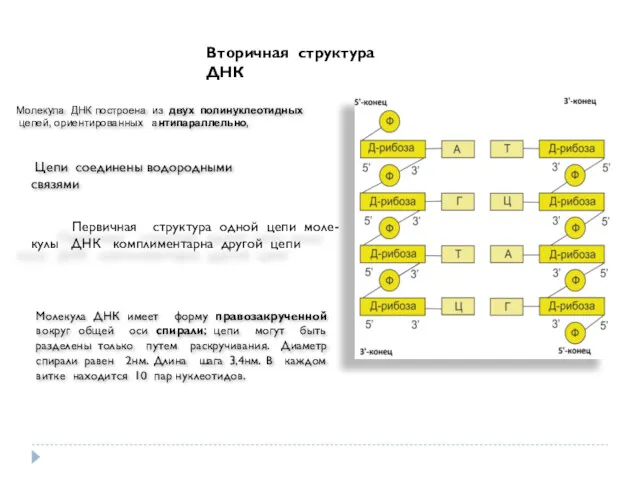
Вторичная структура ДНК
Молекула ДНК построена из двух полинуклеотидных
цепей, ориентированных антипараллельно,
Вторичная структура ДНК
Молекула ДНК построена из двух полинуклеотидных
цепей, ориентированных антипараллельно,
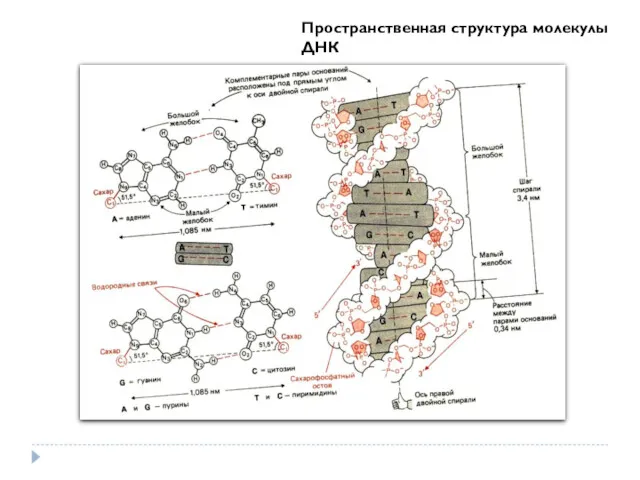
Пространственная структура молекулы ДНК
Пространственная структура молекулы ДНК
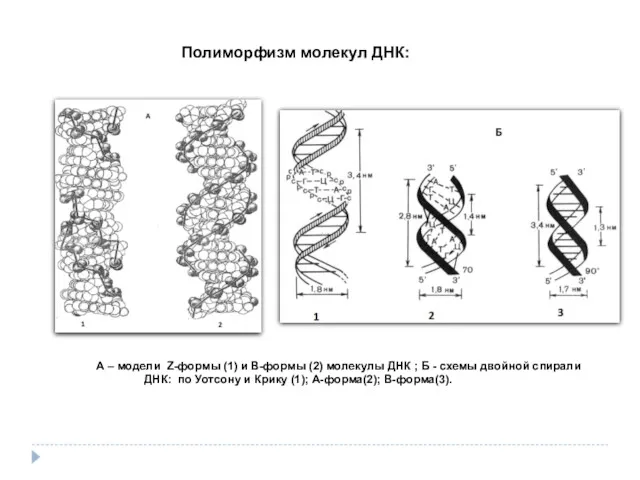
А – модели Z-формы (1) и В-формы (2) молекулы ДНК ;
А – модели Z-формы (1) и В-формы (2) молекулы ДНК ;
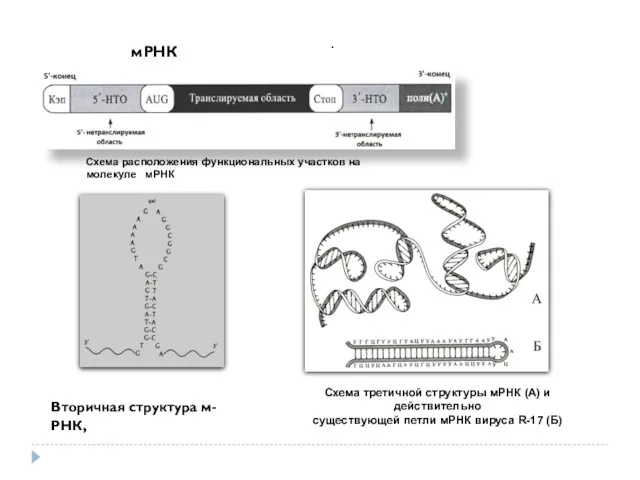
.
Схема расположения функциональных участков на молекуле мРНК
Вторичная структура м-РНК,
Схема третичной
.
Схема расположения функциональных участков на молекуле мРНК
Вторичная структура м-РНК,
Схема третичной
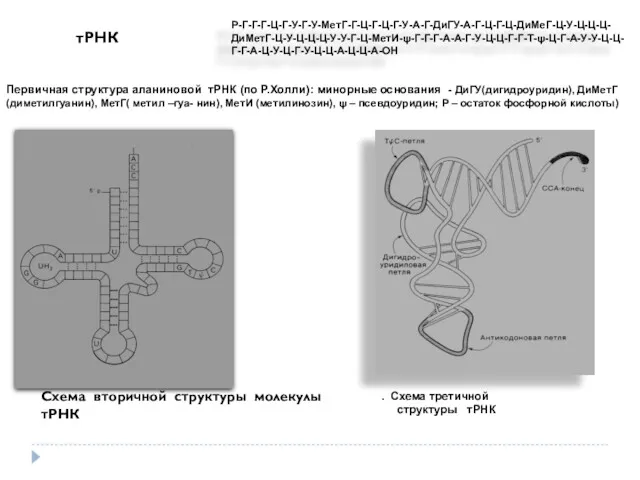
Р-Г-Г-Г-Ц-Г-У-Г-У-МетГ-Г-Ц-Г-Ц-Г-У-А-Г-ДиГУ-А-Г-Ц-Г-Ц-ДиМеГ-Ц-У-Ц-Ц-Ц-
ДиМетГ-Ц-У-Ц-Ц-Ц-У-У-Г-Ц-МетИ-ψ-Г-Г-Г-А-А-Г-У-Ц-Ц-Г-Г-Т-ψ-Ц-Г-А-У-У-Ц-Ц-
Г-Г-А-Ц-У-Ц-Г-У-Ц-Ц-А-Ц-Ц-А-ОН
Первичная структура аланиновой тРНК (по Р.Холли): минорные основания - ДиГУ(дигидроуридин), ДиМетГ
Р-Г-Г-Г-Ц-Г-У-Г-У-МетГ-Г-Ц-Г-Ц-Г-У-А-Г-ДиГУ-А-Г-Ц-Г-Ц-ДиМеГ-Ц-У-Ц-Ц-Ц-
ДиМетГ-Ц-У-Ц-Ц-Ц-У-У-Г-Ц-МетИ-ψ-Г-Г-Г-А-А-Г-У-Ц-Ц-Г-Г-Т-ψ-Ц-Г-А-У-У-Ц-Ц-
Г-Г-А-Ц-У-Ц-Г-У-Ц-Ц-А-Ц-Ц-А-ОН
Первичная структура аланиновой тРНК (по Р.Холли): минорные основания - ДиГУ(дигидроуридин), ДиМетГ
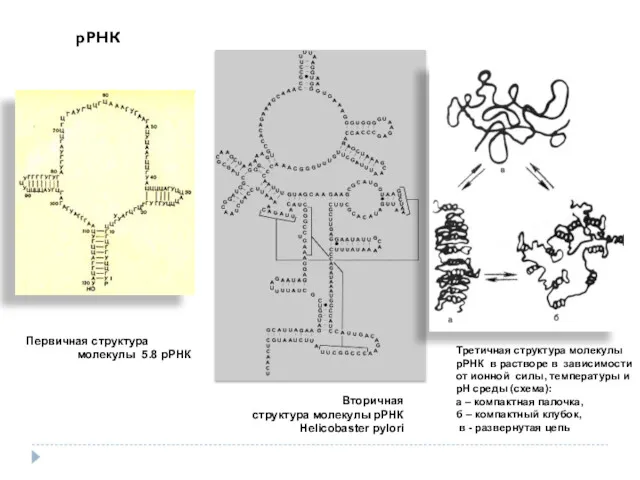
Первичная структура
молекулы 5.8 рРНК
Вторичная
структура молекулы рРНК
Helicobaster pylori
Третичная структура
Первичная структура
молекулы 5.8 рРНК
Вторичная
структура молекулы рРНК
Helicobaster pylori
Третичная структура
 Репликация. Основы
Репликация. Основы Насекомые – вредители сада
Насекомые – вредители сада Птахи, які користуються знаряддями праці
Птахи, які користуються знаряддями праці Растения-хищники
Растения-хищники Этические проблемы использования животных
Этические проблемы использования животных Генная инженерия
Генная инженерия Мышцы шеи
Мышцы шеи Будова, властивості, характеристика, класифікація вірусів
Будова, властивості, характеристика, класифікація вірусів Головной мозг - строение
Головной мозг - строение Хвостатые земноводные
Хвостатые земноводные Возврат органики в почву. Грядка Бублика
Возврат органики в почву. Грядка Бублика Растениеводство. Классификация севооборотов и их основные звенья
Растениеводство. Классификация севооборотов и их основные звенья Органы и системы органов
Органы и системы органов Строение и развитие органов дыхательной системы
Строение и развитие органов дыхательной системы Биотехнология в животноводстве
Биотехнология в животноводстве Экологиялық факторлар
Экологиялық факторлар Уход за морской свинкой
Уход за морской свинкой Влияние сроков посева на рост и развитие сеянцев сосны и ели с закрытой корневой системой
Влияние сроков посева на рост и развитие сеянцев сосны и ели с закрытой корневой системой Растения-хищники
Растения-хищники Физика и анатомия музыки
Физика и анатомия музыки Мейоз
Мейоз Насекомые. Отгадайте загадки
Насекомые. Отгадайте загадки Биология кошек
Биология кошек Презентация по теме: Особенности высшей нервной деятельности человека.
Презентация по теме: Особенности высшей нервной деятельности человека. Хищные животные Хабаровского края
Хищные животные Хабаровского края разновидности тестового контроля
разновидности тестового контроля Внешнее строение листа. Урок биологии в 6 классе
Внешнее строение листа. Урок биологии в 6 классе Разнообразие растительного мира. Семейство Норичниковые
Разнообразие растительного мира. Семейство Норичниковые