- Кальциевые каналы

Содержание
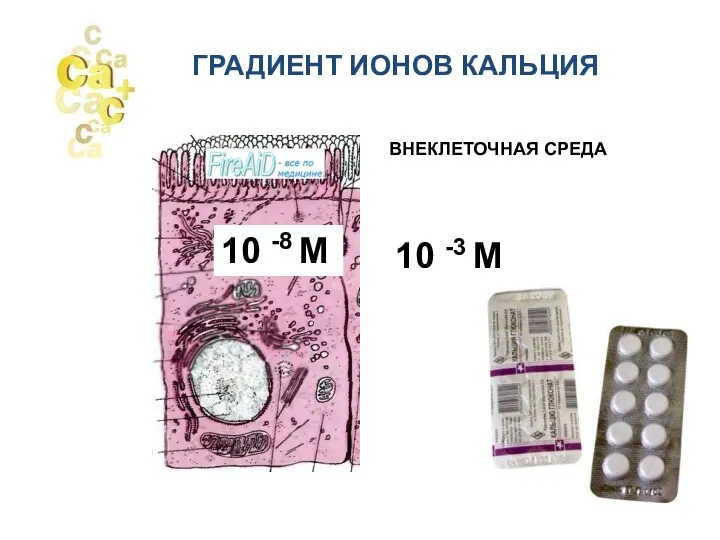
- 2. ГРАДИЕНТ ИОНОВ КАЛЬЦИЯ 10 -8 М 10 -3 М ВНЕКЛЕТОЧНАЯ СРЕДА
- 3. РОЛЬ ИОНОВ КАЛЬЦИЯ 1. СОПРЯЖЕНИЕ ПРОЦЕССОВ ВОЗБУЖДЕНИЯ - СОКРАЩЕНИЯ
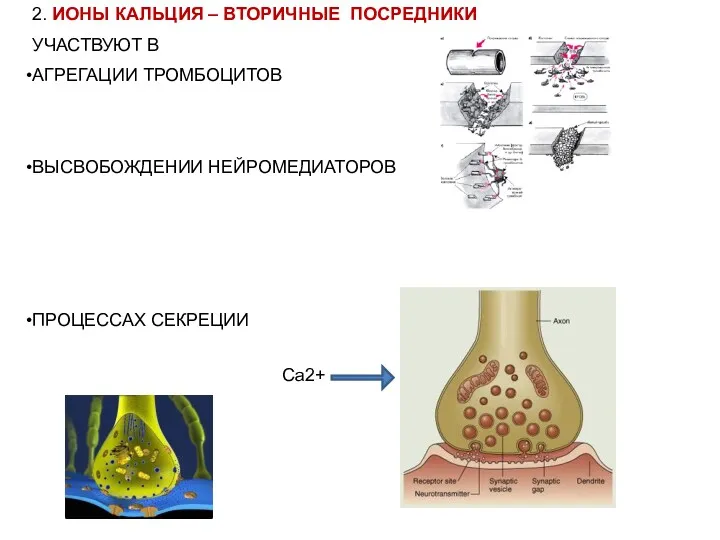
- 4. 2. ИОНЫ КАЛЬЦИЯ – ВТОРИЧНЫЕ ПОСРЕДНИКИ УЧАСТВУЮТ В АГРЕГАЦИИ ТРОМБОЦИТОВ ВЫСВОБОЖДЕНИИ НЕЙРОМЕДИАТОРОВ ПРОЦЕССАХ СЕКРЕЦИИ Са2+
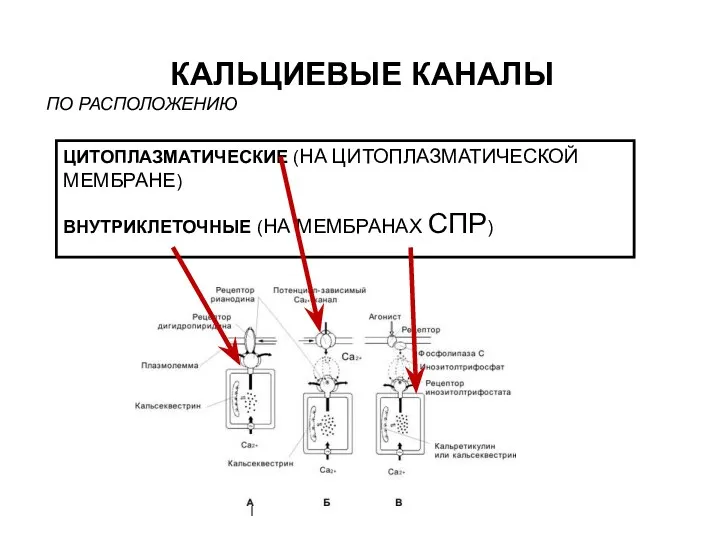
- 5. КАЛЬЦИЕВЫЕ КАНАЛЫ ЦИТОПЛАЗМАТИЧЕСКИЕ (НА ЦИТОПЛАЗМАТИЧЕСКОЙ МЕМБРАНЕ) ВНУТРИКЛЕТОЧНЫЕ (НА МЕМБРАНАХ СПР) ПО РАСПОЛОЖЕНИЮ
- 6. ПО СПОСОБУ УПРАВЛЕНИЯ ПОТЕНЦИАЛОЗАВИСИМЫЕ РЕЦЕПТОРУПРАВЛЯЕМЫЕ КАЛЬЦИЕВЫЕ КАНАЛЫ
- 7. ПОТЕНЦИАЛОЗАВИСИМЫЕ КАЛЬЦИЕВЫЕ КАНАЛЫ
- 8. Классификация каналов по потенциал-управляемости Высокопороговые каналы (HVA) Порог активации значительно выше ПП КАНАЛЫ L-ТИПА КАНАЛЫ –NТИПА
- 9. КАНАЛЫ L-ТИПА (МЕДЛЕННЫЕ КАНАЛЫ) ВЫСОКОВОЛЬТАЖНЫЕ: АКТИВИРУЮТСЯ ПРИ ОТНОСИТЕЛЬНО ВЫСОКИХ ЗНАЧЕНИЯХ МП (свыше 10 мВ) ВЫСОКАЯ ПРОВОДИМОСТЬ
- 10. ВЗАИМОДЕЙСТВИЕ СУБЪЕДИНИЦ КАНАЛА СТРУКТУРА Са2+-канала сходна со структурой Na-канала, но представлена 5-ю субъединицами: α1, α2, β,
- 11. Мембранная локализация субъединиц Са2 +канала и формирование ионопроводящей поры α1субъединицей При этом сегменты S5 и S6
- 12. ДВА МЕХАНИЗМА ИНАКТИВАЦИИ Са2+- каналов L-типа Потенциалозависимая инактивируются деполяризацией Кальцийзависимая (токозависимая) инактивируются ионами Са2+, вошедшими в
- 13. ИЗУЧЕНИЕ ПОТЕНЦИАЛОЗАВИСИМЫХ КАЛЬЦИЕВЫХ КАНАЛОВ – РАБОТЫ ХАГИВАРА С СОАВТ. НА МЫШЦАХ МОРСКОГО ЖЕЛУДЯ (усоногие ракообразные)
- 14. ПРОНИЦАЕМОСТЬ Са 2+-КАНАЛА ДЛЯ ДВУХВАЛЕНТНЫХ КАТИОНОВ Ba2+ > Sr2+ > Ca2+ > Co2+ > Ni2+ >
- 15. МОДЕЛЬ: ИОН КАЛЬЦИЯ СВЯЗЫВАЕТСЯ С ОПРЕДЕЛЕННОЙ СТРУКТУРОЙ КАНАЛА X + Caex X Ca X + Cain
- 16. РОЛЬ КАЛЬЦИЕВЫХ КАНАЛОВ L-ТИПА ОСНОВНОЙ ПУТЬ ДЛЯ ВХОДА ИОНОВ КАЛЬЦИЯ В КЛЕТКУ (ВАЖНО ДЛЯ СОКРАЩЕНИЯ МЫШЦ,
- 18. КАНАЛЫ -NТИПА АКТИВИРУЮТСЯ ПРИ ВЫСОКИХ ЗНАЧЕНИЯХ МП (более 20 мВ) ОТНОСИТЕЛЬНО БЫСТРО ИНАКТИВИРУЮТСЯ (t~50-80 мс) СРЕДНЯЯ
- 19. РОЛЬ КАЛЬЦИЕВЫХ КАНАЛОВ N-ТИПА Регулируют высвобождение медиаторов
- 20. КАНАЛЫ P И QТИПОВ КАНАЛ Р ТИПА ПОРОГ АКТИВАЦИИ СОСТАВЛЯЕТ –50МВ, КИНЕТИКА ИНАКТИВАЦИИ ОЧЕНЬ МЕДЛЕННАЯ (t
- 21. КАНАЛЫ T-ТИПА (БЫСТРЫЕ КАНАЛЫ) НИЗКОВОЛЬТАЖНЫЕ: АКТИВИРУЮТСЯ ПРИ НИЗКИХ ЗНАЧЕНИЯХ МП (МП более положителен, чем -70 мВ)
- 22. РОЛЬ КАЛЬЦИЕВЫХ КАНАЛОВ Т-ТИПА В ЭМБРИОНАЛЬНЫХ КЛЕТКАХ ЗАПУСКАЮТ ПРОЦЕССЫ ДИФФЕРЕНЦИРОВКИ ОБЕСПЕЧИВАЮТ ЗАКОНОМЕРНЫЕ ПЕРИОДИЧЕСКИЕ КОЛЕБАНИЯ МП (НАПРИМЕР,
- 23. Фармакологическая классификация DHP-чувствительные кальциевые каналы Lтипа DHP-нечувствительные кальциевые каналы Nтипа. По чувствительности к дигидропиридинам (DHP) HVA
- 24. ВНУТРИКЛЕТОЧНЫЕ КАЛЬЦИЕВЫЕ КАНАЛЫ
- 26. ИЗОФОРМЫ РИАНОДИНОВЫХ РЕЦЕПТОРОВ обнаружены в мозге
- 29. IP3-РЕЦЕПТОРЫ: Активируются при увеличении внутриклеточной концентрации IP3 вызывают высвобождение внутриклеточных запасов Ca2+ после стимуляции рецепторов на
- 31. АКТИВНЫЙ ТРАНСПОРТ
- 32. ОБЩИЕ СВОЙСТВА: ТРАНСПОРТ СОПРЯЖЕН С ГИДРОЛИЗОМ АТФ ПЕРЕНОС ИОНОВ ОСУЩЕСТВЛЯЮТ ТРАНСПОРТНЫЕ АТФАЗЫ ТРАНСПОРТ ОСУЩЕСТВЛЯЕТСЯ ПРОТИВ ГРАДИЕНТА
- 33. ТИПЫ АТФАЗ АТФаза V-типа АТФаза F-типа АТФаза Р-типа
- 34. АТФазы V типа ЛОКАЛИЗАЦИЯ: МЕМБРАНЫ ВАКОЛЕЙ ДРОЖЖЕЙ ТОНОПЛАСТЫ РАСТЕНИЙ ЛИЗОСОМЫ ЭНДОСОМЫ СЕКРЕТОРНЫЕ ГРАНУЛЫ
- 35. V0 – ГИДРОФОБНАЯ ЧАСТЬ (УЧАСТВУЕТ В ТРАНСЛОКАЦИИ ПРОТОНОВ) V1 – ВОДОРАСТВОРИМАЯ ЧАСТЬ ОБЛАДАЕТ КАТАЛИТИЧЕСКОЙ АКТИВНОСТЬЮ, КАТАЛИЗИРУЕТ
- 36. ФУНКЦИИ ПЕРЕНОСЯТ ПРОТОНЫ УЧАСТВУЮТ В ТРАНСПОРТЕ АНИОНОВ, АМИНОКИСЛОТ И РЕПАРАЦИИ МЕМБРАН ПРИ ЭНДО- И ЭКЗОЦИТОЗЕ ИНГИБИТОРЫ:
- 37. ТРАНСПОРТ ПРОТОНОВ В ТОНОПЛАСТЕ КЛЕТОК РАСТЕНИЙ ПРОТОНЫ ПОСТУПАЮТ ВНУТРЬ ВАКУОЛИ И СОЗДАЮТ ЭЛЕКТРИЧЕСКИЙ (от +20 до
- 38. АТФаза F ТИПА ЛОКАЛИЗАЦИЯ: МЕМБРАНЫ БАКТЕРИЙ, ХЛОРОПЛАСТОВ И МИТОХОНДРИЙ СТРОЕНИЕ: F0 – ГИДРОФОБНАЯ ЧАСТЬ (ТРАНСЛОКАЦИЯ ПРОТОНОВ)
- 39. АТФазы Р ТИПА ОБЩЕЕ СВОЙСТВО: ОБРАЗОВАНИЕ ФОСФОРИЛИРОВАННОГО ПРОДУКТА, КОТОРЫЙ УЧАСТВУЕТ В РЕАКЦИОННОЙ ЦИКЛЕ ПРИМЕРЫ: Na/K-АТФаза, Ca-АТФаза,
- 40. Na/K АТФаза ЛОКАЛИЗОВАНА НА ЦИТОПЛАЗМАТИЧЕСКОЙ МЕМБРАНЕ СОЗДАЕТ ГРАДИЕНТ ИОНОВ НАТРИЯ И КАЛИЯ
- 41. СТРУКТУРА Na+/K+ АТФазы: состоит из 2 полипептидных цепей α и β 1 – участок связывания ионов
- 42. УАБАИН – ингибитор Na,K-АТФазы Строфант
- 43. РЕАКЦИОННЫЙ ЦИКЛ Na/K АТФазы
- 44. 2 - изменение конформации транслоказы, вызванное присоединением 3Na+, приводит к активации каталитической субъединицы и увеличению сродства
- 45. 3 - аутофосфорилирование изменяет заряд и конформацию транслоказы, она закрывается с внутренней стороны мембраны и открывается
- 46. 5 - дефосфорилирование изменяет заряд и конформацию транслоказы, она закрывается с наружной стороны мембраны и открывается
- 47. РЕГУЛЯЦИЯ АКТИВНОСТИ Na/K АТФазы СООТНОШЕНИЕ Na/K И СОДЕРЖАНИЕ АТФ (факторы краткосрочной регуляции) ФОСФОРИЛИРОВАНИЕ ПРОТЕИНКИНАЗАМИ, ЧТО ПРИВОДИТ
- 48. Са2+ АТФаза ЛОКАЛИЗАЦИЯ: САРКО- (ЭНДО)-ПЛАЗМАТИЧЕСКИЙ РЕТИКУЛУМ ЦИТОПЛАЗМАТИЧЕСКАЯ МЕМБРАНА
- 49. ВСЕ Са2+ АТФазы – МОНОМЕРНЫЕ БЕЛКИ, Т.Е. СОСТОЯТ ИЗ ОДНОЙ ПОЛИПЕПТИДНОЙ ЦЕПИ Са2+ АТФаза СПР И
- 50. ФЛБ – фосфоламбан (у Са АТФазы саркоплазматического ретикулума), СПР СПР
- 51. В фосфорилированном состоянии фосфоламбан стимулирует кальциевый насос ретикулума миоцитов, ускоряя восстановление кальциевых градиентов.
- 52. КМ – кальмодулин (у Са АТФазы плазматической мембраны ) (цитозоль) (внеклеточная среда)
- 53. ЦИКЛ РАБОТЫ Са2+АТФазы СТАДИИ ГИДРОЛИЗА АТФ ЧЕРЕДУЮТСЯ СО СТАДИЯМИ ПЕРЕНОСА ИОНОВ КАЛЬЦИЯ
- 54. ЛОКАЛИЗАЦИЯ: ПЛАЗМАТИЧЕСКИЕ МЕМБРАНЫ РАСТИТЕЛЬНЫХ КЛЕТОК Н+АТФаза
- 55. Н+-АТФаза – это интегральный белок, полипептидная цепь которого десять раз пересекает поверхностную (плазматическую) мембрану. Полагают, что
- 57. Роль Н+-АТФазы: поддерживает рН цитоплазмы близкий к нейтральному создает на мембране разность потенциалов (∆ψ), во многом
- 58. Механизм возникновения ПД в проводящих пучках высших растений имеет большое сходство с таковым в нервах животных.
- 59. Благодаря вторичному активному транспорту клетка активно поглощает (или удаляет) многие вещества (ионы, углеводы, аминокислоты и др.).
- 60. ТРАНСПОРТ ВЕЩЕСТВ ЧЕРЕЗ МНОГОМЕМБРАННЫЕ СИСТЕМЫ ВТОРИЧНЫЙ АКТИВНЫЙ ТРАНСПОРТ
- 61. МНОГОМЕМБРАННЫЕ СИСТЕМЫ гликокаликс + коллаген микроворсинки базолатеральная поверхность
- 62. БАЗАЛЬНАЯ МЕМБРАНА – дополнительная мембрана за пределами плазмалеммы эпителиоцита ЕЕ СОСТАВ: гликопротеидный матрикс (гликокаликс) + коллагеновый
- 63. Базолатеральная мембрана несет межклеточные контакты разного типа
- 64. ЭКСПЕРИМЕНТЫ УССИНГА Установка Уссинга Объект исследований
- 65. ЭКСПЕРИМЕНТ УССИНГА: ИЗУЧЕНИЕ АСИММЕТРИЧНЫХ СВОЙСТВ ЭПИТЕЛИЯ 1 – кожа лягушки; 2 – вольтметр; 3 и 4
- 66. МОДЕЛЬ УССИНГА Мукозная (наружная)поверхность кожи Серозная (внутренняя)поверхность кожи
- 67. ВТОРИЧНЫЙ АКТИВНЫЙ ТРАНСПОРТ
- 68. ВТОРИЧНЫЙ АКТИВНЫЙ ТРАНСПОРТ ТРАНСПОРТ САХАРОВ И АМИНОКИСЛОТ ЗА СЧЕТ ЭНЕРГИИ ГРАДИЕНТА Na+ , КОТОРЫЙ СОЗДАЕТСЯ БЛАГОДАРЯ
- 69. Уравнение для транспорта сахаров Jmax = 12 мкмоль / м2 с – одинакова для всех моносахаридов
- 70. Графики Лайнуивера – Берка для транспорта 6-дезокси-D-глюкозы через эпителий кишки, показывающие зависимость транспорта сахара от концентрации
- 71. ТРАНСПОРТ ГЛЮКОЗЫ, СОПРЯЖЕННЫЙ С ИОНАМИ НАТРИЯ
- 72. ТРАНСПОРТ АМИНОКИСЛОТ, СОПРЯЖЕННЫЙ С ИОНАМИ НАТРИЯ
- 75. Скачать презентацию
ГРАДИЕНТ ИОНОВ КАЛЬЦИЯ
10 -8 М
10 -3 М
ВНЕКЛЕТОЧНАЯ СРЕДА
ГРАДИЕНТ ИОНОВ КАЛЬЦИЯ
10 -8 М
10 -3 М
ВНЕКЛЕТОЧНАЯ СРЕДА
РОЛЬ ИОНОВ КАЛЬЦИЯ
1. СОПРЯЖЕНИЕ ПРОЦЕССОВ ВОЗБУЖДЕНИЯ - СОКРАЩЕНИЯ
РОЛЬ ИОНОВ КАЛЬЦИЯ
1. СОПРЯЖЕНИЕ ПРОЦЕССОВ ВОЗБУЖДЕНИЯ - СОКРАЩЕНИЯ
2. ИОНЫ КАЛЬЦИЯ – ВТОРИЧНЫЕ ПОСРЕДНИКИ
УЧАСТВУЮТ В
АГРЕГАЦИИ ТРОМБОЦИТОВ
ВЫСВОБОЖДЕНИИ НЕЙРОМЕДИАТОРОВ
ПРОЦЕССАХ
2. ИОНЫ КАЛЬЦИЯ – ВТОРИЧНЫЕ ПОСРЕДНИКИ
УЧАСТВУЮТ В
АГРЕГАЦИИ ТРОМБОЦИТОВ
ВЫСВОБОЖДЕНИИ НЕЙРОМЕДИАТОРОВ
ПРОЦЕССАХ
КАЛЬЦИЕВЫЕ КАНАЛЫ
ЦИТОПЛАЗМАТИЧЕСКИЕ (НА ЦИТОПЛАЗМАТИЧЕСКОЙ МЕМБРАНЕ)
ВНУТРИКЛЕТОЧНЫЕ (НА МЕМБРАНАХ СПР)
ПО РАСПОЛОЖЕНИЮ
КАЛЬЦИЕВЫЕ КАНАЛЫ
ЦИТОПЛАЗМАТИЧЕСКИЕ (НА ЦИТОПЛАЗМАТИЧЕСКОЙ МЕМБРАНЕ)
ВНУТРИКЛЕТОЧНЫЕ (НА МЕМБРАНАХ СПР)
ПО РАСПОЛОЖЕНИЮ

ПО СПОСОБУ УПРАВЛЕНИЯ
ПОТЕНЦИАЛОЗАВИСИМЫЕ
РЕЦЕПТОРУПРАВЛЯЕМЫЕ
КАЛЬЦИЕВЫЕ КАНАЛЫ
ПО СПОСОБУ УПРАВЛЕНИЯ
ПОТЕНЦИАЛОЗАВИСИМЫЕ
РЕЦЕПТОРУПРАВЛЯЕМЫЕ
КАЛЬЦИЕВЫЕ КАНАЛЫ

ПОТЕНЦИАЛОЗАВИСИМЫЕ КАЛЬЦИЕВЫЕ КАНАЛЫ
ПОТЕНЦИАЛОЗАВИСИМЫЕ КАЛЬЦИЕВЫЕ КАНАЛЫ
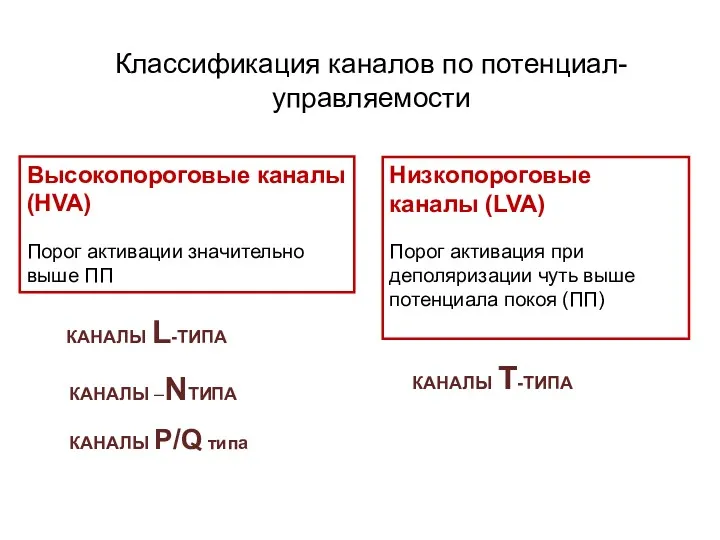
Классификация каналов по потенциал-управляемости
Высокопороговые каналы (HVA)
Порог активации значительно
выше ПП
КАНАЛЫ L-ТИПА
Классификация каналов по потенциал-управляемости
Высокопороговые каналы (HVA)
Порог активации значительно
выше ПП
КАНАЛЫ L-ТИПА

КАНАЛЫ L-ТИПА (МЕДЛЕННЫЕ КАНАЛЫ) ВЫСОКОВОЛЬТАЖНЫЕ:
АКТИВИРУЮТСЯ ПРИ ОТНОСИТЕЛЬНО ВЫСОКИХ ЗНАЧЕНИЯХ МП
КАНАЛЫ L-ТИПА (МЕДЛЕННЫЕ КАНАЛЫ) ВЫСОКОВОЛЬТАЖНЫЕ:
АКТИВИРУЮТСЯ ПРИ ОТНОСИТЕЛЬНО ВЫСОКИХ ЗНАЧЕНИЯХ МП
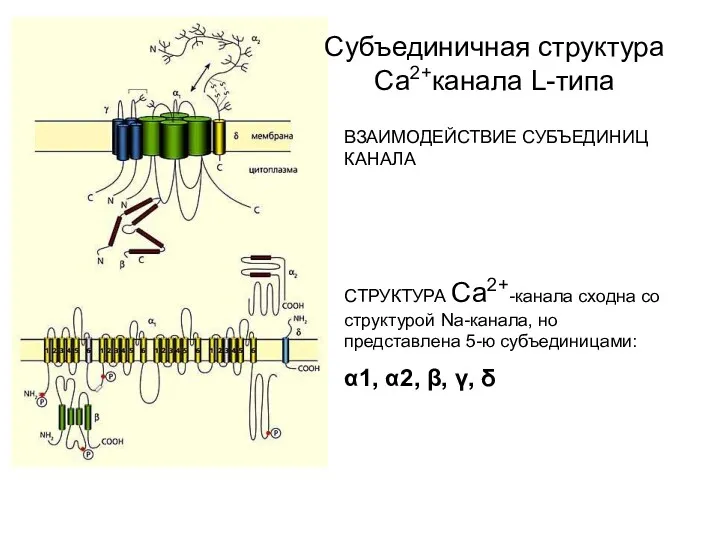
ВЗАИМОДЕЙСТВИЕ СУБЪЕДИНИЦ КАНАЛА
СТРУКТУРА Са2+-канала сходна со структурой Na-канала, но представлена 5-ю
ВЗАИМОДЕЙСТВИЕ СУБЪЕДИНИЦ КАНАЛА
СТРУКТУРА Са2+-канала сходна со структурой Na-канала, но представлена 5-ю
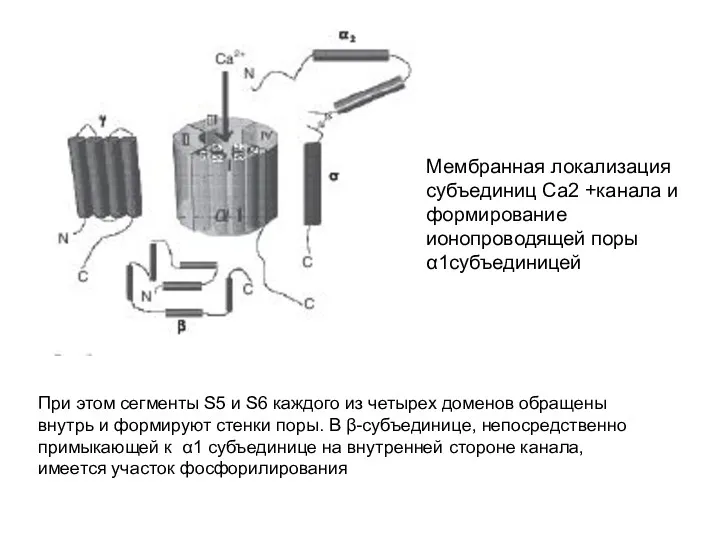
Мембранная локализация субъединиц Са2 +канала и формирование ионопроводящей поры α1субъединицей
При этом
Мембранная локализация субъединиц Са2 +канала и формирование ионопроводящей поры α1субъединицей
При этом
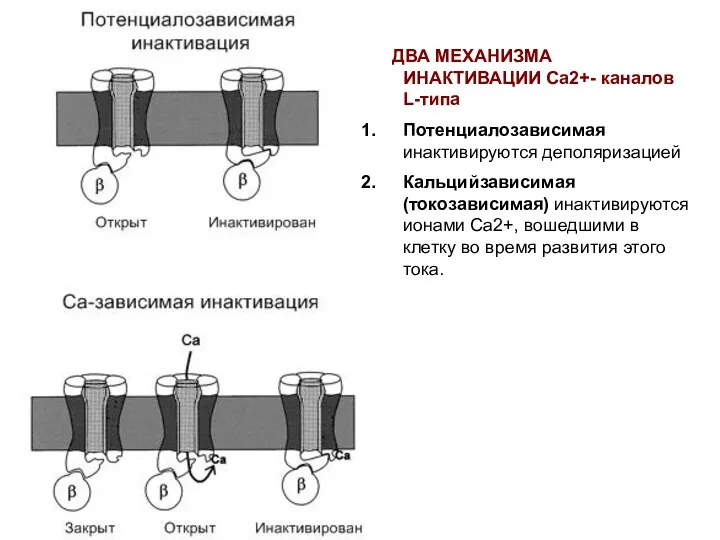
ДВА МЕХАНИЗМА ИНАКТИВАЦИИ Са2+- каналов L-типа
Потенциалозависимая инактивируются деполяризацией
Кальцийзависимая (токозависимая)
ДВА МЕХАНИЗМА ИНАКТИВАЦИИ Са2+- каналов L-типа
Потенциалозависимая инактивируются деполяризацией
Кальцийзависимая (токозависимая)

ИЗУЧЕНИЕ ПОТЕНЦИАЛОЗАВИСИМЫХ КАЛЬЦИЕВЫХ КАНАЛОВ – РАБОТЫ ХАГИВАРА С СОАВТ. НА МЫШЦАХ
ИЗУЧЕНИЕ ПОТЕНЦИАЛОЗАВИСИМЫХ КАЛЬЦИЕВЫХ КАНАЛОВ – РАБОТЫ ХАГИВАРА С СОАВТ. НА МЫШЦАХ
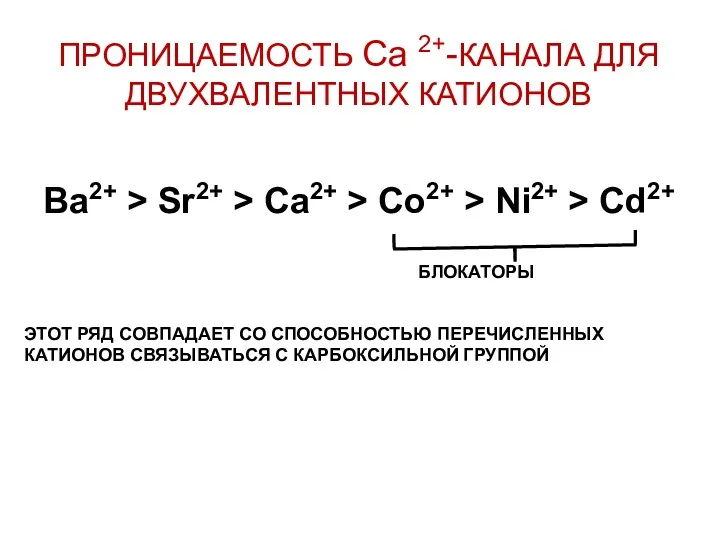
ПРОНИЦАЕМОСТЬ Са 2+-КАНАЛА ДЛЯ ДВУХВАЛЕНТНЫХ КАТИОНОВ
Ba2+ > Sr2+ > Ca2+ >
ПРОНИЦАЕМОСТЬ Са 2+-КАНАЛА ДЛЯ ДВУХВАЛЕНТНЫХ КАТИОНОВ
Ba2+ > Sr2+ > Ca2+ >

МОДЕЛЬ: ИОН КАЛЬЦИЯ СВЯЗЫВАЕТСЯ С ОПРЕДЕЛЕННОЙ СТРУКТУРОЙ КАНАЛА
X + Caex X
МОДЕЛЬ: ИОН КАЛЬЦИЯ СВЯЗЫВАЕТСЯ С ОПРЕДЕЛЕННОЙ СТРУКТУРОЙ КАНАЛА
X + Caex X

РОЛЬ КАЛЬЦИЕВЫХ КАНАЛОВ L-ТИПА
ОСНОВНОЙ ПУТЬ ДЛЯ ВХОДА ИОНОВ КАЛЬЦИЯ В
РОЛЬ КАЛЬЦИЕВЫХ КАНАЛОВ L-ТИПА
ОСНОВНОЙ ПУТЬ ДЛЯ ВХОДА ИОНОВ КАЛЬЦИЯ В


КАНАЛЫ -NТИПА
АКТИВИРУЮТСЯ ПРИ ВЫСОКИХ ЗНАЧЕНИЯХ МП (более 20 мВ)
ОТНОСИТЕЛЬНО
КАНАЛЫ -NТИПА
АКТИВИРУЮТСЯ ПРИ ВЫСОКИХ ЗНАЧЕНИЯХ МП (более 20 мВ)
ОТНОСИТЕЛЬНО

РОЛЬ КАЛЬЦИЕВЫХ КАНАЛОВ N-ТИПА
Регулируют высвобождение медиаторов
РОЛЬ КАЛЬЦИЕВЫХ КАНАЛОВ N-ТИПА
Регулируют высвобождение медиаторов

КАНАЛЫ P И QТИПОВ
КАНАЛ Р ТИПА
ПОРОГ АКТИВАЦИИ СОСТАВЛЯЕТ –50МВ,
КИНЕТИКА
КАНАЛЫ P И QТИПОВ
КАНАЛ Р ТИПА
ПОРОГ АКТИВАЦИИ СОСТАВЛЯЕТ –50МВ,
КИНЕТИКА

КАНАЛЫ T-ТИПА (БЫСТРЫЕ КАНАЛЫ) НИЗКОВОЛЬТАЖНЫЕ:
АКТИВИРУЮТСЯ ПРИ НИЗКИХ ЗНАЧЕНИЯХ МП (МП
КАНАЛЫ T-ТИПА (БЫСТРЫЕ КАНАЛЫ) НИЗКОВОЛЬТАЖНЫЕ:
АКТИВИРУЮТСЯ ПРИ НИЗКИХ ЗНАЧЕНИЯХ МП (МП

РОЛЬ КАЛЬЦИЕВЫХ КАНАЛОВ Т-ТИПА
В ЭМБРИОНАЛЬНЫХ КЛЕТКАХ ЗАПУСКАЮТ ПРОЦЕССЫ ДИФФЕРЕНЦИРОВКИ
ОБЕСПЕЧИВАЮТ ЗАКОНОМЕРНЫЕ ПЕРИОДИЧЕСКИЕ
В ЭМБРИОНАЛЬНЫХ КЛЕТКАХ ЗАПУСКАЮТ ПРОЦЕССЫ ДИФФЕРЕНЦИРОВКИ
ОБЕСПЕЧИВАЮТ ЗАКОНОМЕРНЫЕ ПЕРИОДИЧЕСКИЕ
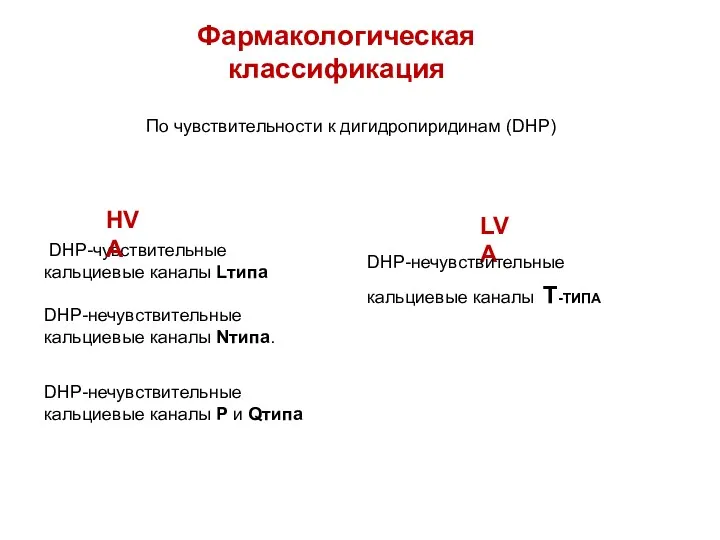
Фармакологическая классификация
DHP-чувствительные
кальциевые каналы Lтипа
DHP-нечувствительные
кальциевые каналы Nтипа.
По чувствительности к
Фармакологическая классификация
DHP-чувствительные
кальциевые каналы Lтипа
DHP-нечувствительные
кальциевые каналы Nтипа.
По чувствительности к

ВНУТРИКЛЕТОЧНЫЕ КАЛЬЦИЕВЫЕ КАНАЛЫ
ВНУТРИКЛЕТОЧНЫЕ КАЛЬЦИЕВЫЕ КАНАЛЫ

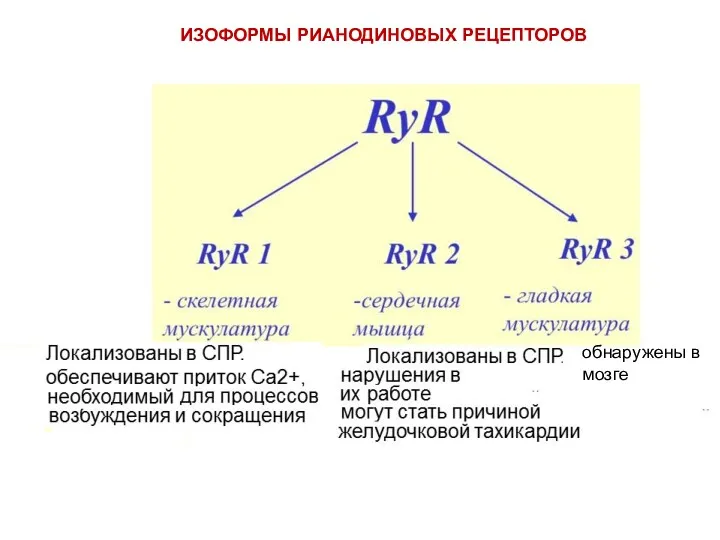
ИЗОФОРМЫ РИАНОДИНОВЫХ РЕЦЕПТОРОВ
обнаружены в мозге
ИЗОФОРМЫ РИАНОДИНОВЫХ РЕЦЕПТОРОВ
обнаружены в мозге
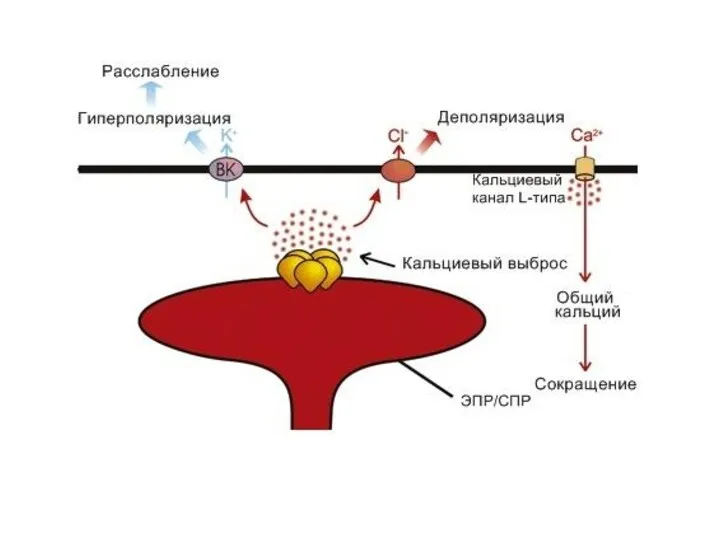
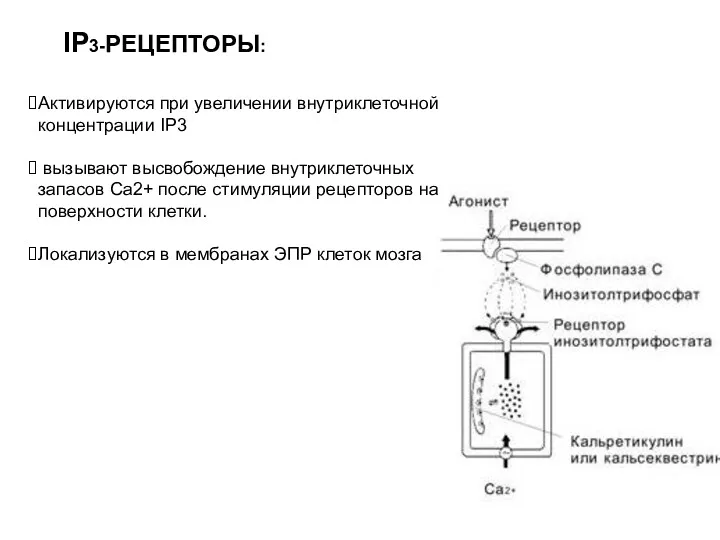
IP3-РЕЦЕПТОРЫ:
Активируются при увеличении внутриклеточной концентрации IP3
вызывают высвобождение внутриклеточных
IP3-РЕЦЕПТОРЫ:
Активируются при увеличении внутриклеточной концентрации IP3
вызывают высвобождение внутриклеточных

АКТИВНЫЙ ТРАНСПОРТ
АКТИВНЫЙ ТРАНСПОРТ

ОБЩИЕ СВОЙСТВА:
ТРАНСПОРТ СОПРЯЖЕН С ГИДРОЛИЗОМ АТФ
ПЕРЕНОС ИОНОВ ОСУЩЕСТВЛЯЮТ ТРАНСПОРТНЫЕ АТФАЗЫ
ТРАНСПОРТ
ОБЩИЕ СВОЙСТВА:
ТРАНСПОРТ СОПРЯЖЕН С ГИДРОЛИЗОМ АТФ
ПЕРЕНОС ИОНОВ ОСУЩЕСТВЛЯЮТ ТРАНСПОРТНЫЕ АТФАЗЫ
ТРАНСПОРТ
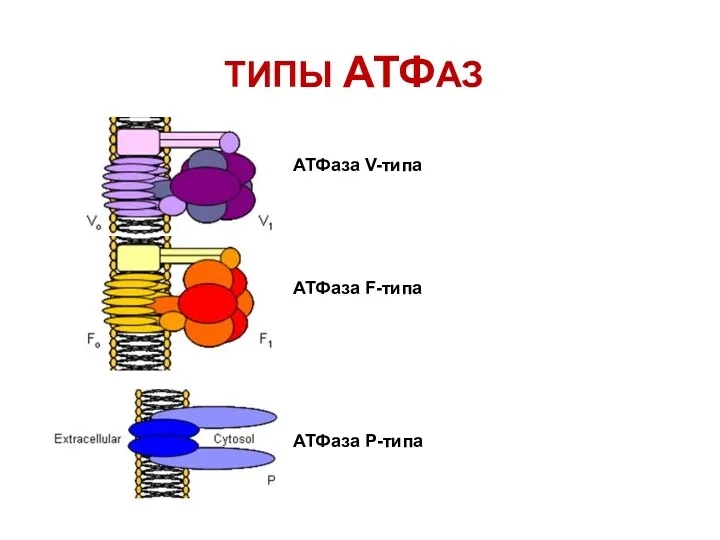
ТИПЫ АТФАЗ
АТФаза V-типа
АТФаза F-типа
АТФаза Р-типа
ТИПЫ АТФАЗ
АТФаза V-типа
АТФаза F-типа
АТФаза Р-типа
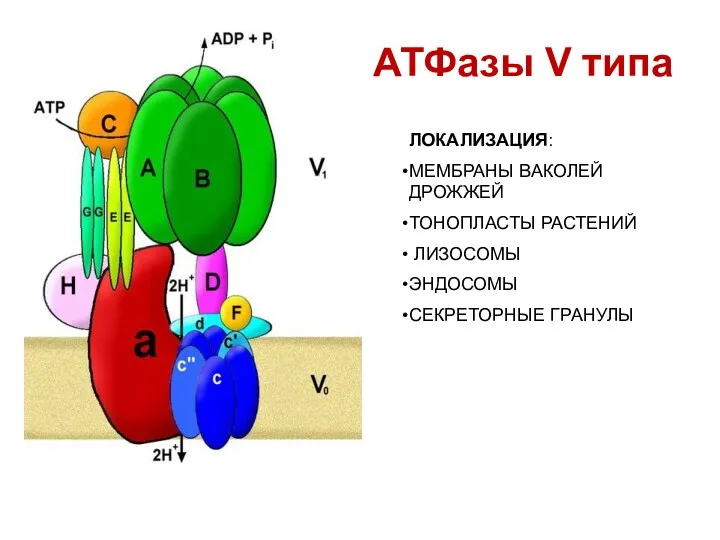
АТФазы V типа
ЛОКАЛИЗАЦИЯ:
МЕМБРАНЫ ВАКОЛЕЙ ДРОЖЖЕЙ
ТОНОПЛАСТЫ РАСТЕНИЙ
ЛИЗОСОМЫ
ЭНДОСОМЫ
СЕКРЕТОРНЫЕ ГРАНУЛЫ
АТФазы V типа
ЛОКАЛИЗАЦИЯ:
МЕМБРАНЫ ВАКОЛЕЙ ДРОЖЖЕЙ
ТОНОПЛАСТЫ РАСТЕНИЙ
ЛИЗОСОМЫ
ЭНДОСОМЫ
СЕКРЕТОРНЫЕ ГРАНУЛЫ
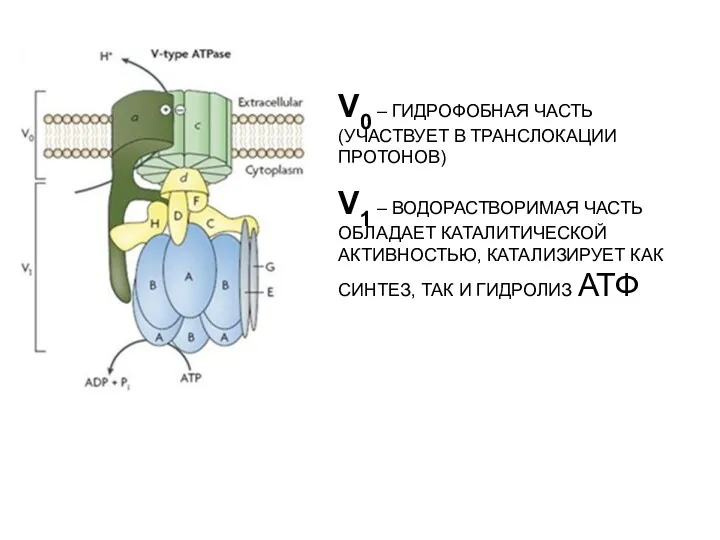
V0 – ГИДРОФОБНАЯ ЧАСТЬ (УЧАСТВУЕТ В ТРАНСЛОКАЦИИ ПРОТОНОВ)
V1 – ВОДОРАСТВОРИМАЯ ЧАСТЬ
V0 – ГИДРОФОБНАЯ ЧАСТЬ (УЧАСТВУЕТ В ТРАНСЛОКАЦИИ ПРОТОНОВ)
V1 – ВОДОРАСТВОРИМАЯ ЧАСТЬ
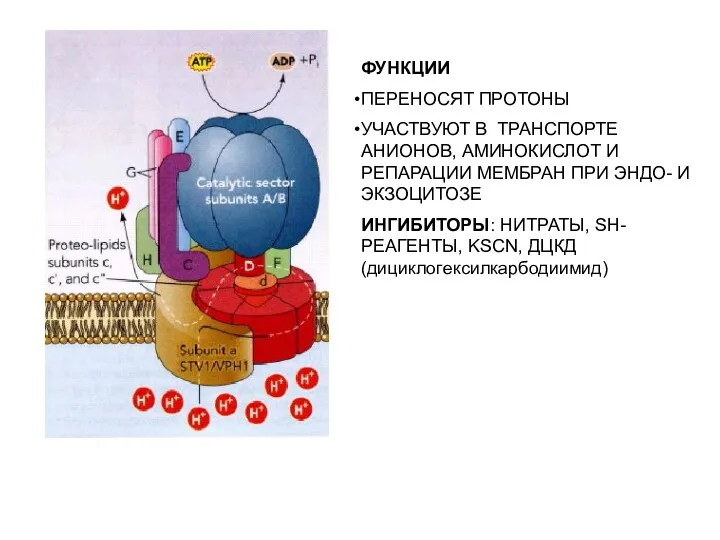
ФУНКЦИИ
ПЕРЕНОСЯТ ПРОТОНЫ
УЧАСТВУЮТ В ТРАНСПОРТЕ АНИОНОВ, АМИНОКИСЛОТ И РЕПАРАЦИИ МЕМБРАН ПРИ ЭНДО-
ФУНКЦИИ
ПЕРЕНОСЯТ ПРОТОНЫ
УЧАСТВУЮТ В ТРАНСПОРТЕ АНИОНОВ, АМИНОКИСЛОТ И РЕПАРАЦИИ МЕМБРАН ПРИ ЭНДО-
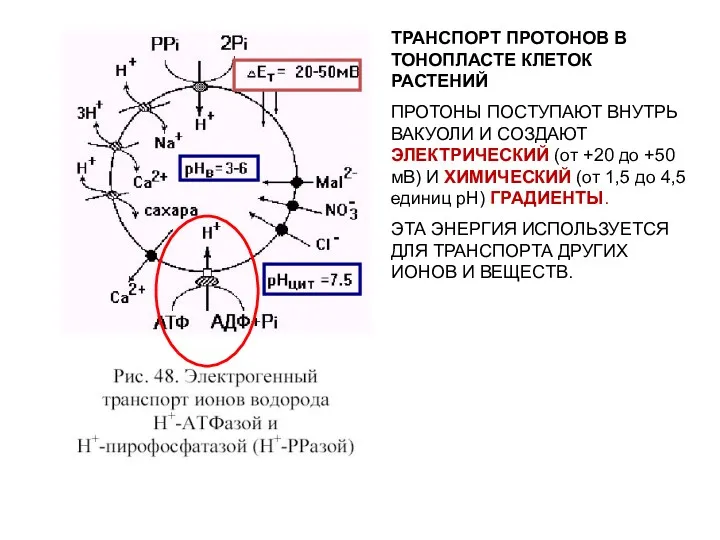
ТРАНСПОРТ ПРОТОНОВ В ТОНОПЛАСТЕ КЛЕТОК РАСТЕНИЙ
ПРОТОНЫ ПОСТУПАЮТ ВНУТРЬ ВАКУОЛИ И СОЗДАЮТ
ТРАНСПОРТ ПРОТОНОВ В ТОНОПЛАСТЕ КЛЕТОК РАСТЕНИЙ
ПРОТОНЫ ПОСТУПАЮТ ВНУТРЬ ВАКУОЛИ И СОЗДАЮТ
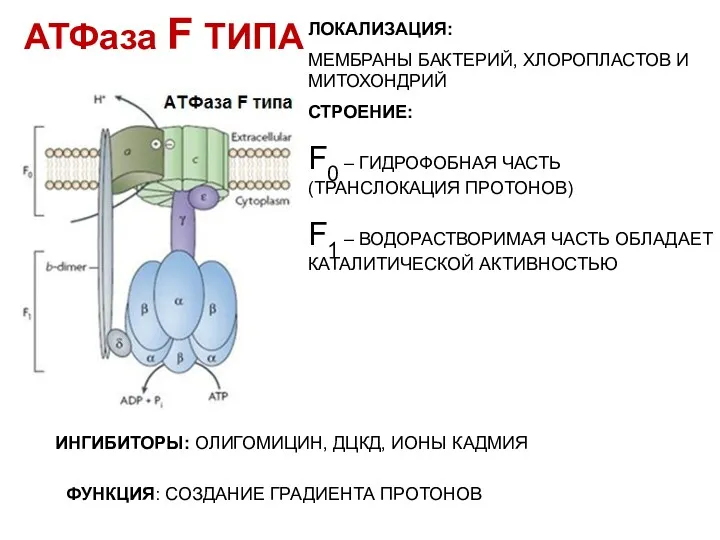
АТФаза F ТИПА
ЛОКАЛИЗАЦИЯ:
МЕМБРАНЫ БАКТЕРИЙ, ХЛОРОПЛАСТОВ И МИТОХОНДРИЙ
СТРОЕНИЕ:
F0 – ГИДРОФОБНАЯ ЧАСТЬ (ТРАНСЛОКАЦИЯ
АТФаза F ТИПА
ЛОКАЛИЗАЦИЯ:
МЕМБРАНЫ БАКТЕРИЙ, ХЛОРОПЛАСТОВ И МИТОХОНДРИЙ
СТРОЕНИЕ:
F0 – ГИДРОФОБНАЯ ЧАСТЬ (ТРАНСЛОКАЦИЯ

АТФазы Р ТИПА
ОБЩЕЕ СВОЙСТВО: ОБРАЗОВАНИЕ ФОСФОРИЛИРОВАННОГО ПРОДУКТА, КОТОРЫЙ УЧАСТВУЕТ В РЕАКЦИОННОЙ
АТФазы Р ТИПА
ОБЩЕЕ СВОЙСТВО: ОБРАЗОВАНИЕ ФОСФОРИЛИРОВАННОГО ПРОДУКТА, КОТОРЫЙ УЧАСТВУЕТ В РЕАКЦИОННОЙ
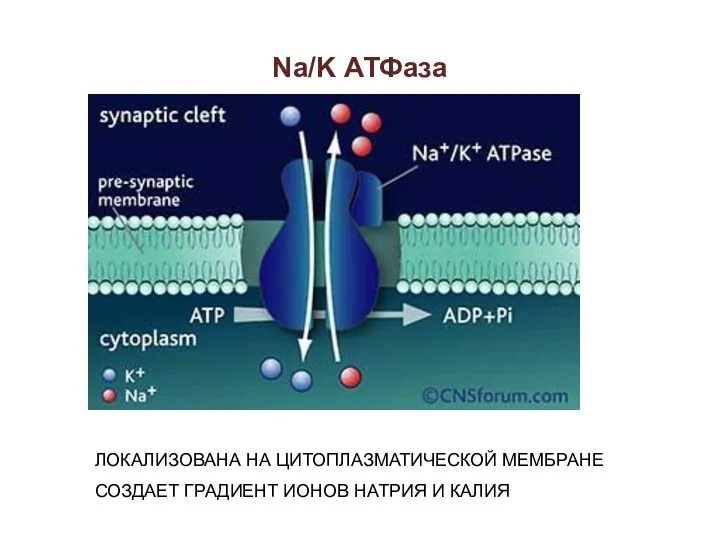
Na/K АТФаза
ЛОКАЛИЗОВАНА НА ЦИТОПЛАЗМАТИЧЕСКОЙ МЕМБРАНЕ
СОЗДАЕТ ГРАДИЕНТ ИОНОВ НАТРИЯ И КАЛИЯ
Na/K АТФаза
ЛОКАЛИЗОВАНА НА ЦИТОПЛАЗМАТИЧЕСКОЙ МЕМБРАНЕ
СОЗДАЕТ ГРАДИЕНТ ИОНОВ НАТРИЯ И КАЛИЯ
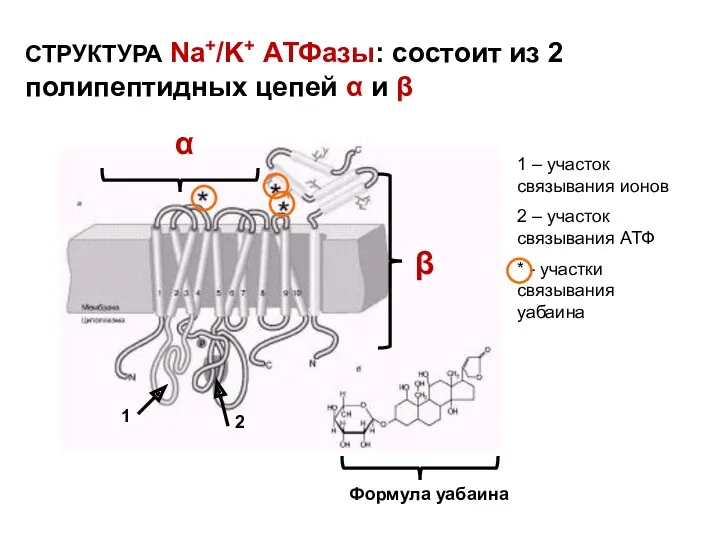
СТРУКТУРА Na+/K+ АТФазы: состоит из 2 полипептидных цепей α и β
1
СТРУКТУРА Na+/K+ АТФазы: состоит из 2 полипептидных цепей α и β
1
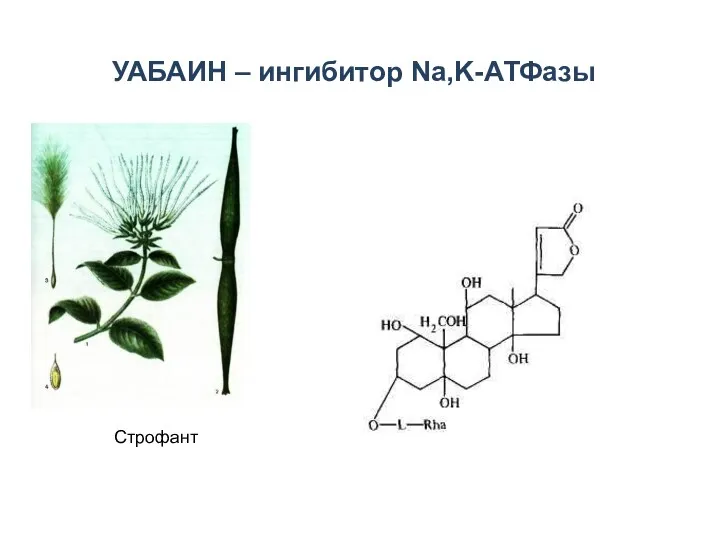
УАБАИН – ингибитор Na,K-АТФазы
Строфант
УАБАИН – ингибитор Na,K-АТФазы
Строфант
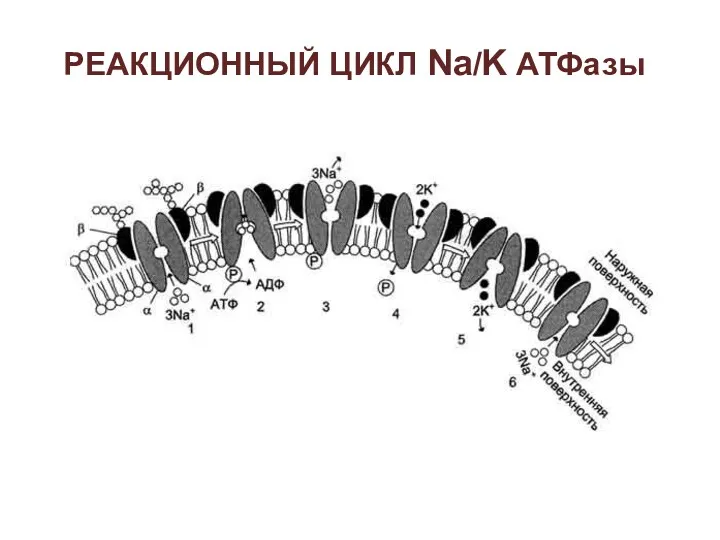
РЕАКЦИОННЫЙ ЦИКЛ Na/K АТФазы
РЕАКЦИОННЫЙ ЦИКЛ Na/K АТФазы
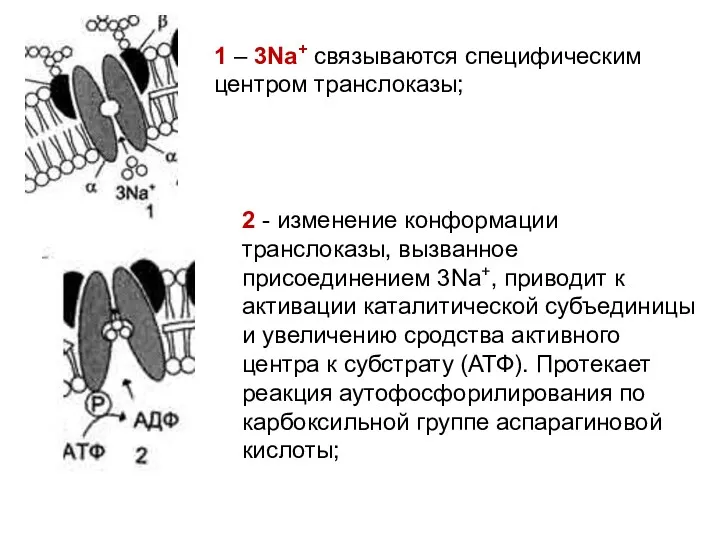
2 - изменение конформации транслоказы, вызванное присоединением 3Na+, приводит к активации
2 - изменение конформации транслоказы, вызванное присоединением 3Na+, приводит к активации

3 - аутофосфорилирование изменяет заряд и конформацию транслоказы, она закрывается с
3 - аутофосфорилирование изменяет заряд и конформацию транслоказы, она закрывается с
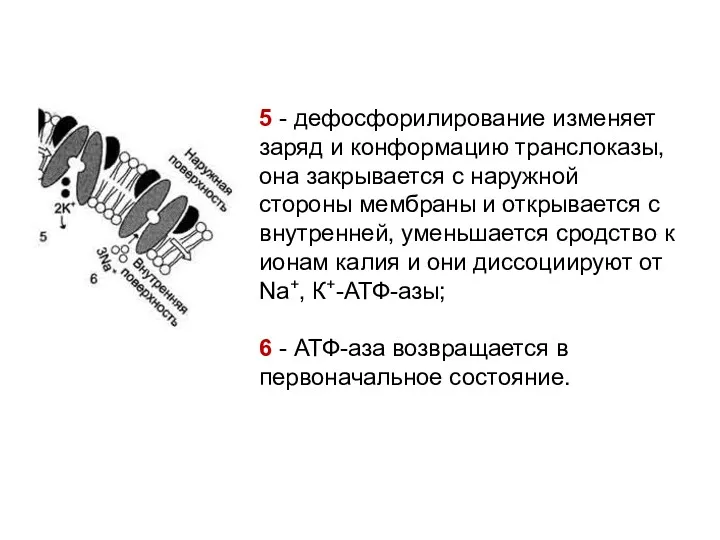
5 - дефосфорилирование изменяет заряд и конформацию транслоказы, она закрывается с
5 - дефосфорилирование изменяет заряд и конформацию транслоказы, она закрывается с

РЕГУЛЯЦИЯ АКТИВНОСТИ Na/K АТФазы
СООТНОШЕНИЕ Na/K И СОДЕРЖАНИЕ АТФ (факторы краткосрочной регуляции)
ФОСФОРИЛИРОВАНИЕ
РЕГУЛЯЦИЯ АКТИВНОСТИ Na/K АТФазы
СООТНОШЕНИЕ Na/K И СОДЕРЖАНИЕ АТФ (факторы краткосрочной регуляции)
ФОСФОРИЛИРОВАНИЕ
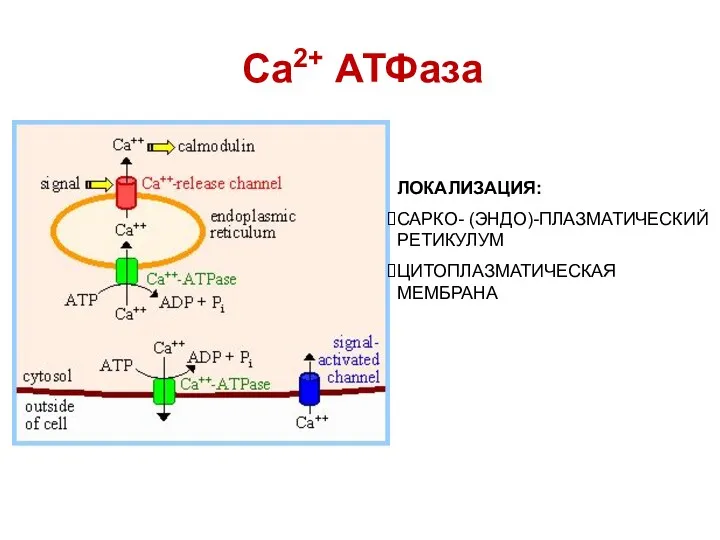
Са2+ АТФаза
ЛОКАЛИЗАЦИЯ:
САРКО- (ЭНДО)-ПЛАЗМАТИЧЕСКИЙ РЕТИКУЛУМ
ЦИТОПЛАЗМАТИЧЕСКАЯ МЕМБРАНА
Са2+ АТФаза
ЛОКАЛИЗАЦИЯ:
САРКО- (ЭНДО)-ПЛАЗМАТИЧЕСКИЙ РЕТИКУЛУМ
ЦИТОПЛАЗМАТИЧЕСКАЯ МЕМБРАНА

ВСЕ Са2+ АТФазы – МОНОМЕРНЫЕ БЕЛКИ, Т.Е. СОСТОЯТ ИЗ ОДНОЙ ПОЛИПЕПТИДНОЙ
ВСЕ Са2+ АТФазы – МОНОМЕРНЫЕ БЕЛКИ, Т.Е. СОСТОЯТ ИЗ ОДНОЙ ПОЛИПЕПТИДНОЙ
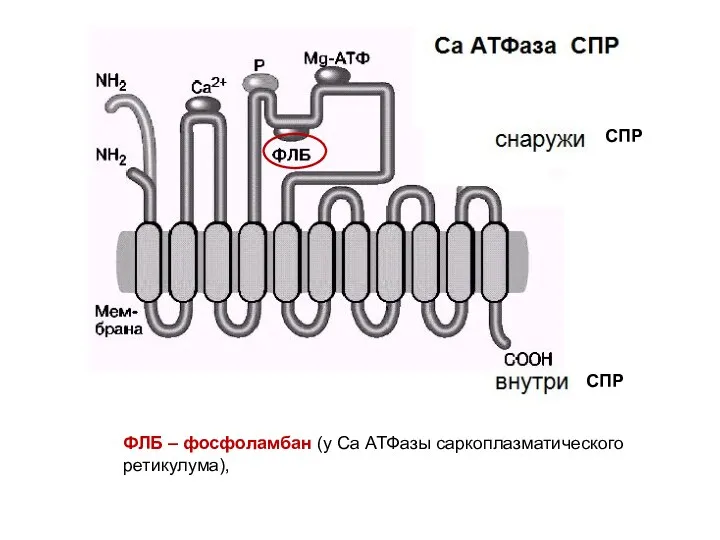
ФЛБ – фосфоламбан (у Са АТФазы саркоплазматического ретикулума),
СПР
СПР
ФЛБ – фосфоламбан (у Са АТФазы саркоплазматического ретикулума),
СПР
СПР
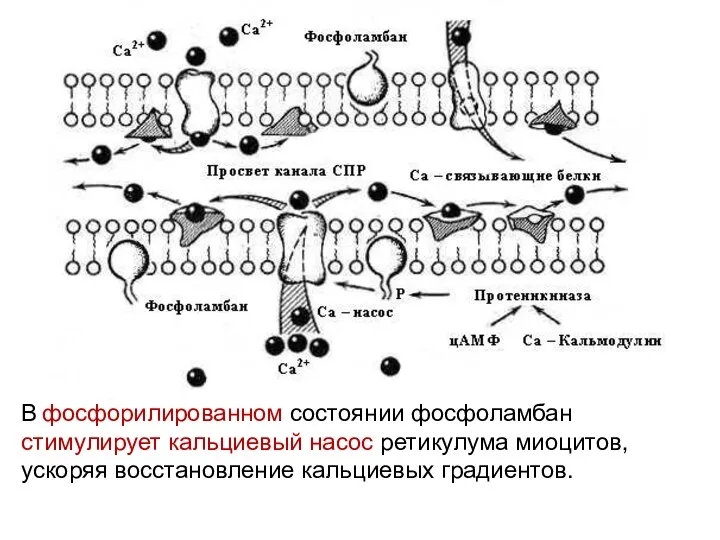
В фосфорилированном состоянии фосфоламбан стимулирует кальциевый насос ретикулума миоцитов, ускоряя восстановление
В фосфорилированном состоянии фосфоламбан стимулирует кальциевый насос ретикулума миоцитов, ускоряя восстановление
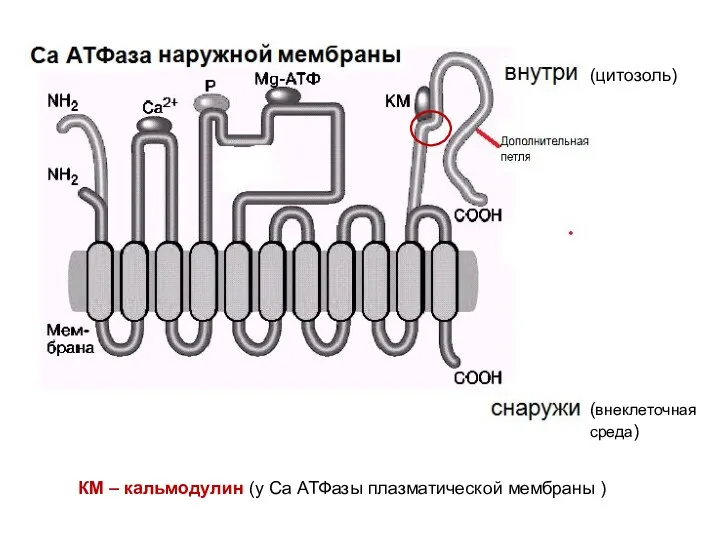
КМ – кальмодулин (у Са АТФазы плазматической мембраны )
(цитозоль)
(внеклеточная среда)
КМ – кальмодулин (у Са АТФазы плазматической мембраны )
(цитозоль)
(внеклеточная среда)
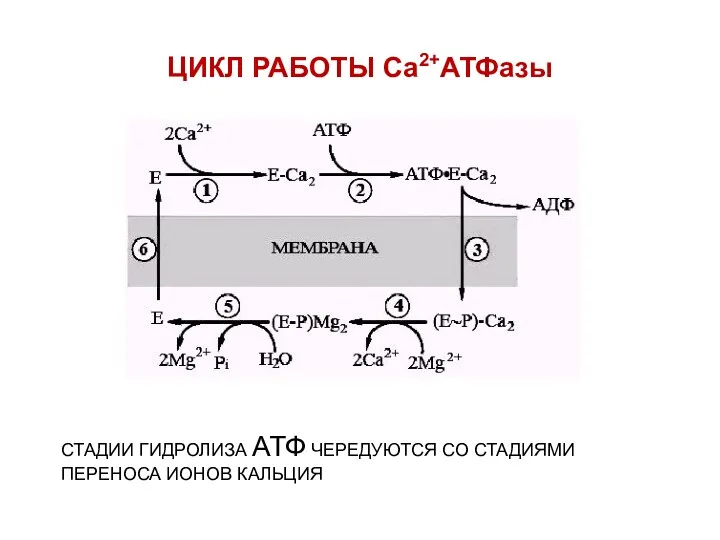
ЦИКЛ РАБОТЫ Са2+АТФазы
СТАДИИ ГИДРОЛИЗА АТФ ЧЕРЕДУЮТСЯ СО СТАДИЯМИ ПЕРЕНОСА ИОНОВ КАЛЬЦИЯ
ЦИКЛ РАБОТЫ Са2+АТФазы
СТАДИИ ГИДРОЛИЗА АТФ ЧЕРЕДУЮТСЯ СО СТАДИЯМИ ПЕРЕНОСА ИОНОВ КАЛЬЦИЯ
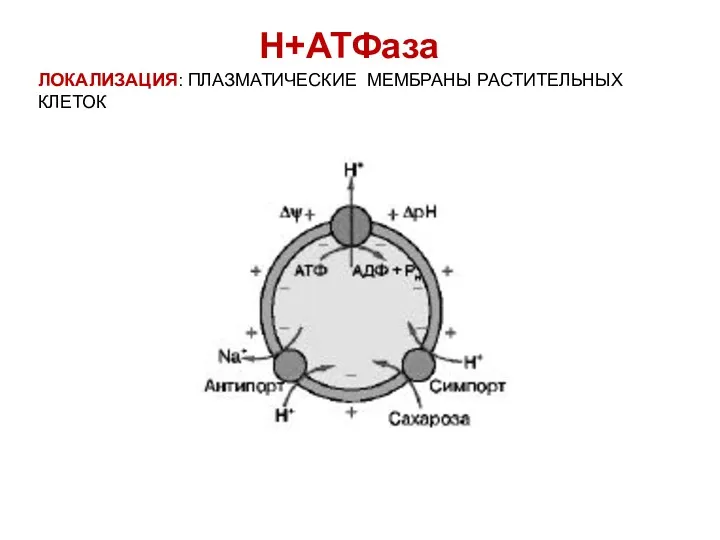
ЛОКАЛИЗАЦИЯ: ПЛАЗМАТИЧЕСКИЕ МЕМБРАНЫ РАСТИТЕЛЬНЫХ КЛЕТОК
Н+АТФаза
ЛОКАЛИЗАЦИЯ: ПЛАЗМАТИЧЕСКИЕ МЕМБРАНЫ РАСТИТЕЛЬНЫХ КЛЕТОК
Н+АТФаза

Н+-АТФаза – это интегральный белок, полипептидная цепь которого десять раз пересекает
Н+-АТФаза – это интегральный белок, полипептидная цепь которого десять раз пересекает


Роль Н+-АТФазы:
поддерживает рН цитоплазмы близкий к нейтральному
создает на мембране разность
Роль Н+-АТФазы:
поддерживает рН цитоплазмы близкий к нейтральному
создает на мембране разность
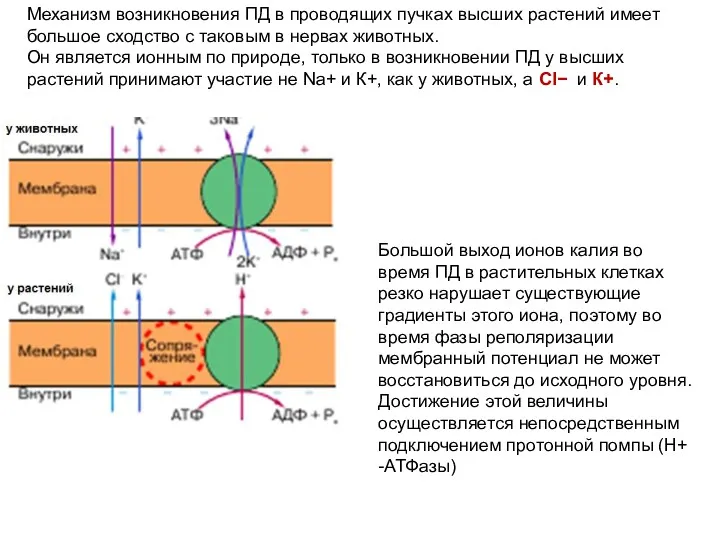
Механизм возникновения ПД в проводящих пучках высших растений имеет большое сходство
Механизм возникновения ПД в проводящих пучках высших растений имеет большое сходство
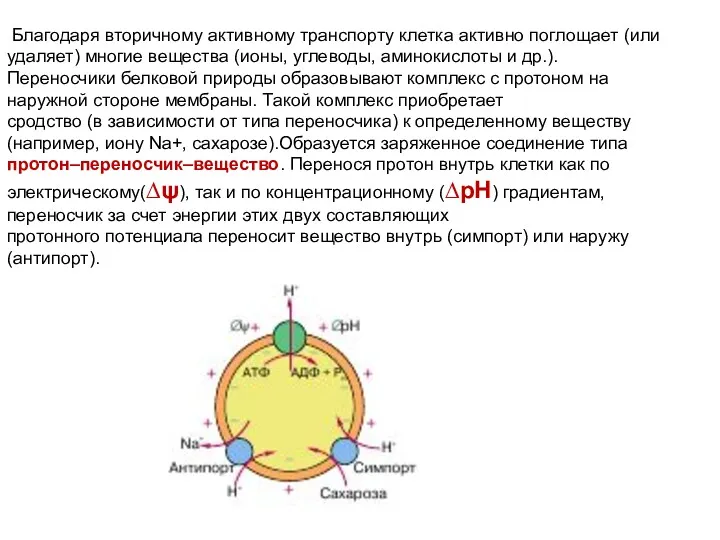
Благодаря вторичному активному транспорту клетка активно поглощает (или удаляет) многие
Благодаря вторичному активному транспорту клетка активно поглощает (или удаляет) многие

ТРАНСПОРТ ВЕЩЕСТВ ЧЕРЕЗ МНОГОМЕМБРАННЫЕ СИСТЕМЫ
ВТОРИЧНЫЙ АКТИВНЫЙ ТРАНСПОРТ
ТРАНСПОРТ ВЕЩЕСТВ ЧЕРЕЗ МНОГОМЕМБРАННЫЕ СИСТЕМЫ
ВТОРИЧНЫЙ АКТИВНЫЙ ТРАНСПОРТ
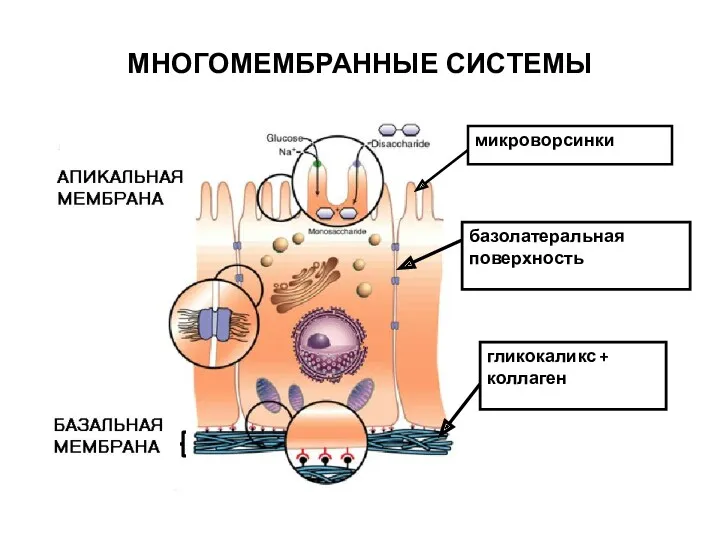
МНОГОМЕМБРАННЫЕ СИСТЕМЫ
гликокаликс + коллаген
микроворсинки
базолатеральная поверхность
МНОГОМЕМБРАННЫЕ СИСТЕМЫ
гликокаликс + коллаген
микроворсинки
базолатеральная поверхность

БАЗАЛЬНАЯ МЕМБРАНА – дополнительная мембрана за пределами плазмалеммы эпителиоцита
ЕЕ СОСТАВ: гликопротеидный
БАЗАЛЬНАЯ МЕМБРАНА – дополнительная мембрана за пределами плазмалеммы эпителиоцита
ЕЕ СОСТАВ: гликопротеидный
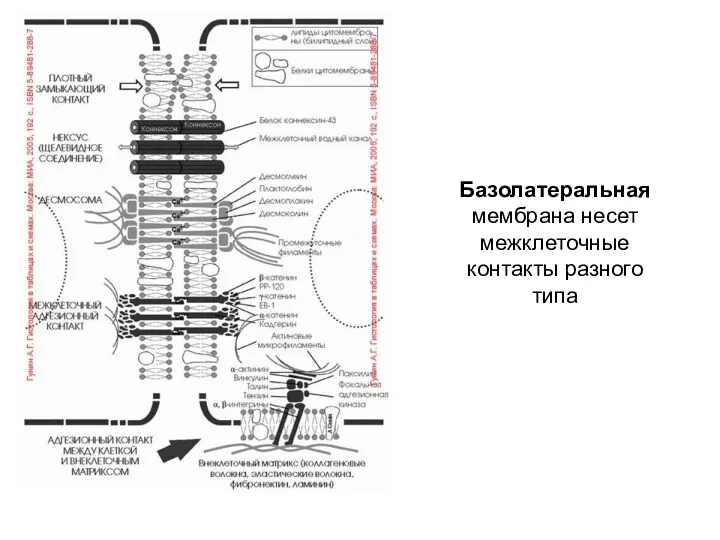
Базолатеральная мембрана несет межклеточные контакты разного типа
Базолатеральная мембрана несет межклеточные контакты разного типа

ЭКСПЕРИМЕНТЫ УССИНГА
Установка Уссинга
Объект исследований
ЭКСПЕРИМЕНТЫ УССИНГА
Установка Уссинга
Объект исследований
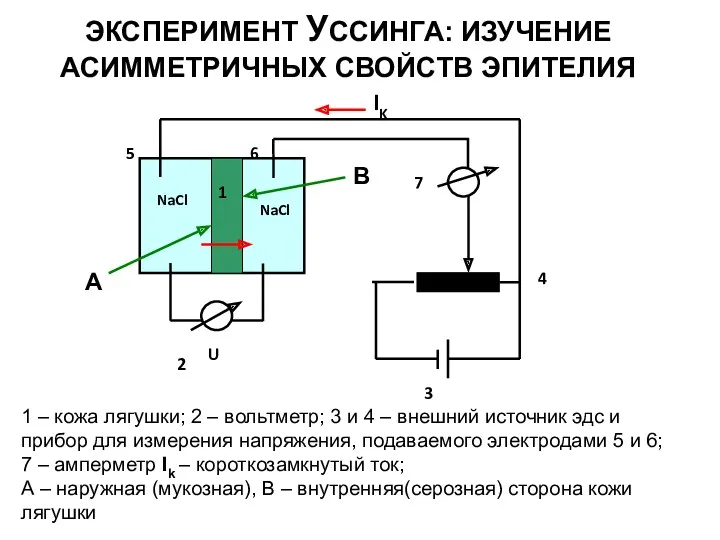
ЭКСПЕРИМЕНТ УССИНГА: ИЗУЧЕНИЕ АСИММЕТРИЧНЫХ СВОЙСТВ ЭПИТЕЛИЯ
1 – кожа лягушки; 2 –
ЭКСПЕРИМЕНТ УССИНГА: ИЗУЧЕНИЕ АСИММЕТРИЧНЫХ СВОЙСТВ ЭПИТЕЛИЯ
1 – кожа лягушки; 2 –
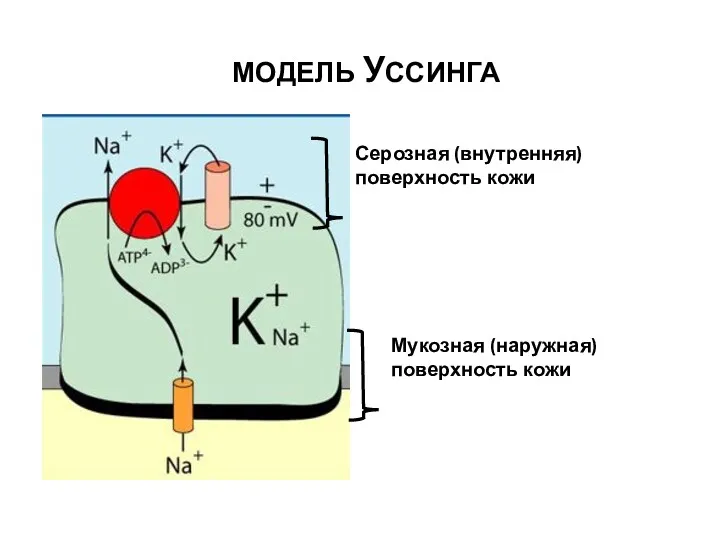
МОДЕЛЬ УССИНГА
Мукозная (наружная)поверхность кожи
Серозная (внутренняя)поверхность кожи
МОДЕЛЬ УССИНГА
Мукозная (наружная)поверхность кожи
Серозная (внутренняя)поверхность кожи
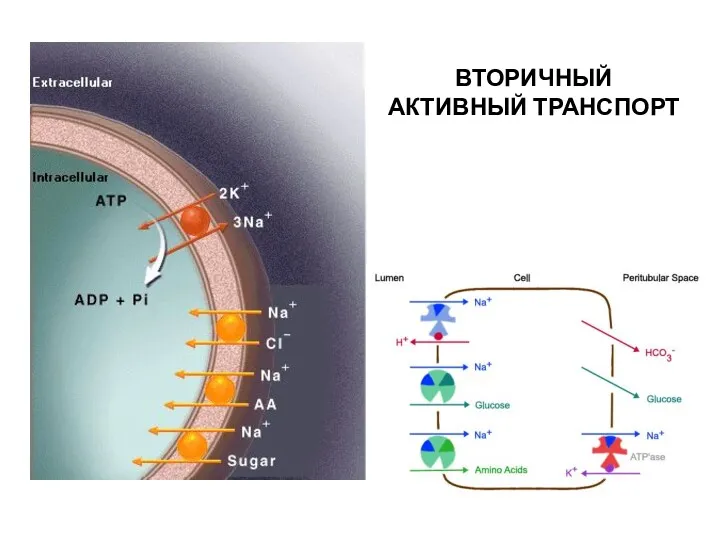
ВТОРИЧНЫЙ АКТИВНЫЙ ТРАНСПОРТ
ВТОРИЧНЫЙ АКТИВНЫЙ ТРАНСПОРТ

ВТОРИЧНЫЙ АКТИВНЫЙ ТРАНСПОРТ
ТРАНСПОРТ САХАРОВ И АМИНОКИСЛОТ ЗА СЧЕТ ЭНЕРГИИ ГРАДИЕНТА Na+
ВТОРИЧНЫЙ АКТИВНЫЙ ТРАНСПОРТ
ТРАНСПОРТ САХАРОВ И АМИНОКИСЛОТ ЗА СЧЕТ ЭНЕРГИИ ГРАДИЕНТА Na+

Уравнение для транспорта сахаров
Jmax = 12 мкмоль / м2 с –
Уравнение для транспорта сахаров
Jmax = 12 мкмоль / м2 с –
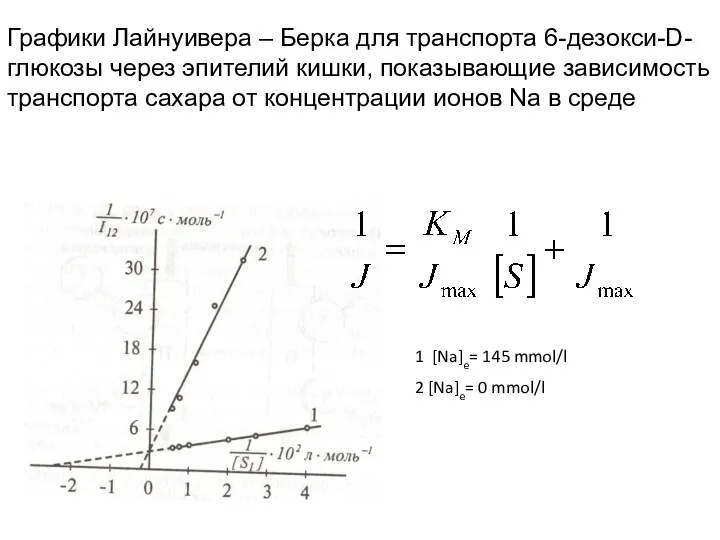
Графики Лайнуивера – Берка для транспорта 6-дезокси-D-глюкозы через эпителий кишки, показывающие
Графики Лайнуивера – Берка для транспорта 6-дезокси-D-глюкозы через эпителий кишки, показывающие
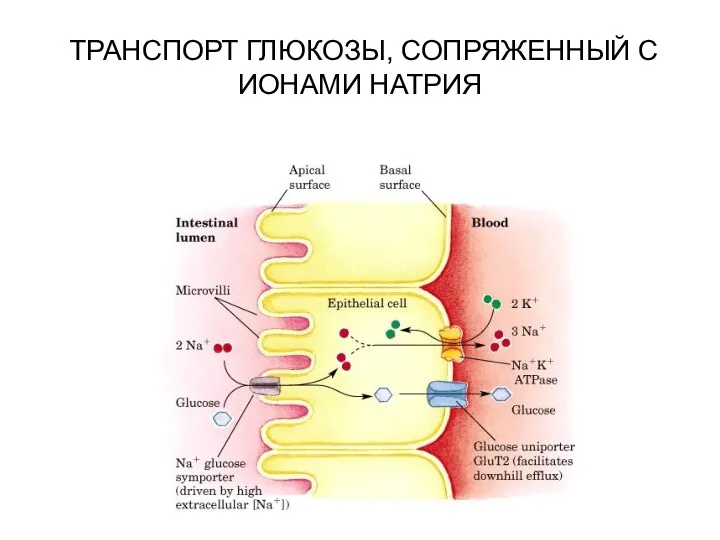
ТРАНСПОРТ ГЛЮКОЗЫ, СОПРЯЖЕННЫЙ С ИОНАМИ НАТРИЯ
ТРАНСПОРТ ГЛЮКОЗЫ, СОПРЯЖЕННЫЙ С ИОНАМИ НАТРИЯ
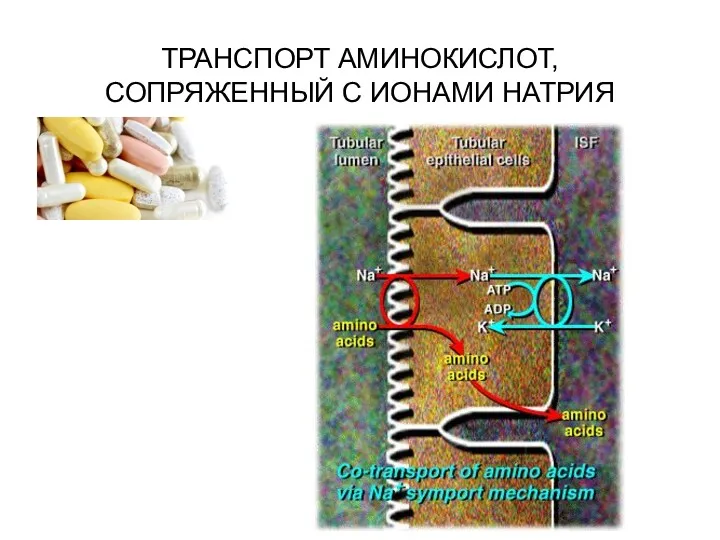
ТРАНСПОРТ АМИНОКИСЛОТ, СОПРЯЖЕННЫЙ С ИОНАМИ НАТРИЯ
ТРАНСПОРТ АМИНОКИСЛОТ, СОПРЯЖЕННЫЙ С ИОНАМИ НАТРИЯ

 Өсімдіктердің вегетативті көбею тәсілдерін зерттеу
Өсімдіктердің вегетативті көбею тәсілдерін зерттеу Молекулярні механізми впливу канцерогенів на організм людини
Молекулярні механізми впливу канцерогенів на організм людини Учение о развитии зародыша - эмбриология
Учение о развитии зародыша - эмбриология Голосеменные растения
Голосеменные растения Презентации к урокам 9 - 11 класс
Презентации к урокам 9 - 11 класс Викторина по биологии Что? Где? Когда?
Викторина по биологии Что? Где? Когда? Немного о бобрах
Немного о бобрах Технологія виробництва м’яса бройлерів
Технологія виробництва м’яса бройлерів Генетика, как наука. Взаимосвязь процессов наследственности и изменчивости
Генетика, как наука. Взаимосвязь процессов наследственности и изменчивости Модификациялық өзгергіштік
Модификациялық өзгергіштік Сырдарияның гидробиологиялық режимі
Сырдарияның гидробиологиялық режимі Вегетативное размножение растений
Вегетативное размножение растений Домашние кошки
Домашние кошки Представители парнокопытных
Представители парнокопытных Формы естественного отбора
Формы естественного отбора Основные группы рыб, их роль в природе и практическое значение
Основные группы рыб, их роль в природе и практическое значение Отряд Чешуйчатые. Подотряд Ящерицы
Отряд Чешуйчатые. Подотряд Ящерицы Биометрические системы контроля доступа
Биометрические системы контроля доступа Всероссийская проверочная работа по биологии. 5 класс, 3 вариант
Всероссийская проверочная работа по биологии. 5 класс, 3 вариант Генно-инженерно-модифицированные организмы. Методы детекции и идентификации
Генно-инженерно-модифицированные организмы. Методы детекции и идентификации Здоровьесберегающий подход к выбору туалетного мыла
Здоровьесберегающий подход к выбору туалетного мыла Бактерии
Бактерии Наследственность и изменчивость как основа способности к развитию и эволюции
Наследственность и изменчивость как основа способности к развитию и эволюции 10 самых необычных растений мира
10 самых необычных растений мира Царство Вирусы
Царство Вирусы Фауна, животные (урок 14)
Фауна, животные (урок 14) Цианеи (сине-зелёные водоросли)
Цианеи (сине-зелёные водоросли) Основы гистологии. Ткани, виды тканей, соединительная ткань
Основы гистологии. Ткани, виды тканей, соединительная ткань