- Клеточное дыхание. Лекция 3

Содержание
- 2. ЛЕКЦИЯ_3 КЛЕТОЧНОЕ ДЫХАНИЕ
- 3. ПЛАН ЛЕКЦИИ: Клеточное дыхание Дыхание при терморегуляции Дыхание как механизм образования необходимых соединений Дыхание как механизм
- 4. ОСНОВНЫЕ ПРОЦЕССЫ, ОБЕСПЕЧИВАЮЩИЕ КЛЕТКУ ЭНЕРГИЕЙ
- 5. по отношению к энергетическим источникам Хемотрофы используют для синтеза АТФ энергию, освобождаемую в результате химических реакций,
- 6. Синтез молекул АТФ из АДФ и фосфатов может происходить двумя способами: 1. фосфорилированием в дыхательной или
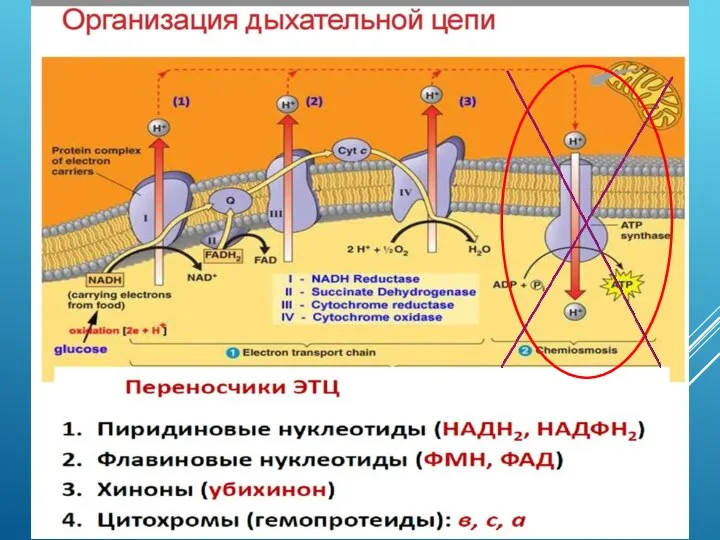
- 7. Клеточное дыхание – это окислительный распад органических питательных веществ с участием кислорода, сопровождающийся образованием химически активных
- 8. ОКСИДАЗЫ Клеточным дыханием называют совокупность протекающих в каждой клетке ферментативных процессов, в результате которых молекулы углеводов,
- 9. Процесс дыхания катализируется ферментами оксидазами: (1) более редкий случай – внедрение одного из атомов молекулы кислорода
- 10. ГЛИКОЛИЗ Ферменты: 1 - Гексокиназа 2 - 2 - Глюкозо-6-фосфатизомераза 3 - 6-3 - 6-Фосфофруктокиназа 4
- 11. Важнейший представитель оксидаз - цитохромоксидаза (цитохром с-оксидаза, цитохром aa3) локализован во внутренней мембране митохондрий животных, растений
- 12. Цитохромоксидаза – конечный фермент дыхательной цепи, последовательности ферментов и коферментов – переносчиков электронов от субстратов дыхания
- 13. ЦИТОХРОМОКСИДАЗА Каталитический центр фермента содержит гемы и медные комплексы. У эукариотов оксидазы находятся во внутренней митохондриальной
- 14. ЦИТОХРОМОКСИДАЗА, КОТОРАЯ НЕ ЛЮБИТ ПРОТОНЫ http://www.pnas.org/content/112/25/7695
- 16. Митохондрии взрослого человека перекачивают через свои мембраны около 500 г ионов Н+ в день, образуя протонный
- 17. Функции клеточного дыхания:
- 19. ЗАПАСАНИЕ ПОЛЕЗНОЙ ЭНЕРГИИ Основная функция в количественном отношении: большая часть кислорода поглощается клеткой для окисления субстратов
- 20. ГИДРОЛИЗ АТФ ИСПОЛЬЗУЕТСЯ ДЛЯ ОБЕСПЕЧЕНИЯ: Митохондрии взрослого человека перекачивают через свои мембраны около 500 г ионов
- 21. Альтернативные функции клеточного дыхания: образование тепла в целях терморегуляции.
- 22. РАССЕЯНИЕ ЭНЕРГИИ ДЫХАНИЯ ПРИ ТЕРМОРЕГУЛЯЦИИ Почти вся энергия, извлекаемая при дыхании путем окисления субстратов кислородом, в
- 23. МЕХАНИЗМ ОБРАЗОВАНИЯ ТЕПЛА ПРИ ХОЛОДОВОЙ ДРОЖИ: Сначала накапливается энергия в виде АТФ внутри митохондрий, затем транспортируется
- 25. Образование и использование протонного потенциала
- 26. Причина повышения протонной проводимости митохондриальной мембраны - резкое возрастание концентрации свободных жирных кислот (пальмитиновой и стеариновой,
- 27. МЕХАНИЗМ РАЗОБЩАЮ-ЩЕГО ДЕЙСТВИЯ ЖИРНЫХ КИСЛОТ
- 28. Главная функция АТФ /АДФ-антипортера состоит в обмене аниона внешнего (цитозольного) АДФ на внутримито-хондриальный анион АТФ, образованный
- 30. БУРЫЙ ЖИР Митохондрии бурого жира имеют большой избыток дыхательных ферментов по сравнению с Н+-АТФ-синтазой Около 10%
- 33. ДВА ФИЗИОЛОГИЧЕСКИХ СПОСОБА ВЫЗВАТЬ РАЗОБЩЕНИЕ ДЫХАНИЯ И ФОСФОРИЛИРОВАНИЯ В БУРОМ ЖИРЕ: Один из них специфичен для
- 34. ТЕРМОРЕГУЛЯТОРНОЕ РАЗОБЩЕНИЕ У РАСТЕНИЙ в 1788 году Ж.Б. Ламарк - в холодную погоду цветы некоторых лилейных
- 35. ВОСТОЧНАЯ КАПУСТА В результате скорость дыхания возрастает до 100 раз, а градиент температуры между цветком и
- 37. ДЫХАНИЕ КАК МЕХАНИЗМ ОБРАЗОВАНИЯ ПОЛЕЗНЫХ СОЕДИНЕНИЙ ФУНКЦИИ ДЫХАНИЯ, ИГРАЮЩИЕ РОЛЬ В ПРЕВРАЩЕНИИ ВЕЩЕСТВ: Образование (синтез) соединений
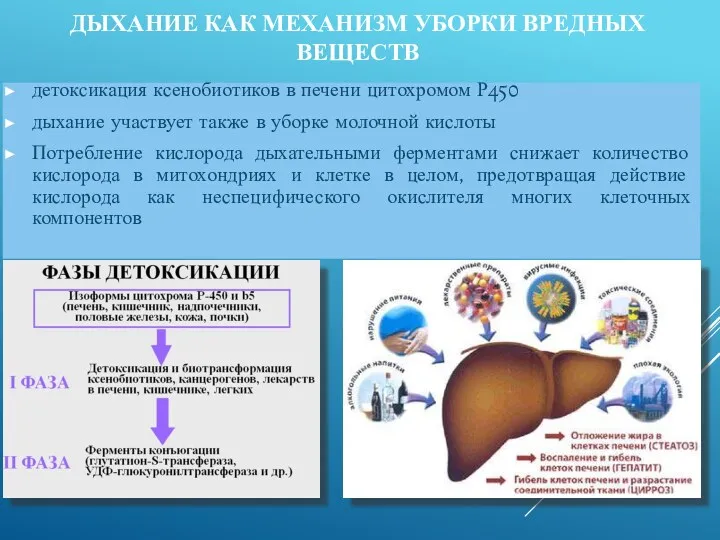
- 38. ДЫХАНИЕ КАК МЕХАНИЗМ УБОРКИ ВРЕДНЫХ ВЕЩЕСТВ детоксикация ксенобиотиков в печени цитохромом Р450 дыхание участвует также в
- 39. ДЫХАНИЕ КАК МЕХАНИЗМ УБОРКИ ВРЕДНЫХ ВЕЩЕСТВ (особенности) У дрожжей описан специальный фермент – цитохром b2, переносящий
- 40. ДЫХАНИЕ КАК МЕХАНИЗМ, УСКОРЯЮЩИЙ ЭВОЛЮЦИЮ Окисление ДНК активными формами кислорода - важнейший инструмент природного мутагенеза. Возможно,
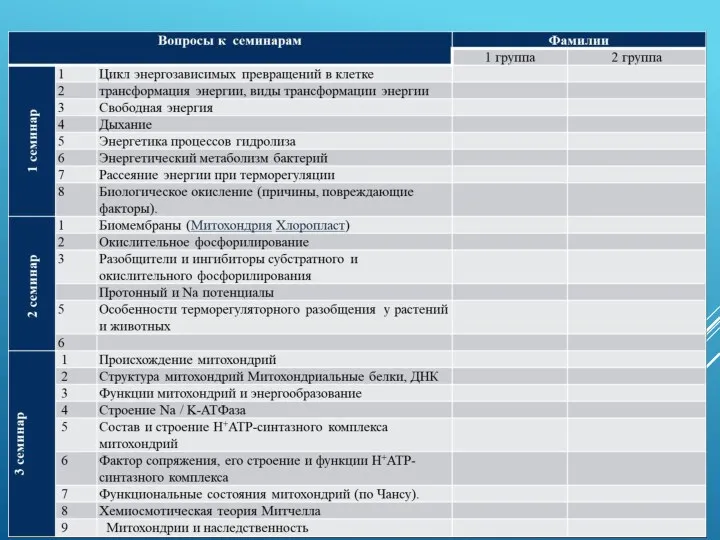
- 41. ВОПРОСЫ К СЕМИНАРАМ
- 43. Скачать презентацию

ЛЕКЦИЯ_3
КЛЕТОЧНОЕ ДЫХАНИЕ
ЛЕКЦИЯ_3
КЛЕТОЧНОЕ ДЫХАНИЕ

ПЛАН ЛЕКЦИИ:
Клеточное дыхание
Дыхание при терморегуляции
Дыхание как механизм образования необходимых соединений
Дыхание как
ПЛАН ЛЕКЦИИ:
Клеточное дыхание
Дыхание при терморегуляции
Дыхание как механизм образования необходимых соединений
Дыхание как
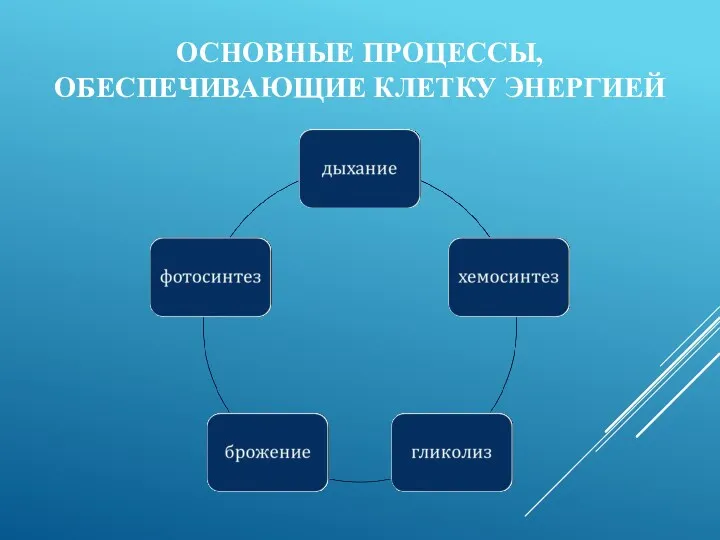
ОСНОВНЫЕ ПРОЦЕССЫ, ОБЕСПЕЧИВАЮЩИЕ КЛЕТКУ ЭНЕРГИЕЙ
ОСНОВНЫЕ ПРОЦЕССЫ, ОБЕСПЕЧИВАЮЩИЕ КЛЕТКУ ЭНЕРГИЕЙ
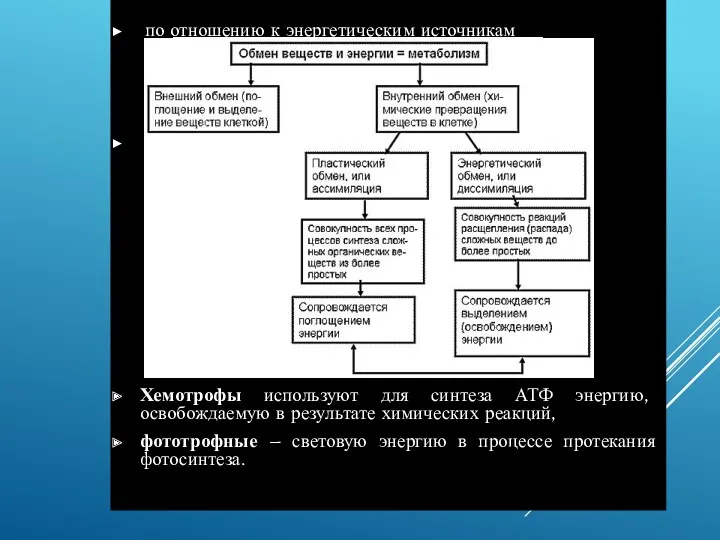
по отношению к энергетическим источникам
Хемотрофы используют для синтеза АТФ энергию, освобождаемую
по отношению к энергетическим источникам
Хемотрофы используют для синтеза АТФ энергию, освобождаемую

Синтез молекул АТФ из АДФ и фосфатов может происходить двумя способами:
1.
Синтез молекул АТФ из АДФ и фосфатов может происходить двумя способами: 1.

Клеточное дыхание – это окислительный распад органических питательных веществ с участием
Клеточное дыхание – это окислительный распад органических питательных веществ с участием
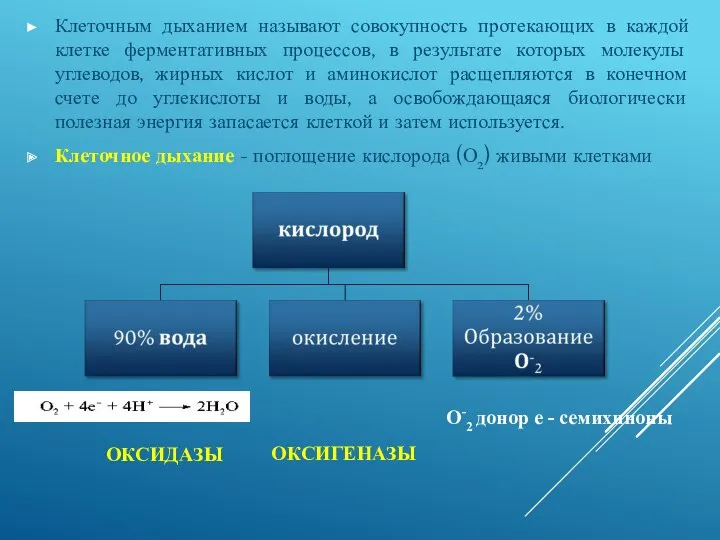
ОКСИДАЗЫ
Клеточным дыханием называют совокупность протекающих в каждой клетке ферментативных процессов, в
ОКСИДАЗЫ
Клеточным дыханием называют совокупность протекающих в каждой клетке ферментативных процессов, в

Процесс дыхания катализируется ферментами оксидазами:
(1)
более редкий случай – внедрение одного
Процесс дыхания катализируется ферментами оксидазами:
(1)
более редкий случай – внедрение одного
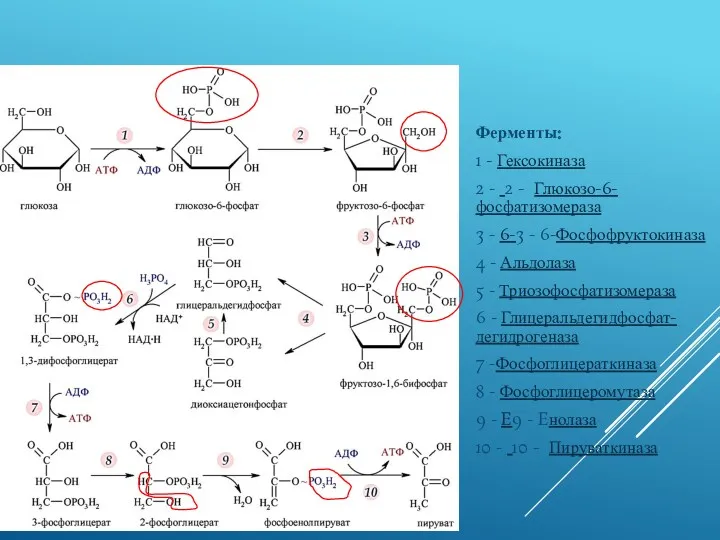
ГЛИКОЛИЗ
Ферменты:
1 - Гексокиназа
2 - 2 - Глюкозо-6-фосфатизомераза
3 - 6-3 - 6-Фосфофруктокиназа
4 - Альдолаза
5 - Триозофосфатизомераза
6 - Глицеральдегидфосфат-дегидрогеназа
7
ГЛИКОЛИЗ
Ферменты:
1 - Гексокиназа
2 - 2 - Глюкозо-6-фосфатизомераза
3 - 6-3 - 6-Фосфофруктокиназа
4 - Альдолаза
5 - Триозофосфатизомераза
6 - Глицеральдегидфосфат-дегидрогеназа
7

Важнейший представитель оксидаз - цитохромоксидаза (цитохром с-оксидаза, цитохром aa3)
локализован во внутренней
Важнейший представитель оксидаз - цитохромоксидаза (цитохром с-оксидаза, цитохром aa3)
локализован во внутренней
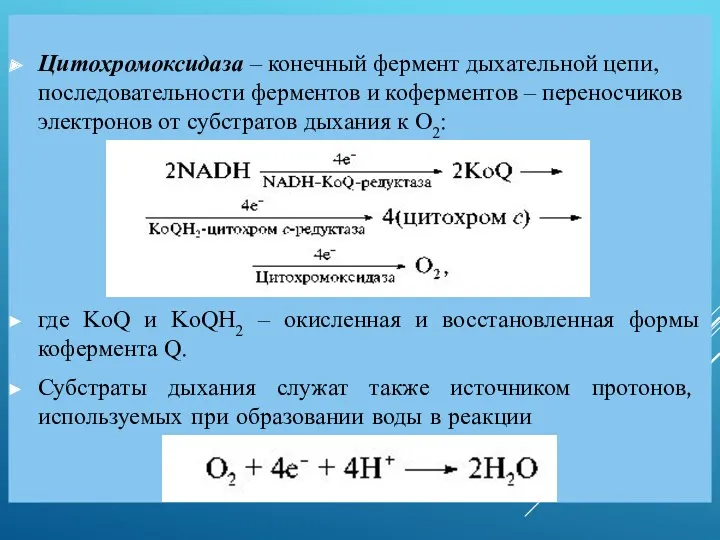
Цитохромоксидаза – конечный фермент дыхательной цепи, последовательности ферментов и коферментов –
Цитохромоксидаза – конечный фермент дыхательной цепи, последовательности ферментов и коферментов –
ЦИТОХРОМОКСИДАЗА
Каталитический центр фермента содержит гемы и медные комплексы.
У эукариотов оксидазы находятся во внутренней
ЦИТОХРОМОКСИДАЗА
Каталитический центр фермента содержит гемы и медные комплексы.
У эукариотов оксидазы находятся во внутренней

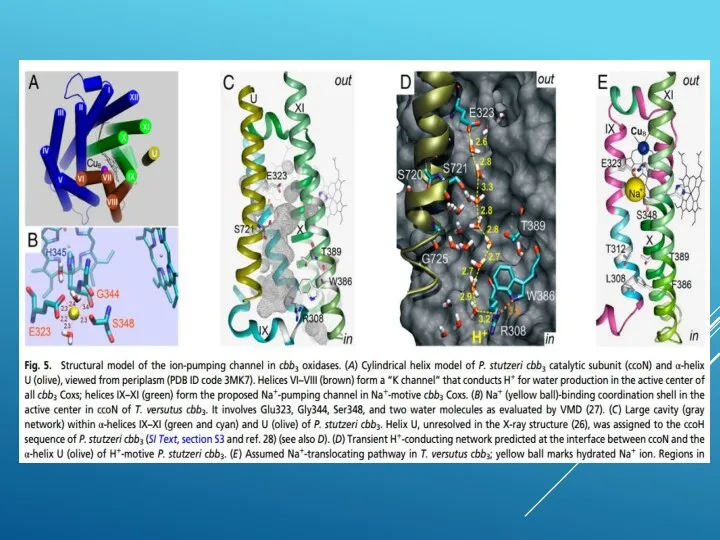
ЦИТОХРОМОКСИДАЗА, КОТОРАЯ НЕ ЛЮБИТ ПРОТОНЫ
http://www.pnas.org/content/112/25/7695
ЦИТОХРОМОКСИДАЗА, КОТОРАЯ НЕ ЛЮБИТ ПРОТОНЫ
http://www.pnas.org/content/112/25/7695

Митохондрии взрослого человека перекачивают через свои мембраны около 500 г ионов
Митохондрии взрослого человека перекачивают через свои мембраны около 500 г ионов
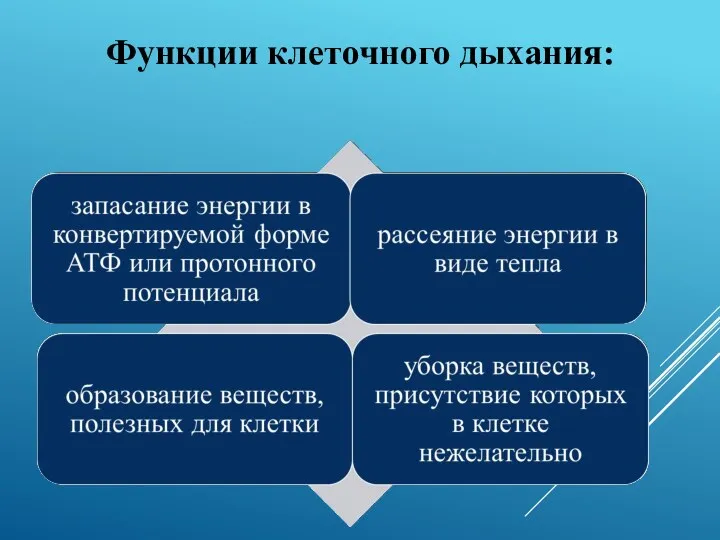
Функции клеточного дыхания:
Функции клеточного дыхания:


ЗАПАСАНИЕ ПОЛЕЗНОЙ ЭНЕРГИИ
Основная функция в количественном отношении: большая часть кислорода поглощается
ЗАПАСАНИЕ ПОЛЕЗНОЙ ЭНЕРГИИ
Основная функция в количественном отношении: большая часть кислорода поглощается

ГИДРОЛИЗ АТФ ИСПОЛЬЗУЕТСЯ ДЛЯ ОБЕСПЕЧЕНИЯ:
Митохондрии взрослого человека перекачивают через свои
ГИДРОЛИЗ АТФ ИСПОЛЬЗУЕТСЯ ДЛЯ ОБЕСПЕЧЕНИЯ:
Митохондрии взрослого человека перекачивают через свои

Альтернативные функции клеточного дыхания:
образование тепла в целях терморегуляции.
Альтернативные функции клеточного дыхания:
образование тепла в целях терморегуляции.

РАССЕЯНИЕ ЭНЕРГИИ ДЫХАНИЯ ПРИ ТЕРМОРЕГУЛЯЦИИ
Почти вся энергия, извлекаемая при дыхании путем
РАССЕЯНИЕ ЭНЕРГИИ ДЫХАНИЯ ПРИ ТЕРМОРЕГУЛЯЦИИ
Почти вся энергия, извлекаемая при дыхании путем

МЕХАНИЗМ ОБРАЗОВАНИЯ ТЕПЛА ПРИ ХОЛОДОВОЙ ДРОЖИ:
Сначала накапливается энергия в виде АТФ
МЕХАНИЗМ ОБРАЗОВАНИЯ ТЕПЛА ПРИ ХОЛОДОВОЙ ДРОЖИ:
Сначала накапливается энергия в виде АТФ
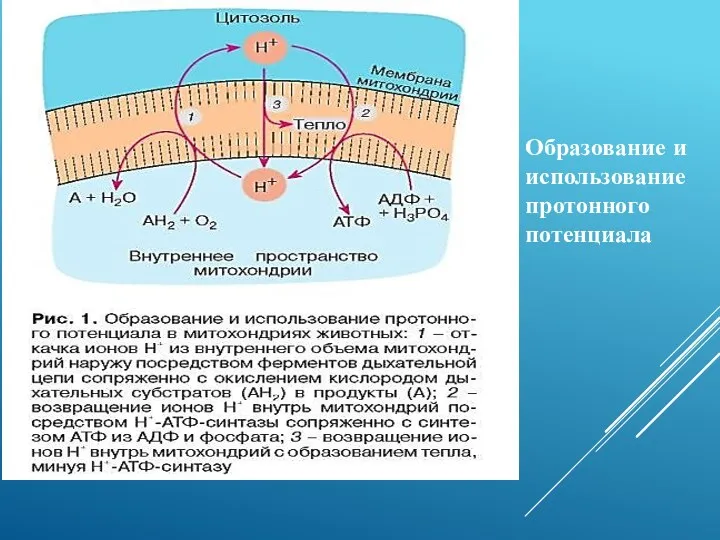
Образование и использование протонного потенциала
Образование и использование протонного потенциала

Причина повышения протонной проводимости митохондриальной мембраны - резкое возрастание концентрации свободных
Причина повышения протонной проводимости митохондриальной мембраны - резкое возрастание концентрации свободных
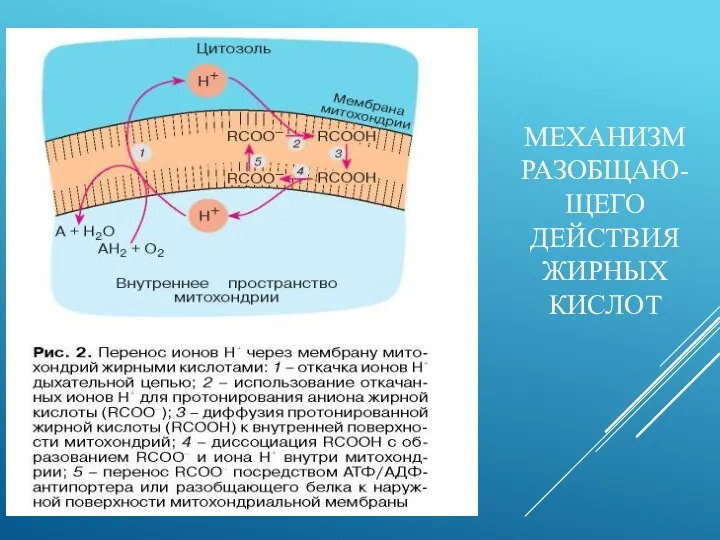
МЕХАНИЗМ РАЗОБЩАЮ-ЩЕГО ДЕЙСТВИЯ ЖИРНЫХ КИСЛОТ
МЕХАНИЗМ РАЗОБЩАЮ-ЩЕГО ДЕЙСТВИЯ ЖИРНЫХ КИСЛОТ
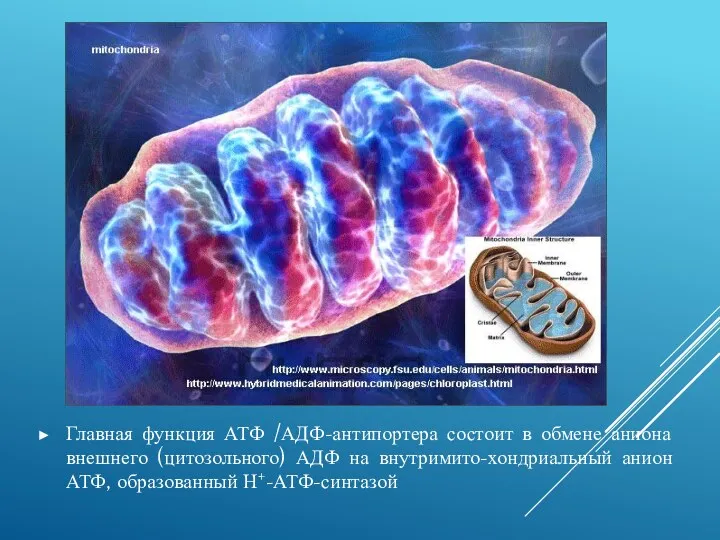
Главная функция АТФ /АДФ-антипортера состоит в обмене аниона внешнего (цитозольного) АДФ
Главная функция АТФ /АДФ-антипортера состоит в обмене аниона внешнего (цитозольного) АДФ
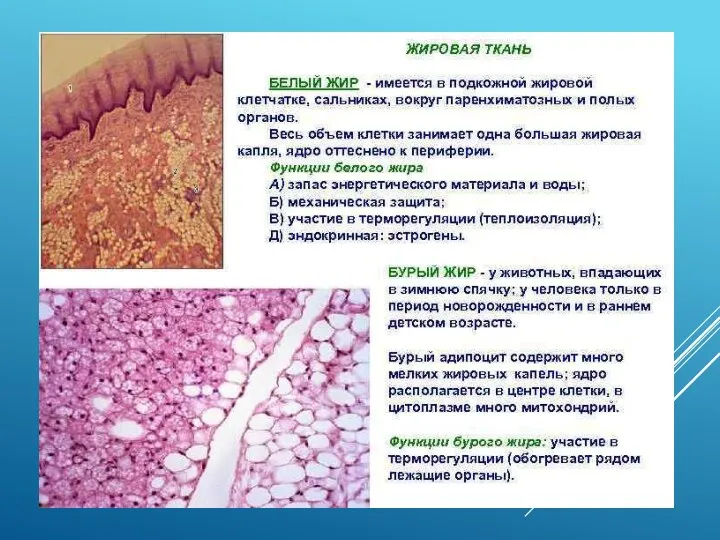
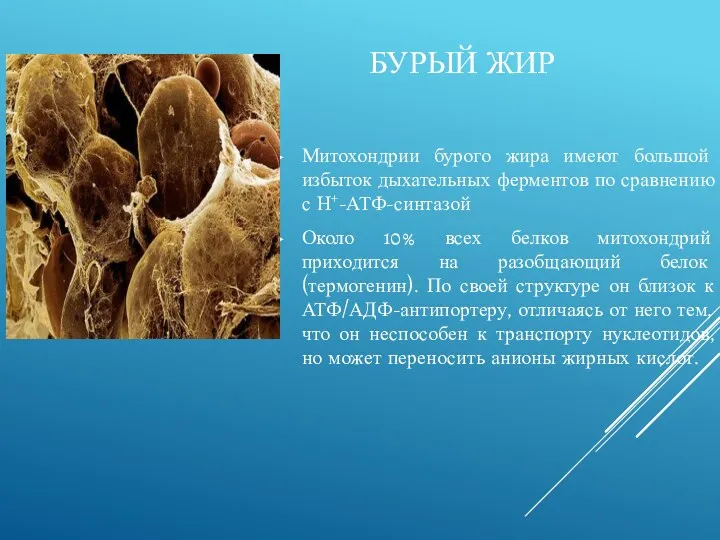
БУРЫЙ ЖИР
Митохондрии бурого жира имеют большой избыток дыхательных ферментов по сравнению
БУРЫЙ ЖИР
Митохондрии бурого жира имеют большой избыток дыхательных ферментов по сравнению
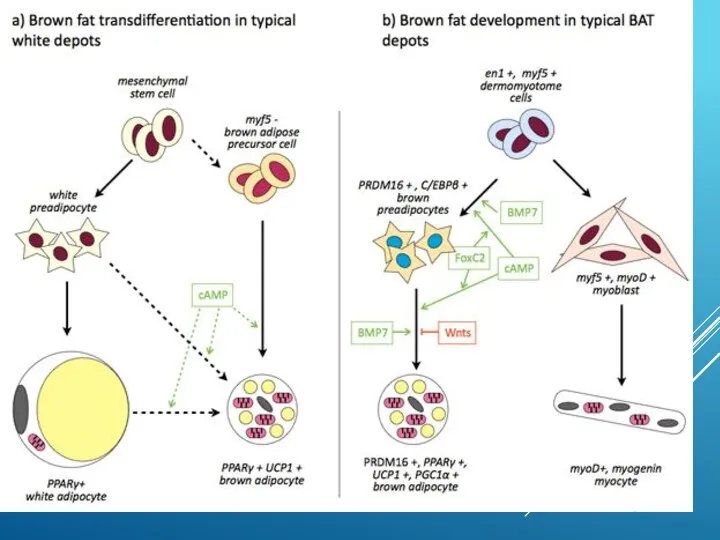
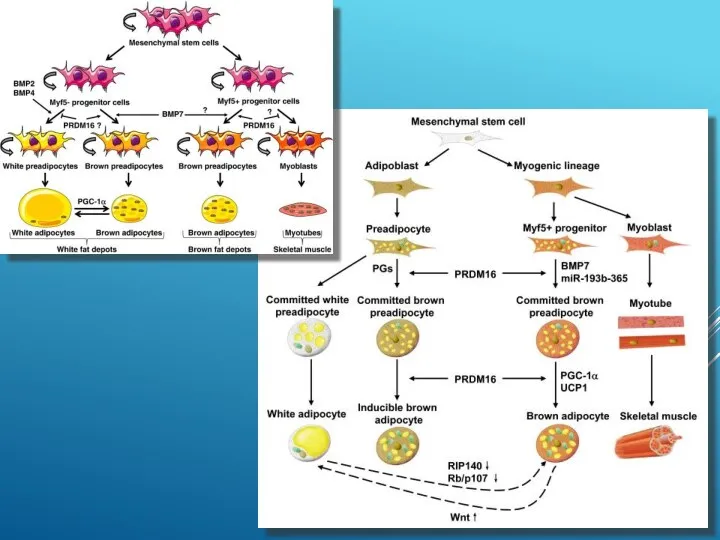

ДВА ФИЗИОЛОГИЧЕСКИХ СПОСОБА ВЫЗВАТЬ РАЗОБЩЕНИЕ ДЫХАНИЯ И ФОСФОРИЛИРОВАНИЯ В БУРОМ ЖИРЕ:
Один
ДВА ФИЗИОЛОГИЧЕСКИХ СПОСОБА ВЫЗВАТЬ РАЗОБЩЕНИЕ ДЫХАНИЯ И ФОСФОРИЛИРОВАНИЯ В БУРОМ ЖИРЕ:
Один

ТЕРМОРЕГУЛЯТОРНОЕ РАЗОБЩЕНИЕ У РАСТЕНИЙ
в 1788 году Ж.Б. Ламарк - в холодную
ТЕРМОРЕГУЛЯТОРНОЕ РАЗОБЩЕНИЕ У РАСТЕНИЙ
в 1788 году Ж.Б. Ламарк - в холодную

ВОСТОЧНАЯ КАПУСТА
В результате скорость дыхания возрастает до 100 раз, а градиент
ВОСТОЧНАЯ КАПУСТА
В результате скорость дыхания возрастает до 100 раз, а градиент


ДЫХАНИЕ КАК МЕХАНИЗМ ОБРАЗОВАНИЯ ПОЛЕЗНЫХ СОЕДИНЕНИЙ
ФУНКЦИИ ДЫХАНИЯ, ИГРАЮЩИЕ РОЛЬ В ПРЕВРАЩЕНИИ
ДЫХАНИЕ КАК МЕХАНИЗМ ОБРАЗОВАНИЯ ПОЛЕЗНЫХ СОЕДИНЕНИЙ
ФУНКЦИИ ДЫХАНИЯ, ИГРАЮЩИЕ РОЛЬ В ПРЕВРАЩЕНИИ
ДЫХАНИЕ КАК МЕХАНИЗМ УБОРКИ ВРЕДНЫХ ВЕЩЕСТВ
детоксикация ксенобиотиков в печени цитохромом Р450
дыхание
ДЫХАНИЕ КАК МЕХАНИЗМ УБОРКИ ВРЕДНЫХ ВЕЩЕСТВ
детоксикация ксенобиотиков в печени цитохромом Р450
дыхание

ДЫХАНИЕ КАК МЕХАНИЗМ УБОРКИ ВРЕДНЫХ ВЕЩЕСТВ (особенности)
У дрожжей описан специальный фермент
ДЫХАНИЕ КАК МЕХАНИЗМ УБОРКИ ВРЕДНЫХ ВЕЩЕСТВ (особенности)
У дрожжей описан специальный фермент

ДЫХАНИЕ КАК МЕХАНИЗМ, УСКОРЯЮЩИЙ ЭВОЛЮЦИЮ
Окисление ДНК активными формами кислорода - важнейший
ДЫХАНИЕ КАК МЕХАНИЗМ, УСКОРЯЮЩИЙ ЭВОЛЮЦИЮ
Окисление ДНК активными формами кислорода - важнейший
ВОПРОСЫ К СЕМИНАРАМ
ВОПРОСЫ К СЕМИНАРАМ
 Обмен веществ и энергии в клетке
Обмен веществ и энергии в клетке презентация по биологии Ароморфозы и идиоадаптации
презентация по биологии Ароморфозы и идиоадаптации Бионика: природа – главный патентнтообладатель
Бионика: природа – главный патентнтообладатель Внутрішнє середовище організму. Кров, її склад та функції
Внутрішнє середовище організму. Кров, її склад та функції Коферменттердің құрылысы мен биологиялық функциясы
Коферменттердің құрылысы мен биологиялық функциясы Теории возникновения жизни на Земле и самой Земли
Теории возникновения жизни на Земле и самой Земли Основные направления и перспективы биотехнологии
Основные направления и перспективы биотехнологии Приспособленность организмов к среде обитания
Приспособленность организмов к среде обитания Движение крови и лимфы
Движение крови и лимфы Структура и функции хромосом
Структура и функции хромосом Анатомия и физиология органа зрения
Анатомия и физиология органа зрения Ауыз- қуысының қорғаныс механизмі
Ауыз- қуысының қорғаныс механизмі Частная биотехнология лекарственных средств
Частная биотехнология лекарственных средств Исследовательский проект на тему: Исследование действия антибиотиков на микроорганизмы
Исследовательский проект на тему: Исследование действия антибиотиков на микроорганизмы Петуния. Выращивание и уход за петунией
Петуния. Выращивание и уход за петунией Мир комнатных растений. Окружающий мир 2 класс
Мир комнатных растений. Окружающий мир 2 класс Вегетативное размножение растений
Вегетативное размножение растений Құстардың тыныс алуы және газ алмасуы
Құстардың тыныс алуы және газ алмасуы Перспектива применения злаковых растений в озеленении г. Саратова
Перспектива применения злаковых растений в озеленении г. Саратова Мордовский государственный заповедник имени П.Г.Смидовича
Мордовский государственный заповедник имени П.Г.Смидовича Бактерии в геохимических круговоротах. Палеомикробиология и космическая микробиология. Нефтяная микробиология
Бактерии в геохимических круговоротах. Палеомикробиология и космическая микробиология. Нефтяная микробиология Түйеқұстар отряды
Түйеқұстар отряды Дослідження процесу росту вегетативних органів (пагін і корінь): пшениці, квасолі, насіння соняшника
Дослідження процесу росту вегетативних органів (пагін і корінь): пшениці, квасолі, насіння соняшника Ядро эукариотической клетки
Ядро эукариотической клетки Современные представления о возникновении жизни на Земле
Современные представления о возникновении жизни на Земле Эволюционная теория
Эволюционная теория Адам миының құрылысы және қызметі
Адам миының құрылысы және қызметі Закономерности роста и развития различных периодов онтогенеза
Закономерности роста и развития различных периодов онтогенеза