- Клеточный цикл. Апоптоз. Раковая трансформация клеток. Хромосомы

Содержание
- 2. Клеточный цикл и его регуляция
- 3. Митотический цикл клетки (клеточный цикл) – это совокупность процессов, происходящих в клетке от одного ее деления
- 4. Пример перехода клетки из G0 периода к делению – реакция бласттрансформации лимфоцитов. Была описана в 1902
- 5. Отрывок взят из: http://www.spletnik.ru/blogs/kruto/87576_emigratciya-umov Чтобы по-настоящему оценить масштаб возможностей, упущенных Россией, нужно вспомнить о тех, кто
- 6. Вернемся к митотическому циклу. Во время митоза хромосомы хорошо видны, но события интерфазы долго оставались загадкой.
- 7. Исследование митотического цикла с помощью меченого тритием тимидина (метод авторадиографии) Метод авторадиографии: В культуру делящихся клеток
- 8. Фазы митоза Профаза – хромосомы начинают конденсацию, ядрышки и мембрана ядра исчезают, центриоли расходятся к полюсам
- 9. Нарушения митоза Патология митоза, связанная с повреждением хромосом. Например, фрагментация хромосом. Слияние фрагментов может приводить к
- 10. В зависимости от митотической активности ткани делят на: Стабильные ткани — это ткани, в которых клетки
- 11. S CHECKPOINT Регуляция митотического цикла. Для клеток чрезвычайно важно знать – надо делиться или нет… Существуют
- 12. Белки циклины и циклин-зависимые киназы (англ. cyclin-dependent kinases, CDK) регулируют ход митотического цикла Всего у млекопитающих
- 13. Раскрытие тайн регуляции клеточного цикла. В 2001 году Нобелевской премии в области физиологии и медицины удостоились
- 14. Контрольные точки прохождения клеткой митотического цикла СDK присутствует в клетке постоянно, меняется концентрация циклинов. CDK4/6+cyclin D
- 15. Белок p53 является одним из факторов транскрипции, который инициирует синтез белка p21, являющегося ингибитором комплекса CDK-циклин
- 16. Нарушение регуляции клеточного цикла может вести к появлению нежизнеспособных клеток или к раковому перерождению. А. В
- 17. Апоптоз
- 18. Апоптоз – (др.-греч. ἀπόπτωσις — опадание листьев) это запрограммированная клеточная гибель (в отличие от некроза)
- 19. Гибель клетки путем апоптоза Фермент ДНК протеинкиназа узнает двухцепочечные разрывы в ДНК и активирует белок р53.
- 20. Апоптоз очень важен в многоклеточном организме. Первые доказательства наличия генетической программы клеточной смерти были получены при
- 21. Формирование пальцев (пример апоптоза). Гистологический срез лапы эмбриона мыши (Mus musculus), сделанное на 15-й день развития
- 22. Обобщённая схема апоптоза млекопитающих Не для запоминания!
- 23. Рак
- 24. У всех раковых клеток есть два общих признака: Неконтролируемый рост вследствие нарушения контроля клеточного цикла. Способность
- 25. Номенклатура онкогенов и супрессоров опухолей Онкогены записывают трёхзначным кодом из строчных латинских букв, который обычно указывает
- 26. Пример 1. Развитие ретинобластомы – опухоли сетчатки глаза связано с мутациями в гене Rb, супрессоре опухоли.
- 27. типах опухолей эта цифра доходит до 90%. Ras — это семейство генов и кодируемых ими белков,
- 28. Пример 3. Хронический миелоидный лейкоз (рак крови) связан с мутацией протонкогена с-alb Хромосомная мутация – транслокация
- 29. Клеточные сигналы, ответственные за апоптоз, деление клетки и регуляцию генов
- 30. Хроматин. Хромосомы. Цитогенетика.
- 31. Хроматин (греч. chroma — цвет, краска и греч. nitos — нить). Это комплекс ДНК с белками.
- 32. Хромосомная теория наследственности Гены лежат в хромосомах в линейном порядке Каждый ген занимает определенное место –
- 33. ДНК бактерий тоже иногда называется бактериальной хромосомой. Она кольцевая и лишена гистонов. Хотя белки бактериальной хромосомы
- 34. Хромосома (греч. – «окрашенное тело») – комплекс ДНК с белками (гистоновыми и негистоновыми) Строение хромосом эукариот
- 35. Уровни организации эукариотической хромосомы Уровни организации хромосом эукариот
- 36. ДНК – 2 нм Нуклеосома – 11 нм Фибрилла 30 нм «Соленоид» - 300 нм Нить
- 37. Гистоны играют важную роль как в упаковке, так и регуляции активности хроматина. Гистоны – небольшие глобулярные
- 38. Нуклеосома – низший уровень конденсации хроматина ДНК Кор из 8 молекул гистонов 2 х (Н2а, Н2b,
- 39. белковый каркас (остов, скаффолд) Нуклеосомы (11 нм) 30 нм Укладка типа «соленоида» 300 нм Нуклеосомы плотно
- 40. И, наконец, образуются максимально конденсированные метафазные хромосомы
- 41. Хромосомы к клетке в зависимости от фазы клеточного цикла бывают: Интерфазные, активные Митотические, неактивные интерфаза митоз
- 42. Интерфазные хромосомы – слабо упакованы и готовы к использованию (репликации, транскрипции и др.) Вид интерфазного ядра
- 43. Митотические хромосомы – подобны упакованным для переезда вещам Метафазная хромосома видна в микроскоп и неактивна
- 44. Рассмотрим интерфазные хромосомы Интерфазная хромосома
- 45. Интерфазные хромосомы – слабо упакованы и готовы к использованию (репликации, транскрипции и др.) Вид интерфазного ядра
- 46. В интерфазе хромосомы расположены в ядре неслучайным образом Хромосомные территории в интерфазном ядре фибробласта человека. FISH
- 47. Хроматин – комплекс ДНК и белков (гистонов и не гистонов) Хроматин Эухроматин (слабо конденсированный, активный) Гетерохроматин
- 48. Конститутивный гетерохроматин не содержит генов и сосредоточен в области центромеры и теломеров Центромера - это структура,
- 49. Центромерный район «Центромерная ДНК и ДНK прицентромерного гетерохроматина состоит из альфоидной (альфа-сателлитной) ДНК и ДНК, представляющей
- 50. Строение центромеры
- 51. Строение теломеры Теломеры представляют собой в основном двунитевые некодирующие повторы (ТТАГГГ)n, заканчивающиеся 3’-однонитевым участком Размер двунитевого
- 52. Тельце Барра – пример факультативного гетерохроматина. Его можно видеть в соматических клетках самок млекопитающих ФАКУЛЬТАТИВНЫЙ ГЕТЕРОХРОМАТИН
- 53. В выключении Х-хромосомы у самок млекопитающих участвует нкРНК Канадский ученый Барр (1908 – 1995) (и его
- 54. Инактивация Х-хромосомы у самок млекопитающих В части клеток активна Х от отца, в части от матери
- 55. Xist РНК окружает ту Х хромосому, с которой экспрессируется, и подавляет ее почти всю Инактивация –
- 56. Исследование Х-полового хроматина (тельца Барра). У пациента берется соскоб эпителия ротовой полости Помещается на предметное стекло
- 57. Теперь поговорим о митотических хромосомах Митотическая хромосома
- 58. Изучение митотических хромосом.
- 59. Краткая история цитогенетики «запоздалое, но счастливое рождение» До 50-х годов ХХ века хромосомы человека исследовались, но
- 60. Определение числа хромосом человека (1956 г. ) J-H. Tjio (1919–2001) A. Levan (1905–1998).
- 61. Т. Касперсон и другие – методы дифференциальной окраски хромосом (1968 – 70гг.) G-окраска Q-окраска
- 62. FISH- этап – внедрение и широкое использование молекулярно-цитогенетических методов (с 1986 г.) 1986 г . -
- 63. Существуют разные варианты FISH метода как для метафазных, так и для интерфазных хромосом.
- 64. Отечественные цитогенетики Г.А. Левитский (1878-1942) - ввел термин «кариотип» в современном его понимании. Автор одного из
- 65. «Если бы эти лаборатории в СССР продолжали работать, то большинство открытий по кариотипу человека, сделанных в
- 66. Этапы цитогенетического исследования Этапы цитогенетического исследования
- 67. Кровь (или другой материал: костный мозг, околоплодная жидкость и др.) Отделение лейкоцитов Добавление стимулятора митоза –
- 68. Добавление гипотонического раствора – клетки разбухают. Фиксация. Х ХХ х х при раскапывании от удара о
- 69. На стекле хромосомы образуют метафазные пластинки Сплошное (рутинное) окрашивание
- 70. Виды окраски хромосом Рутинная, появилась в 50-х годах ХХ века. (Денверская классификация поделила все хромосомы человека
- 71. G –окраска, самая распространненная окраска, выявляются полосы - бэнды
- 72. Q –окраска выявляет те же бэнды, что и G окраска.
- 73. C –окраска выявляет конститутивный гетерохроматин
- 74. Окраска азотнокислым серебром выявляет ядрышко-образующие районы хромосом
- 75. Ядрышко-образующие районы хромосом Ядрышко-образующие районы хромосом (ЯОР) локализованы в коротких плечах акроцентрических хромосом человека 13, 14,
- 76. FISH -метод – Fluorescent in situ hybridization, используются разноцветные красители, а затем компьютер присваивает хромосомам условные
- 77. Схема FISH-метода Флуоресцентная метка ДНК-зонд Участок хромосомы, комплементарный зонду Метафазные хромосомы с меткой, здесь использовали 2
- 78. Может использоваться как для делящихся, так и неделящихся клеток
- 79. FISH-метод позволяет лучше распознавать хромосомные перестройки, чем одноцветная окраска
- 80. Классификация (номенклатура) хромосом
- 81. Виды метафазных хромосом согласно Денверской классификации Метацентрическая, субметацентрическая, акроцентрическая, телоцентрическая*, со спутником *По современным представлениям телоцентрических
- 82. Хромосомы человека по Денверской классификации делят на 7 групп
- 83. Парижская классификация дополняет Денверскую и основана на дифференциальной окраске (чаще всего G-окраска)
- 84. 300 бэндов 550 бэндов 700-бэндов На слабо конденсированных (прометафазных) хромосомах можно видеть больше бэндов
- 85. Плечи делят на районы (бэнды) и суббэнды Нумерация бэндов идет от центромеры к теломерам
- 86. первая цифра - номер хромосомы, в которой локализован данный бэнд; второй символ (p или q) -
- 87. На Парижской конференции была принята единая запись хромосомных аберраций Фрагмент f + + теряется rob r
- 88. Метафазная пластинка с кольцевой хромосомой, парными ацентрическими фрагментами и дицентрическими хромосомами.
- 89. Трисомия записывается знаком «+»
- 91. Примеры записи хромосомного диагноза 46,XX обычная женщина 46,XY обычный мужчина 69,XXY Мужской триплоидный кариотип 46,XX,del(14)(q23) Женщина
- 92. V Международный конгресс по генетике человека (Мехико, 1972г.) оформил официальную номенклатуру хромосом человека - «An International
- 93. Необычные хромосомы
- 94. Необычные виды хромосом «ламповые щетки». Впервые хромосомы типа ламповых щёток были описаны В. Флеммингом в 1882
- 95. Хромосомы типа ламповых щеток в овоцитах амфибий Хромосомы типа ламповых щёток — это гигантская форма хромосом,
- 96. Схема строения «ламповой щетки»
- 97. Политенные (многонитчатые) хромосомы личинок двукрылых Репликация ДНК не сопровождается делением клетки, что приводит к накоплению вновь
- 98. Активные участки -пуфы Неактивные - диски Политенные хромосомы дрозофилы
- 99. Хромосомные карты
- 100. Хромосомные карты делят на Генетические – показывают, где лежит какой ген Цитологические – по окраске Физические
- 101. Основные методы составления генетических (хромосомных) карт На основе скрещиваний - не у человека! (гибридологический метод) -
- 102. Расстояние генов В и V – 17 морганид Первым организмом, для которого была получена генетическая карта,
- 103. Родословная, показывающая сцепление у человека Был кроссинговер Был кроссинговер Было проанализировано много семей. По частоте кроссинговера
- 104. Метод изучения генетики соматических клеток Клетки выращивают в культуре. Этим методом удалось картировать гены человека. Метод
- 105. В ходе клеточных делений в гибридной клетке утрачиваются все хромосомы человека, кроме одной (например, № 17)
- 106. Картирование FISH-методом c использованием ДНК - зондов Флуоресцентная метка ДНК-зонд Участок хромосомы, комплементарный зонду Метафазные хромосомы
- 107. Карта хромосомы 9 – пример комбинированной карты
- 109. Скачать презентацию

Клеточный цикл и его регуляция
Клеточный цикл и его регуляция
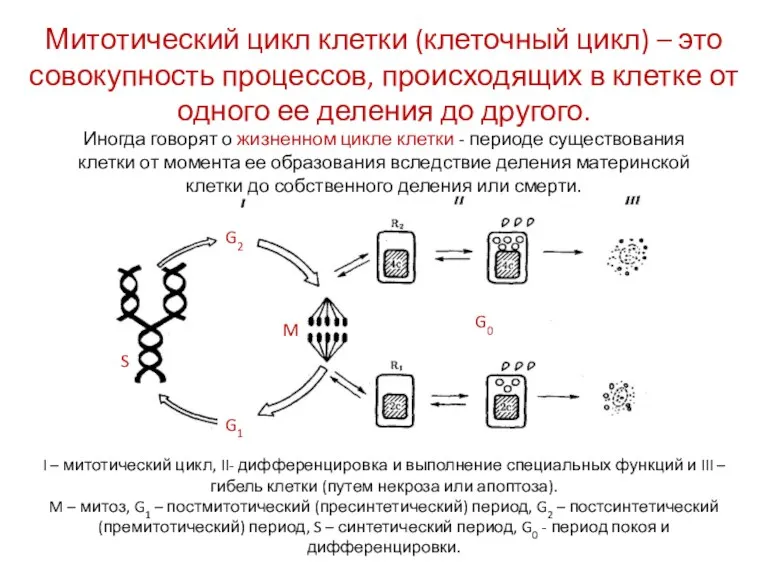
Митотический цикл клетки (клеточный цикл) – это совокупность процессов, происходящих в
Митотический цикл клетки (клеточный цикл) – это совокупность процессов, происходящих в
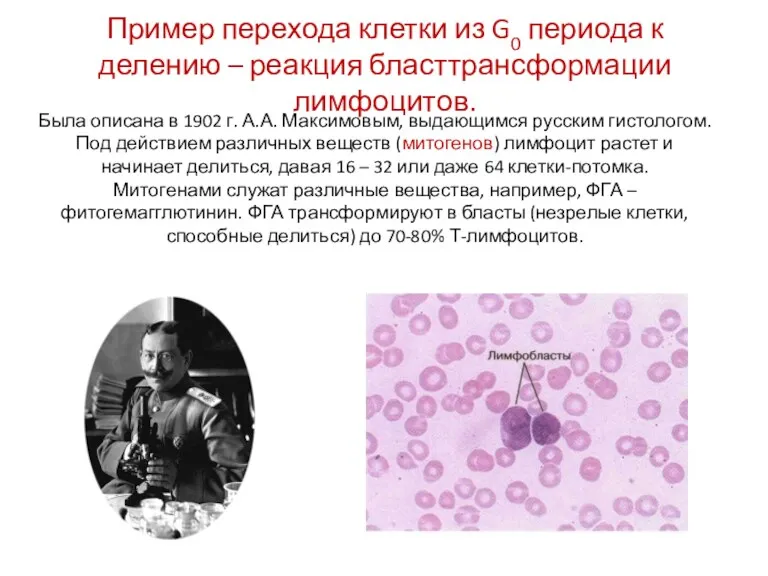
Пример перехода клетки из G0 периода к делению – реакция бласттрансформации
Пример перехода клетки из G0 периода к делению – реакция бласттрансформации

Отрывок взят из: http://www.spletnik.ru/blogs/kruto/87576_emigratciya-umov
Чтобы по-настоящему оценить масштаб возможностей, упущенных Россией, нужно
Отрывок взят из: http://www.spletnik.ru/blogs/kruto/87576_emigratciya-umov
Чтобы по-настоящему оценить масштаб возможностей, упущенных Россией, нужно
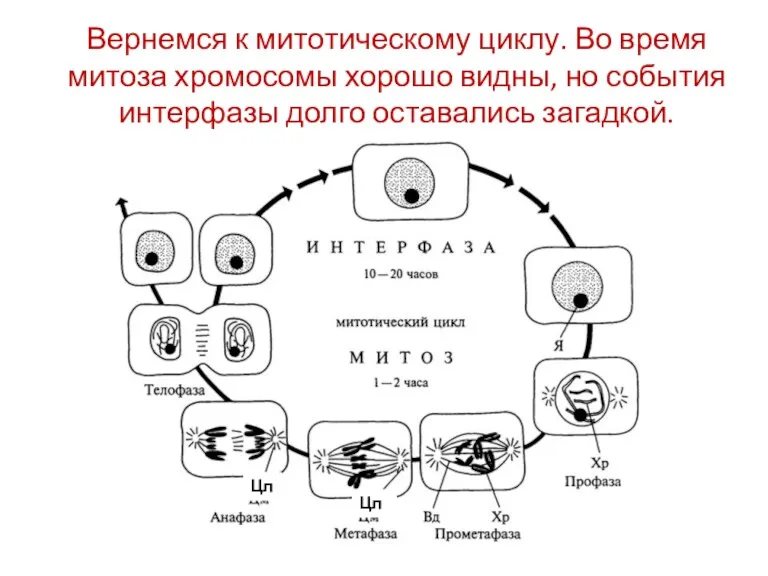
Вернемся к митотическому циклу. Во время митоза хромосомы хорошо видны, но
Вернемся к митотическому циклу. Во время митоза хромосомы хорошо видны, но
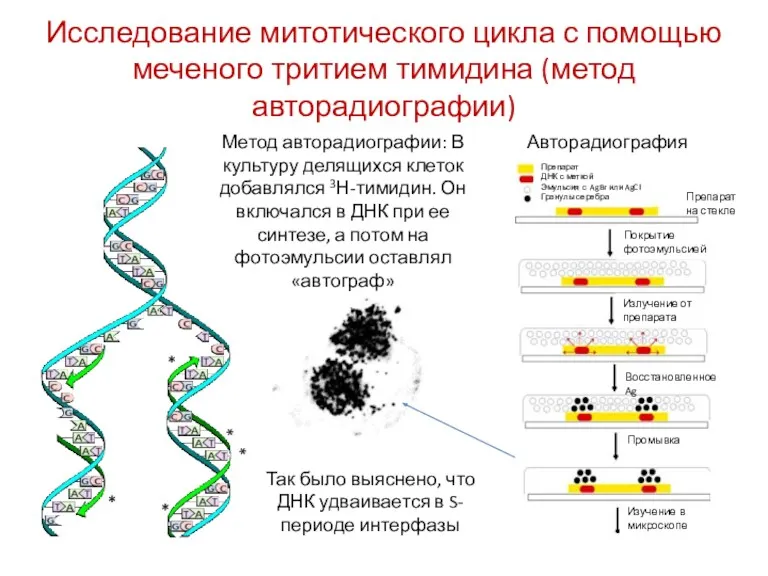
Исследование митотического цикла с помощью меченого тритием тимидина (метод авторадиографии)
Метод авторадиографии:
Исследование митотического цикла с помощью меченого тритием тимидина (метод авторадиографии)
Метод авторадиографии:
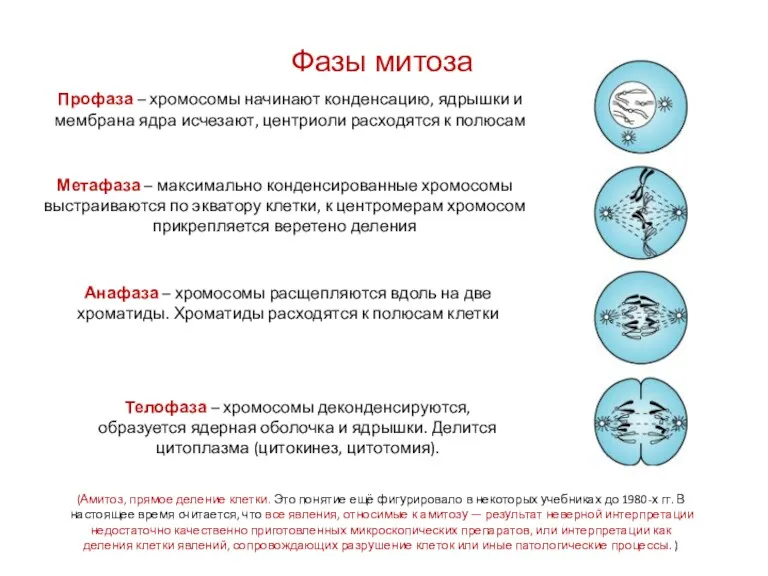
Фазы митоза
Профаза – хромосомы начинают конденсацию, ядрышки и мембрана ядра исчезают,
Фазы митоза
Профаза – хромосомы начинают конденсацию, ядрышки и мембрана ядра исчезают,
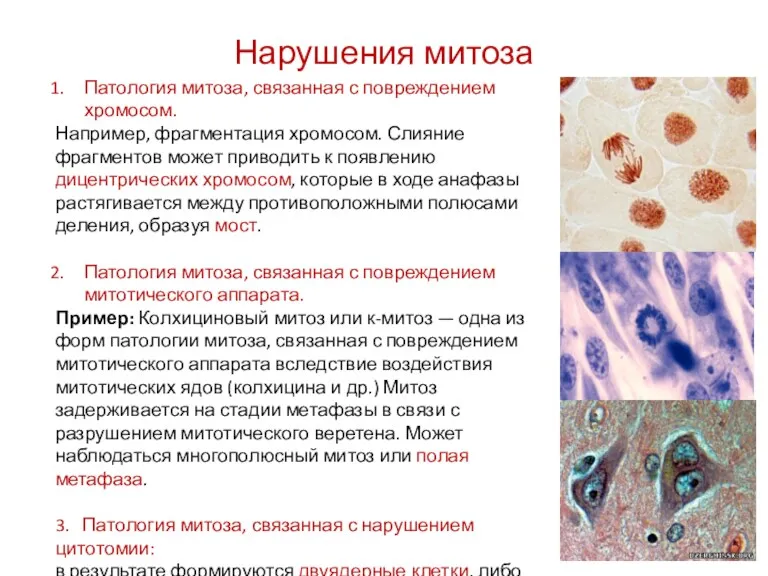
Нарушения митоза
Патология митоза, связанная с повреждением хромосом.
Например, фрагментация хромосом. Слияние
Нарушения митоза
Патология митоза, связанная с повреждением хромосом.
Например, фрагментация хромосом. Слияние
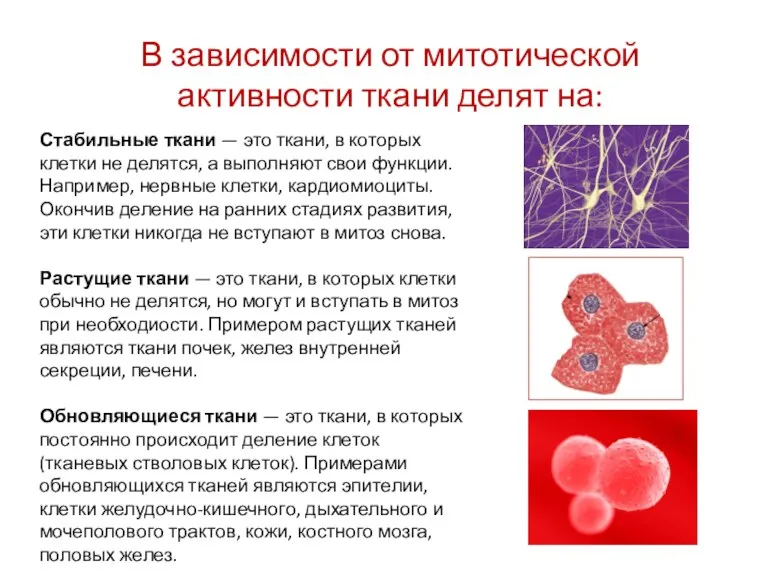
В зависимости от митотической активности ткани делят на:
Стабильные ткани — это
В зависимости от митотической активности ткани делят на:
Стабильные ткани — это
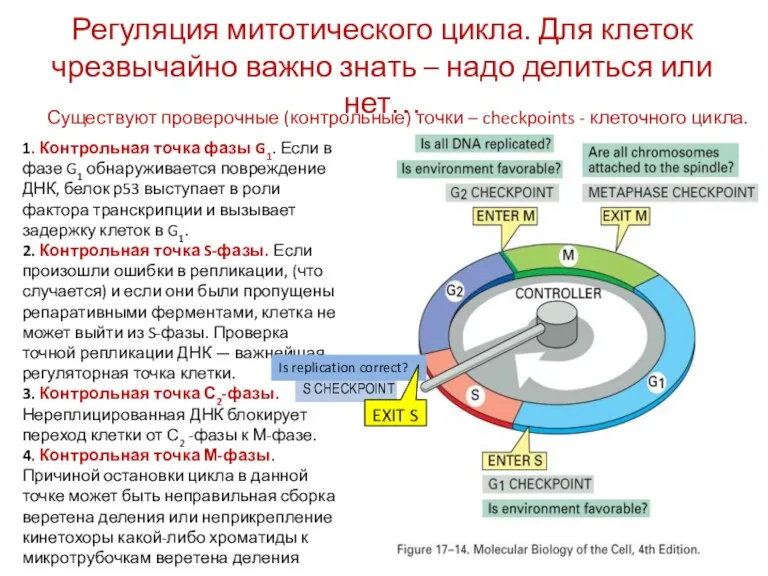
S CHECKPOINT
Регуляция митотического цикла. Для клеток чрезвычайно важно знать – надо
S CHECKPOINT
Регуляция митотического цикла. Для клеток чрезвычайно важно знать – надо
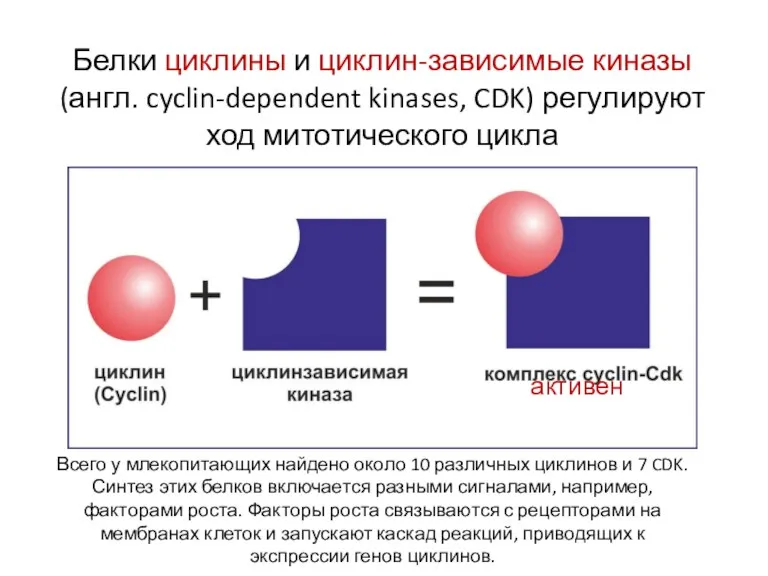
Белки циклины и циклин-зависимые киназы (англ. cyclin-dependent kinases, CDK) регулируют ход
Белки циклины и циклин-зависимые киназы (англ. cyclin-dependent kinases, CDK) регулируют ход

Раскрытие тайн регуляции клеточного цикла.
В 2001 году Нобелевской премии
Раскрытие тайн регуляции клеточного цикла.
В 2001 году Нобелевской премии
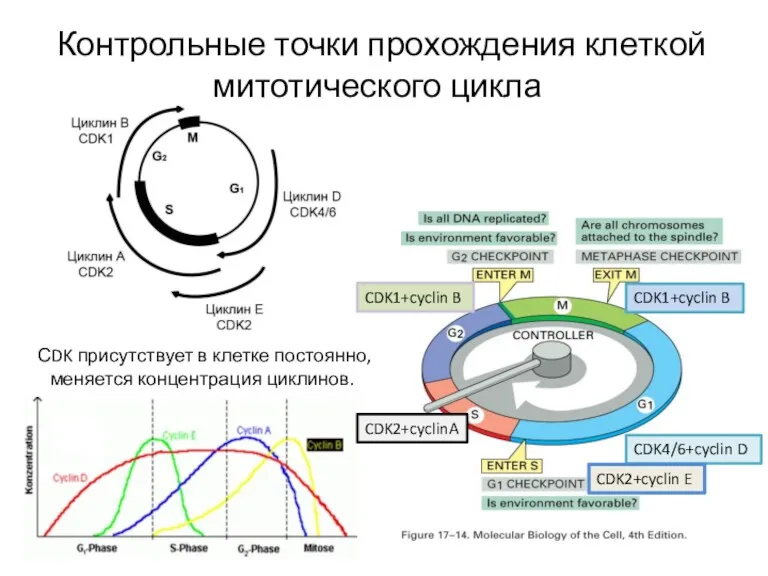
Контрольные точки прохождения клеткой митотического цикла
СDK присутствует в клетке постоянно, меняется
Контрольные точки прохождения клеткой митотического цикла
СDK присутствует в клетке постоянно, меняется
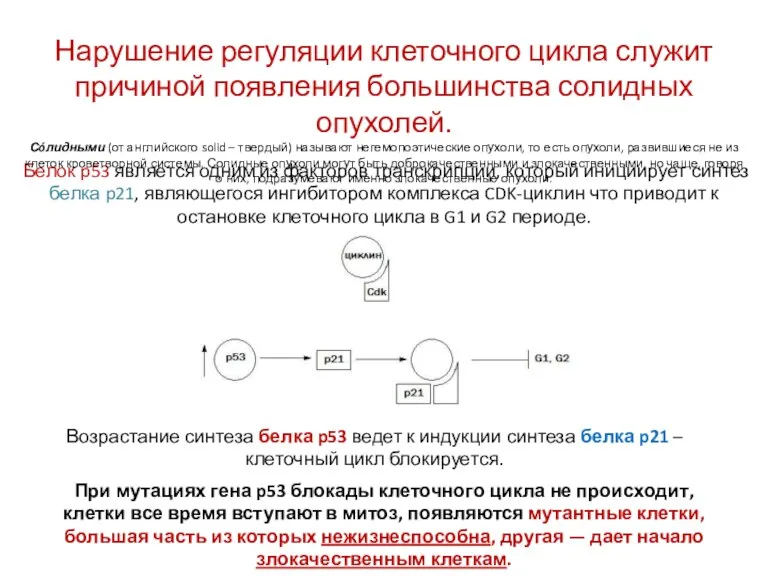
Белок p53 является одним из факторов транскрипции, который инициирует синтез белка
Белок p53 является одним из факторов транскрипции, который инициирует синтез белка
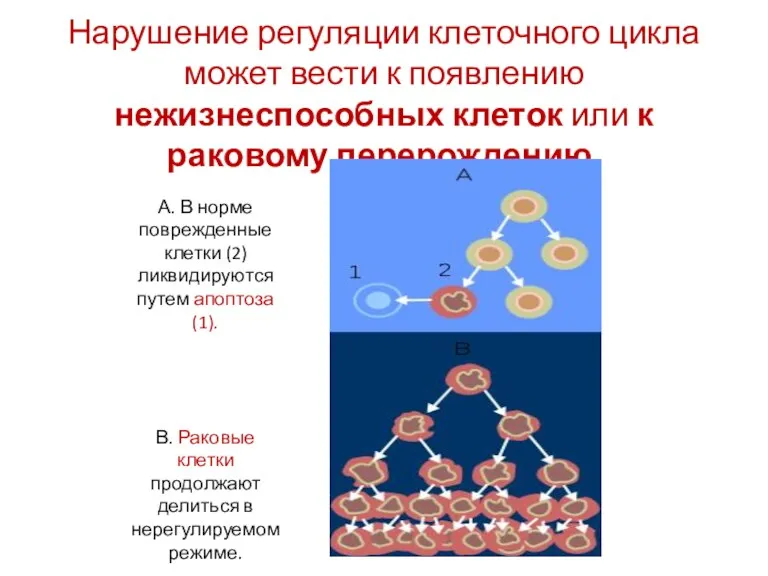
Нарушение регуляции клеточного цикла может вести к появлению нежизнеспособных клеток или
Нарушение регуляции клеточного цикла может вести к появлению нежизнеспособных клеток или

Апоптоз
Апоптоз
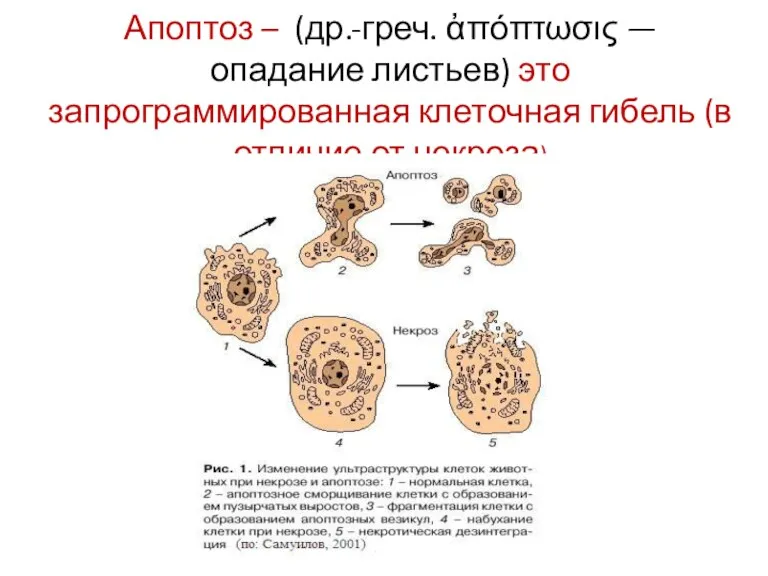
Апоптоз – (др.-греч. ἀπόπτωσις — опадание листьев) это запрограммированная клеточная гибель
Апоптоз – (др.-греч. ἀπόπτωσις — опадание листьев) это запрограммированная клеточная гибель
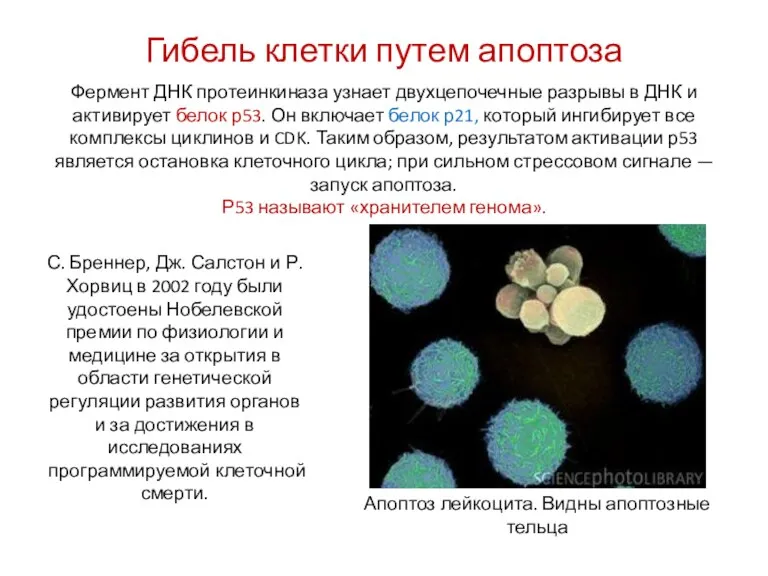
Гибель клетки путем апоптоза
Фермент ДНК протеинкиназа узнает двухцепочечные разрывы в ДНК
Гибель клетки путем апоптоза
Фермент ДНК протеинкиназа узнает двухцепочечные разрывы в ДНК

Апоптоз очень важен в многоклеточном организме.
Первые доказательства наличия генетической программы клеточной
Апоптоз очень важен в многоклеточном организме.
Первые доказательства наличия генетической программы клеточной
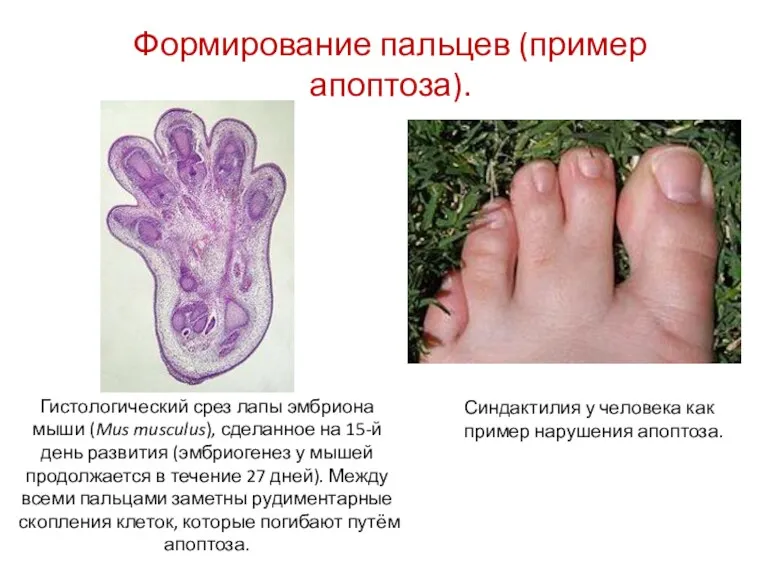
Формирование пальцев (пример апоптоза).
Гистологический срез лапы эмбриона мыши (Mus musculus), сделанное
Формирование пальцев (пример апоптоза).
Гистологический срез лапы эмбриона мыши (Mus musculus), сделанное
Обобщённая схема апоптоза млекопитающих
Не для запоминания!
Обобщённая схема апоптоза млекопитающих
Не для запоминания!

Рак
Рак

У всех раковых клеток есть два общих признака:
Неконтролируемый рост вследствие нарушения
У всех раковых клеток есть два общих признака:
Неконтролируемый рост вследствие нарушения

Номенклатура онкогенов и супрессоров опухолей
Онкогены записывают трёхзначным кодом из строчных латинских
Номенклатура онкогенов и супрессоров опухолей
Онкогены записывают трёхзначным кодом из строчных латинских
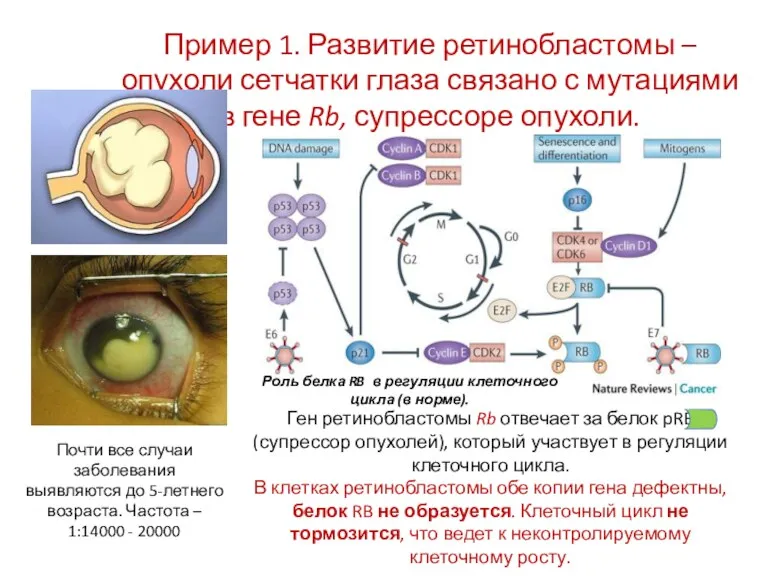
Пример 1. Развитие ретинобластомы – опухоли сетчатки глаза связано с мутациями
Пример 1. Развитие ретинобластомы – опухоли сетчатки глаза связано с мутациями
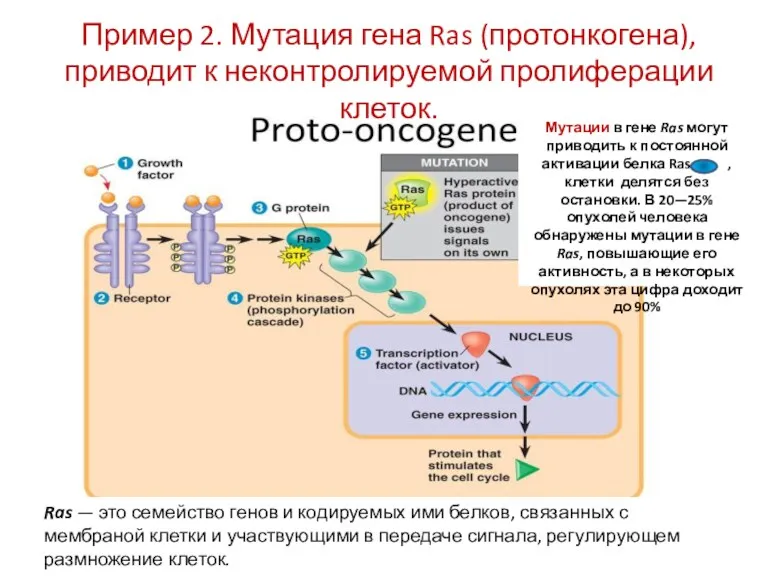
типах опухолей эта цифра доходит до 90%.
Ras — это семейство генов
типах опухолей эта цифра доходит до 90%.
Ras — это семейство генов
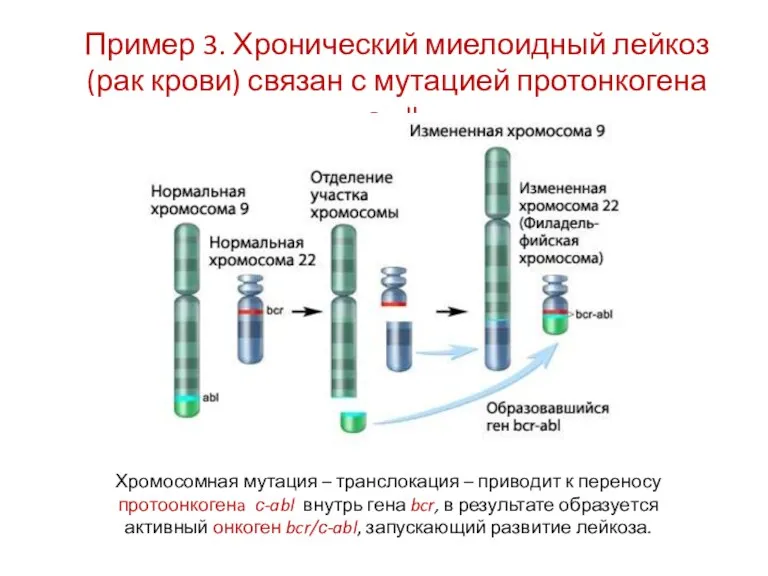
Пример 3. Хронический миелоидный лейкоз (рак крови) связан с мутацией протонкогена
Пример 3. Хронический миелоидный лейкоз (рак крови) связан с мутацией протонкогена
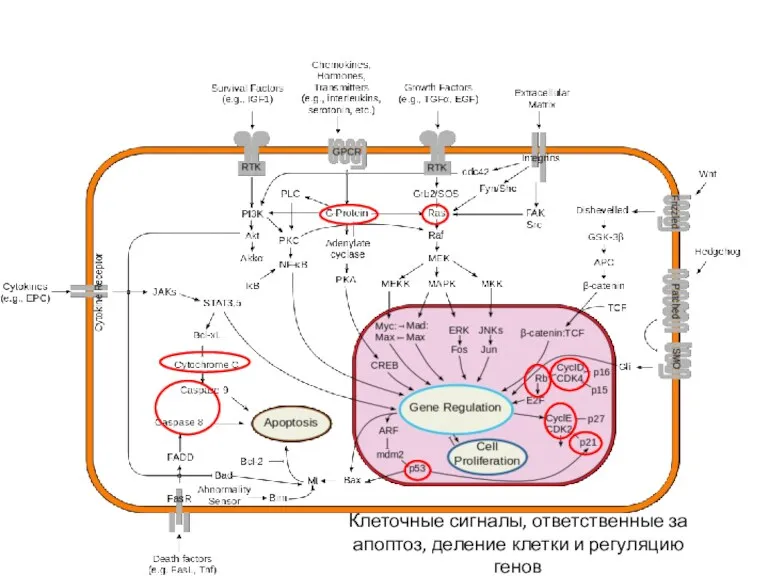
Клеточные сигналы, ответственные за апоптоз, деление клетки и регуляцию генов
Клеточные сигналы, ответственные за апоптоз, деление клетки и регуляцию генов

Хроматин. Хромосомы. Цитогенетика.
Хроматин. Хромосомы. Цитогенетика.

Хроматин (греч. chroma — цвет, краска и греч. nitos — нить). Это комплекс ДНК с белками.
Хромосома
Хроматин (греч. chroma — цвет, краска и греч. nitos — нить). Это комплекс ДНК с белками.
Хромосома

Хромосомная теория наследственности
Гены лежат в хромосомах в линейном порядке
Каждый ген
Хромосомная теория наследственности
Гены лежат в хромосомах в линейном порядке
Каждый ген
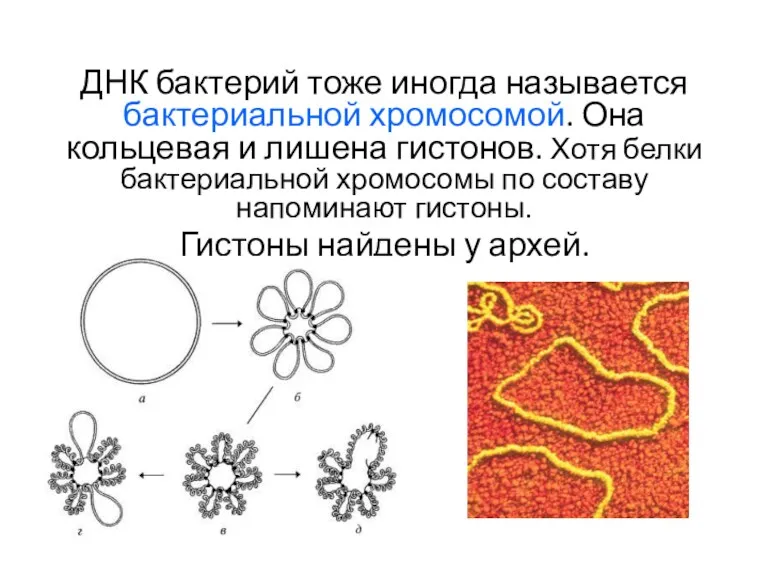
ДНК бактерий тоже иногда называется бактериальной хромосомой. Она кольцевая и лишена
ДНК бактерий тоже иногда называется бактериальной хромосомой. Она кольцевая и лишена
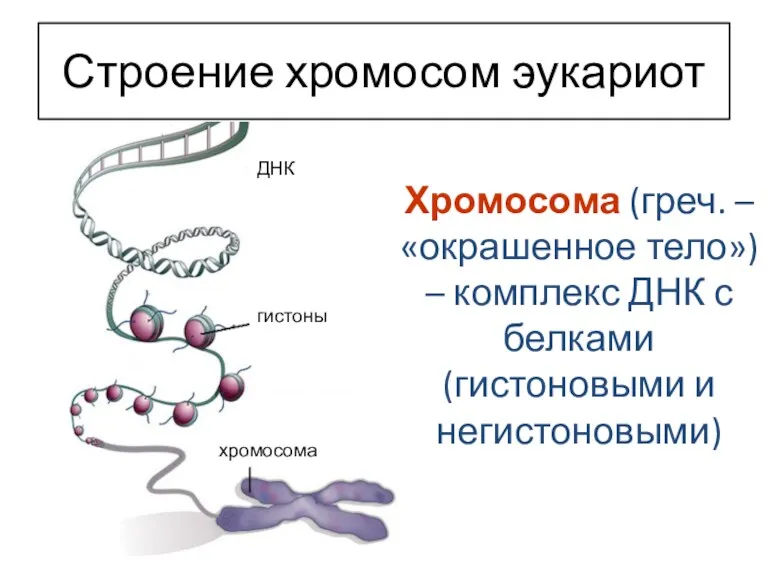
Хромосома (греч. – «окрашенное тело») – комплекс ДНК с белками (гистоновыми
Хромосома (греч. – «окрашенное тело») – комплекс ДНК с белками (гистоновыми

Уровни организации эукариотической хромосомы
Уровни организации хромосом эукариот
Уровни организации эукариотической хромосомы
Уровни организации хромосом эукариот
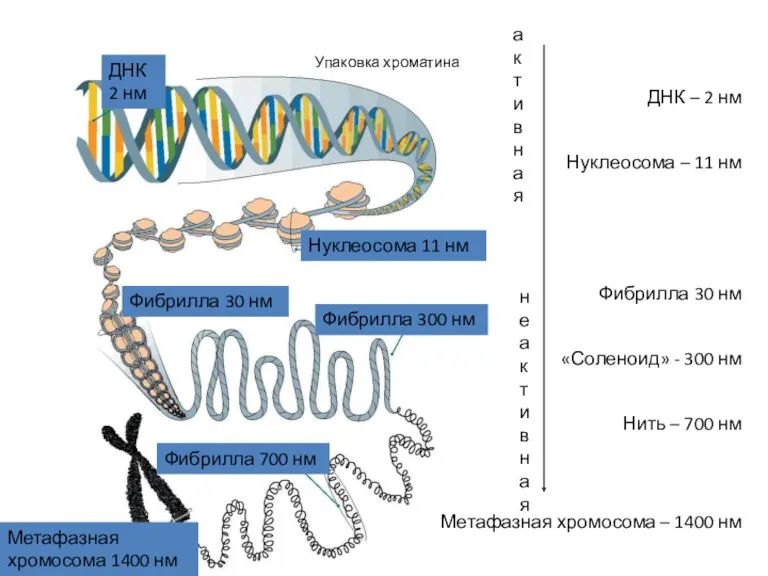
ДНК – 2 нм
Нуклеосома – 11 нм
Фибрилла 30 нм
«Соленоид» - 300
ДНК – 2 нм
Нуклеосома – 11 нм
Фибрилла 30 нм
«Соленоид» - 300
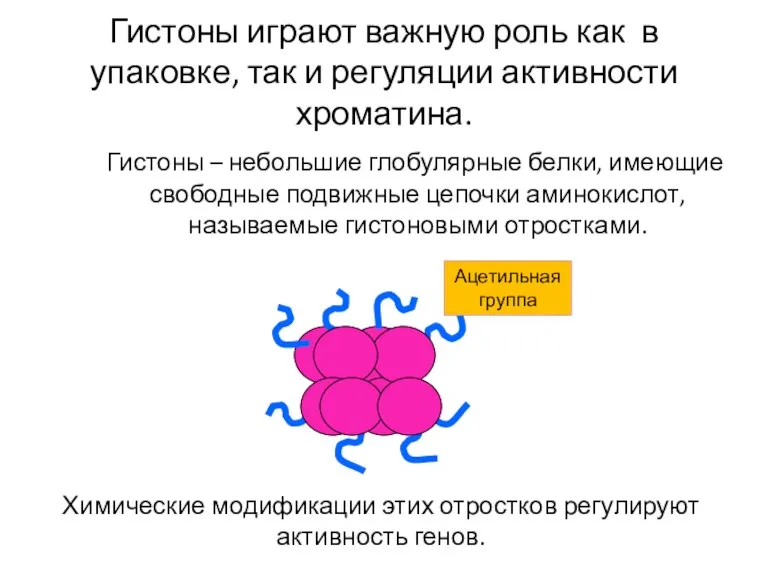
Гистоны играют важную роль как в упаковке, так и регуляции активности
Гистоны играют важную роль как в упаковке, так и регуляции активности
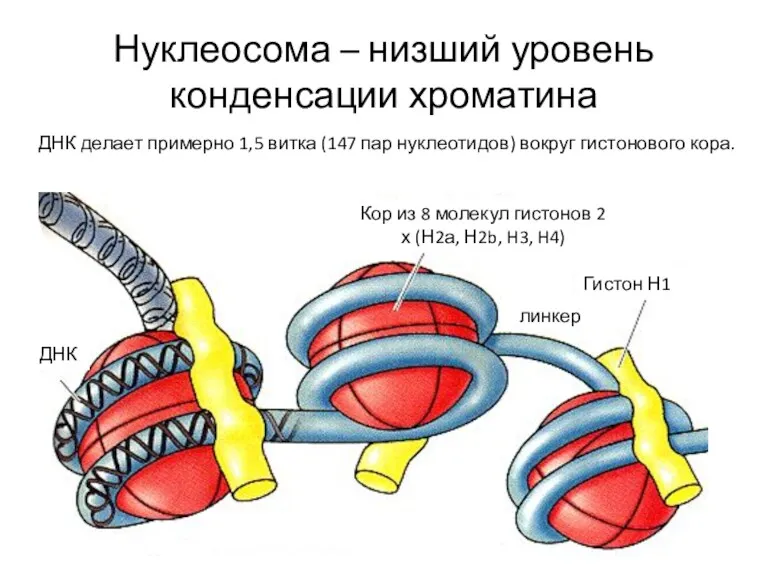
Нуклеосома – низший уровень конденсации хроматина
ДНК
Кор из 8 молекул гистонов 2
Нуклеосома – низший уровень конденсации хроматина
ДНК
Кор из 8 молекул гистонов 2
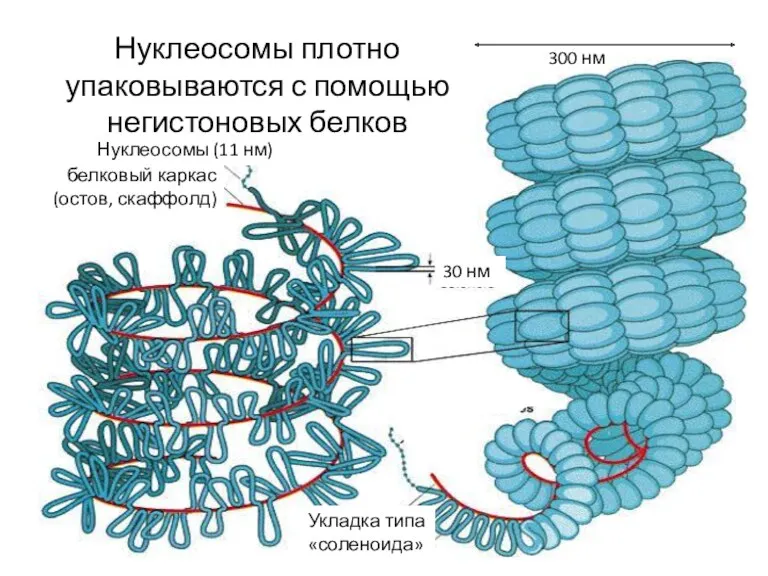
белковый каркас (остов, скаффолд)
Нуклеосомы (11 нм)
30 нм
Укладка типа «соленоида»
300 нм
Нуклеосомы плотно
белковый каркас (остов, скаффолд)
Нуклеосомы (11 нм)
30 нм
Укладка типа «соленоида»
300 нм
Нуклеосомы плотно
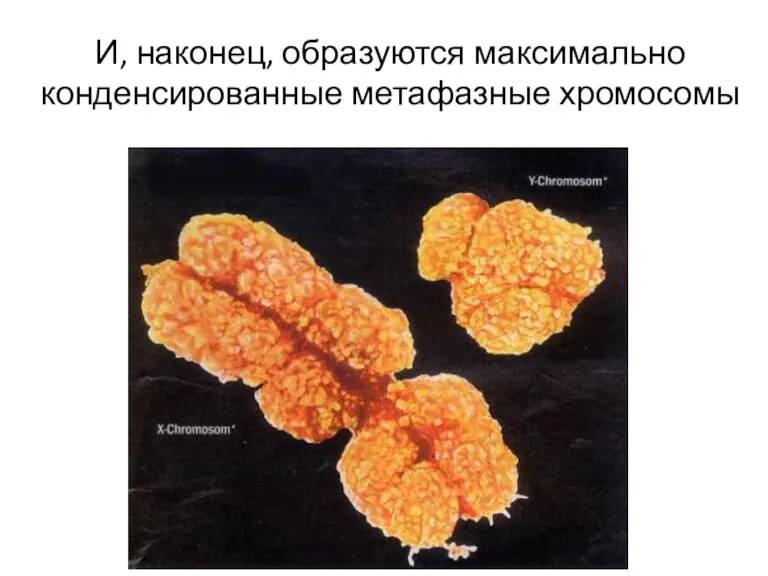
И, наконец, образуются максимально конденсированные метафазные хромосомы
И, наконец, образуются максимально конденсированные метафазные хромосомы
Хромосомы к клетке в зависимости от фазы клеточного цикла бывают:
Интерфазные,
Хромосомы к клетке в зависимости от фазы клеточного цикла бывают:
Интерфазные,
Интерфазные хромосомы – слабо упакованы и готовы к использованию (репликации, транскрипции
Интерфазные хромосомы – слабо упакованы и готовы к использованию (репликации, транскрипции

Митотические хромосомы – подобны упакованным для переезда вещам
Метафазная хромосома видна в
Митотические хромосомы – подобны упакованным для переезда вещам
Метафазная хромосома видна в

Рассмотрим интерфазные хромосомы
Интерфазная хромосома
Рассмотрим интерфазные хромосомы
Интерфазная хромосома
Интерфазные хромосомы – слабо упакованы и готовы к использованию (репликации, транскрипции
Интерфазные хромосомы – слабо упакованы и готовы к использованию (репликации, транскрипции
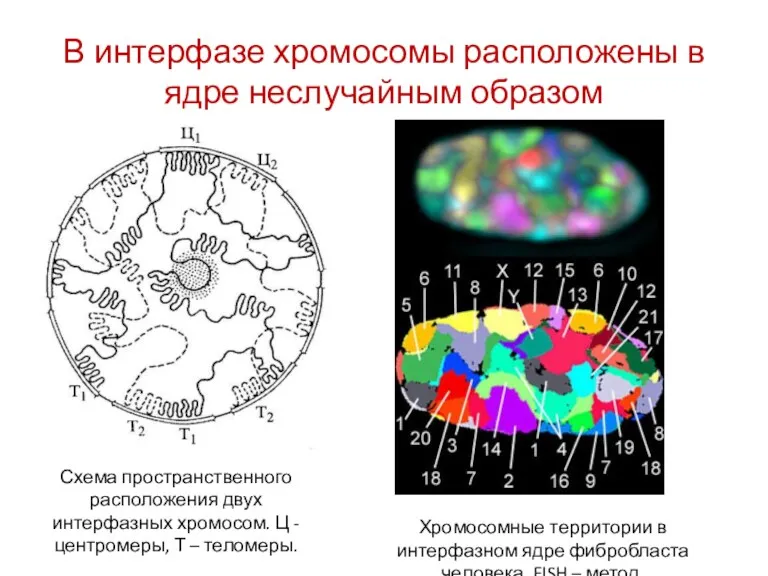
В интерфазе хромосомы расположены в ядре неслучайным образом
Хромосомные территории в интерфазном
В интерфазе хромосомы расположены в ядре неслучайным образом
Хромосомные территории в интерфазном
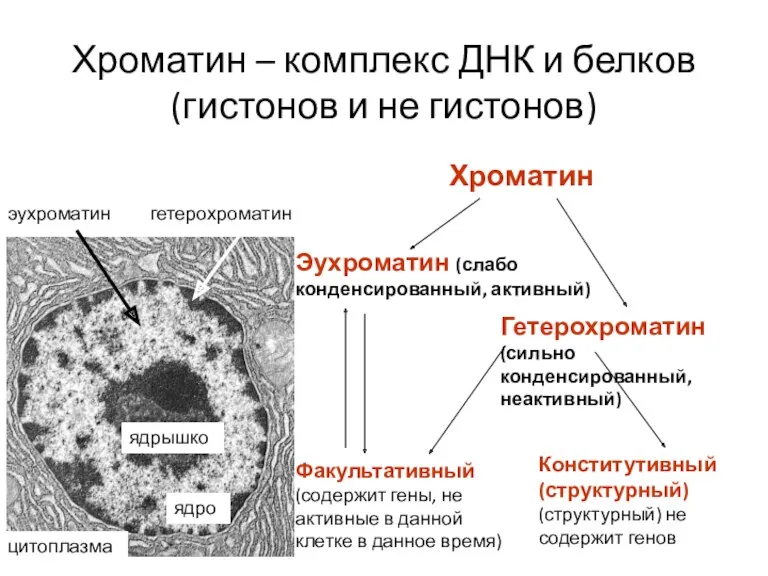
Хроматин – комплекс ДНК и белков (гистонов и не гистонов)
Хроматин
Эухроматин
Хроматин – комплекс ДНК и белков (гистонов и не гистонов)
Хроматин
Эухроматин

Конститутивный гетерохроматин не содержит генов и сосредоточен в области центромеры и
Конститутивный гетерохроматин не содержит генов и сосредоточен в области центромеры и
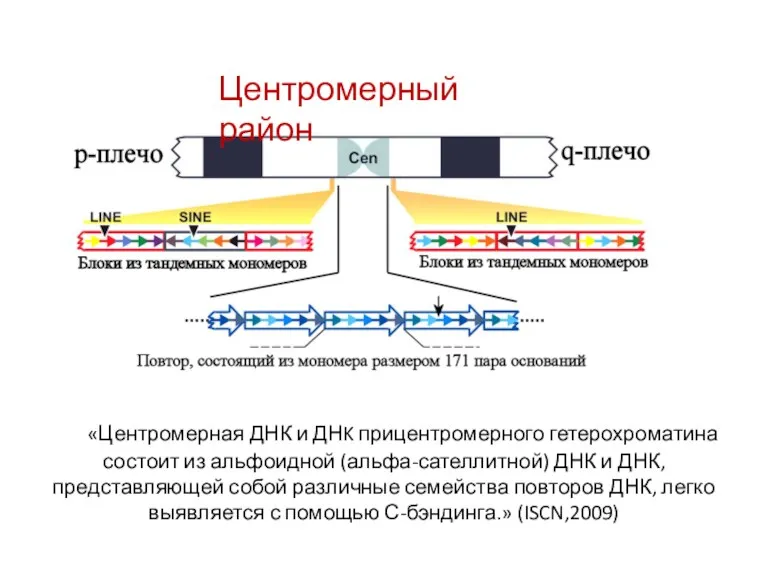
Центромерный район
«Центромерная ДНК и ДНK прицентромерного гетерохроматина состоит из альфоидной
Центромерный район
«Центромерная ДНК и ДНK прицентромерного гетерохроматина состоит из альфоидной
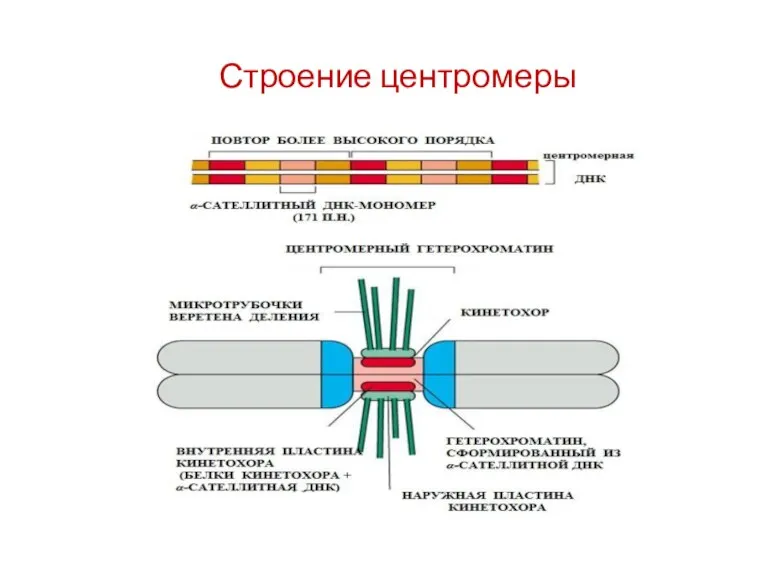
Строение центромеры
Строение центромеры
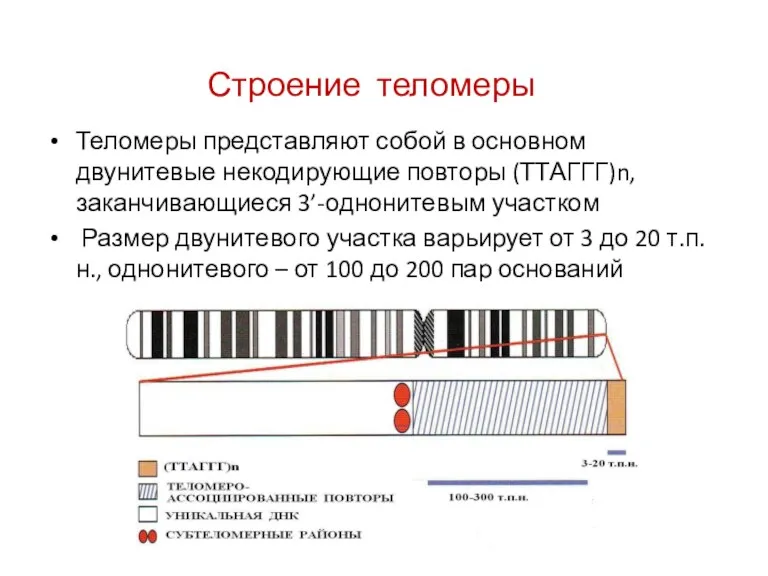
Строение теломеры
Теломеры представляют собой в основном двунитевые некодирующие повторы (ТТАГГГ)n, заканчивающиеся
Строение теломеры
Теломеры представляют собой в основном двунитевые некодирующие повторы (ТТАГГГ)n, заканчивающиеся
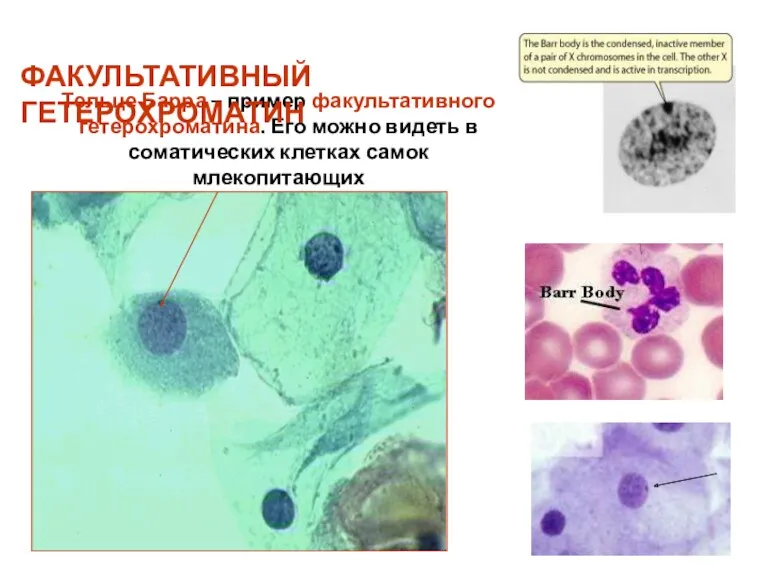
Тельце Барра – пример факультативного гетерохроматина. Его можно видеть в соматических
Тельце Барра – пример факультативного гетерохроматина. Его можно видеть в соматических
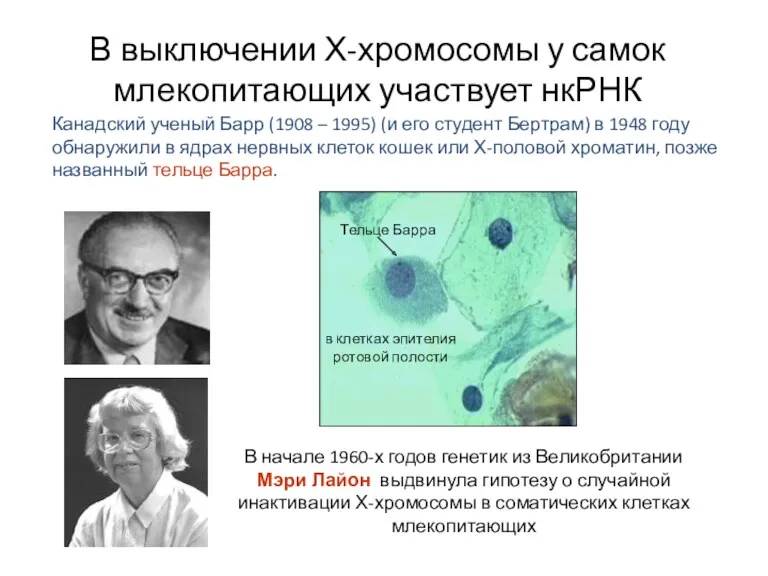
В выключении Х-хромосомы у самок млекопитающих участвует нкРНК
Канадский ученый Барр (1908
В выключении Х-хромосомы у самок млекопитающих участвует нкРНК
Канадский ученый Барр (1908
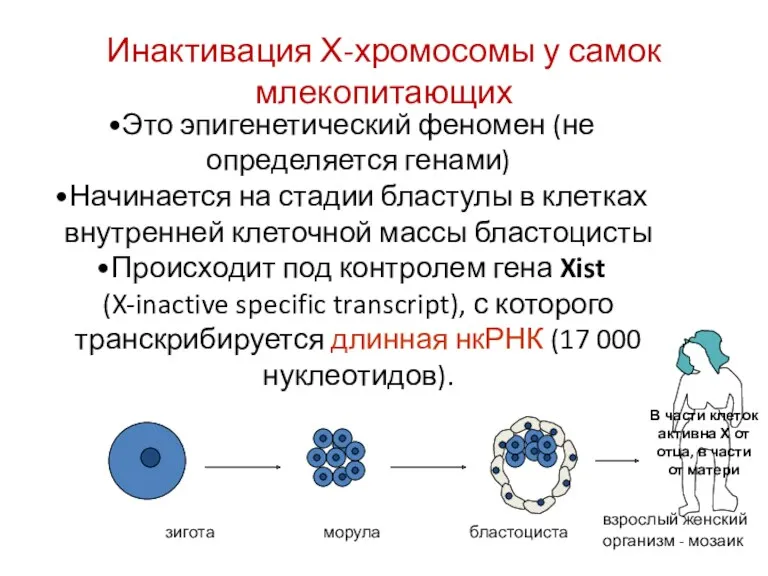
Инактивация Х-хромосомы у самок млекопитающих
В части клеток активна Х от отца,
Инактивация Х-хромосомы у самок млекопитающих
В части клеток активна Х от отца,
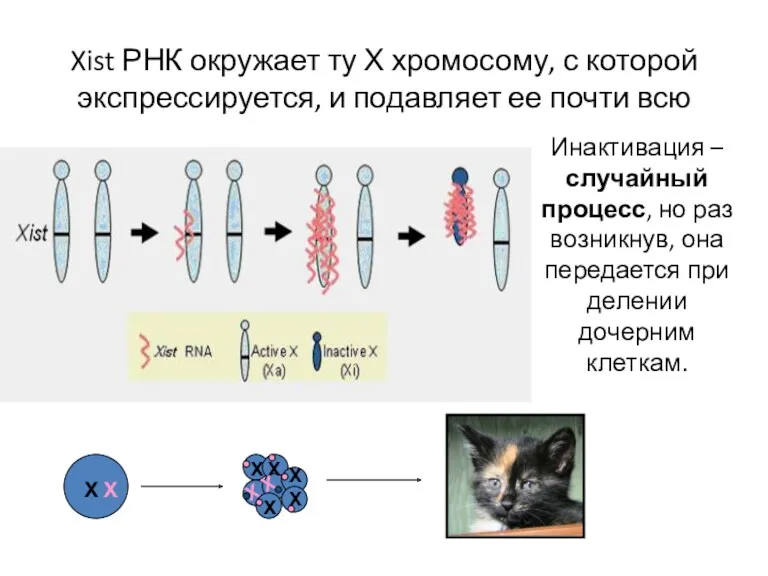
Xist РНК окружает ту Х хромосому, с которой экспрессируется, и подавляет
Xist РНК окружает ту Х хромосому, с которой экспрессируется, и подавляет
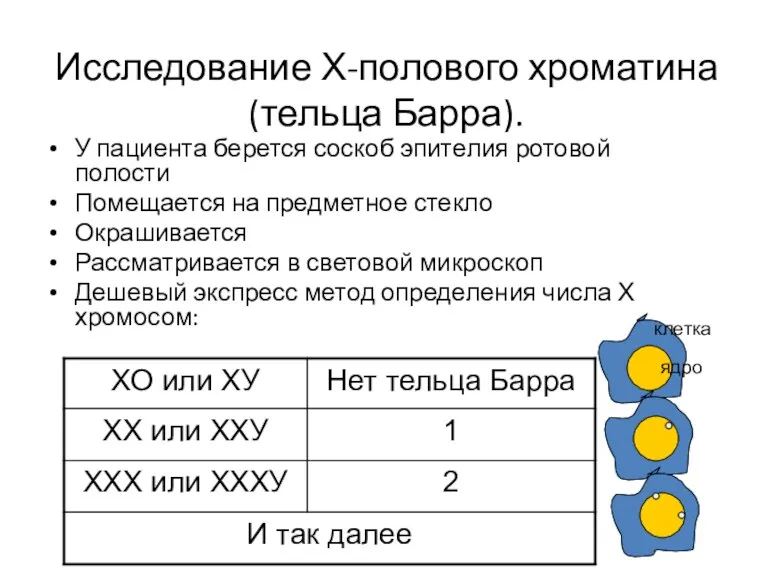
Исследование Х-полового хроматина (тельца Барра).
У пациента берется соскоб эпителия ротовой полости
Помещается
Исследование Х-полового хроматина (тельца Барра).
У пациента берется соскоб эпителия ротовой полости
Помещается

Теперь поговорим о митотических хромосомах
Митотическая хромосома
Теперь поговорим о митотических хромосомах
Митотическая хромосома

Изучение митотических хромосом.
Изучение митотических хромосом.

Краткая история цитогенетики
«запоздалое, но счастливое рождение»
До 50-х годов ХХ века хромосомы
Краткая история цитогенетики
«запоздалое, но счастливое рождение»
До 50-х годов ХХ века хромосомы

Определение числа хромосом человека (1956 г. )
J-H. Tjio (1919–2001)
A. Levan (1905–1998).
Определение числа хромосом человека (1956 г. )
J-H. Tjio (1919–2001)
A. Levan (1905–1998).
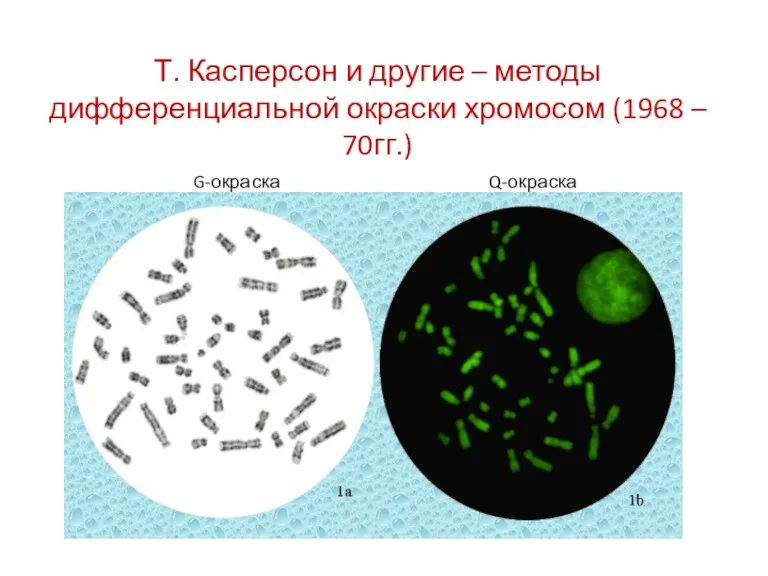
Т. Касперсон и другие – методы дифференциальной окраски хромосом (1968 –
Т. Касперсон и другие – методы дифференциальной окраски хромосом (1968 –

FISH- этап – внедрение и широкое использование молекулярно-цитогенетических методов (с 1986
FISH- этап – внедрение и широкое использование молекулярно-цитогенетических методов (с 1986
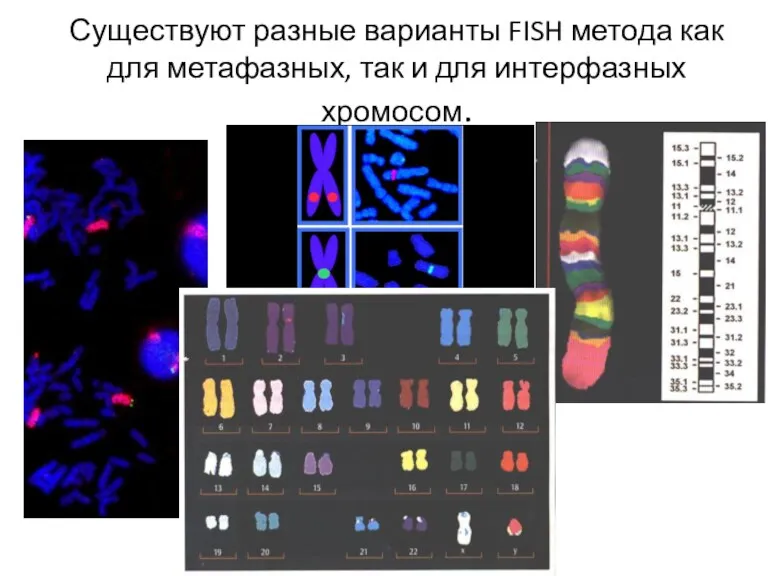
Существуют разные варианты FISH метода как для метафазных, так и для
Существуют разные варианты FISH метода как для метафазных, так и для

Отечественные цитогенетики
Г.А. Левитский (1878-1942) - ввел термин «кариотип» в современном
Отечественные цитогенетики
Г.А. Левитский (1878-1942) - ввел термин «кариотип» в современном

«Если бы эти лаборатории в СССР продолжали работать, то большинство открытий
«Если бы эти лаборатории в СССР продолжали работать, то большинство открытий

Этапы цитогенетического исследования
Этапы цитогенетического исследования
Этапы цитогенетического исследования
Этапы цитогенетического исследования
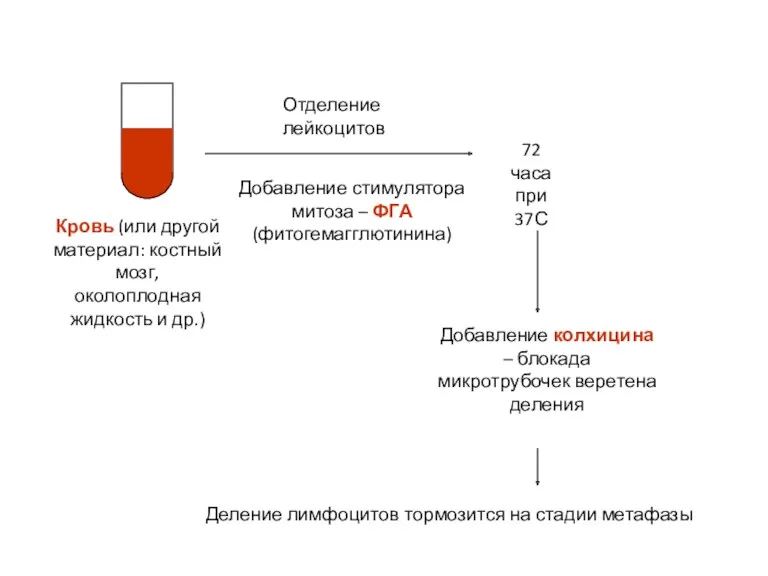
Кровь (или другой материал: костный мозг, околоплодная жидкость и др.)
Отделение лейкоцитов
Добавление
Кровь (или другой материал: костный мозг, околоплодная жидкость и др.)
Отделение лейкоцитов
Добавление
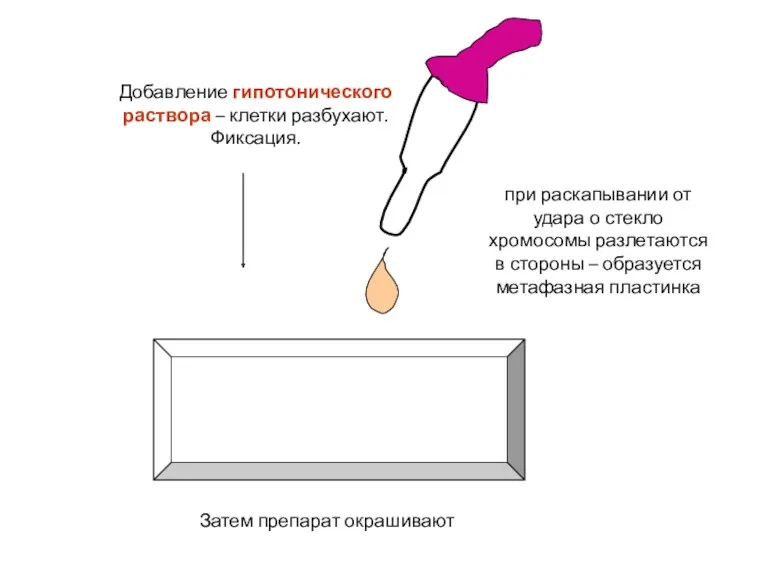
Добавление гипотонического раствора – клетки разбухают. Фиксация.
Х ХХ
х х
при
Добавление гипотонического раствора – клетки разбухают. Фиксация.
Х ХХ
х х
при
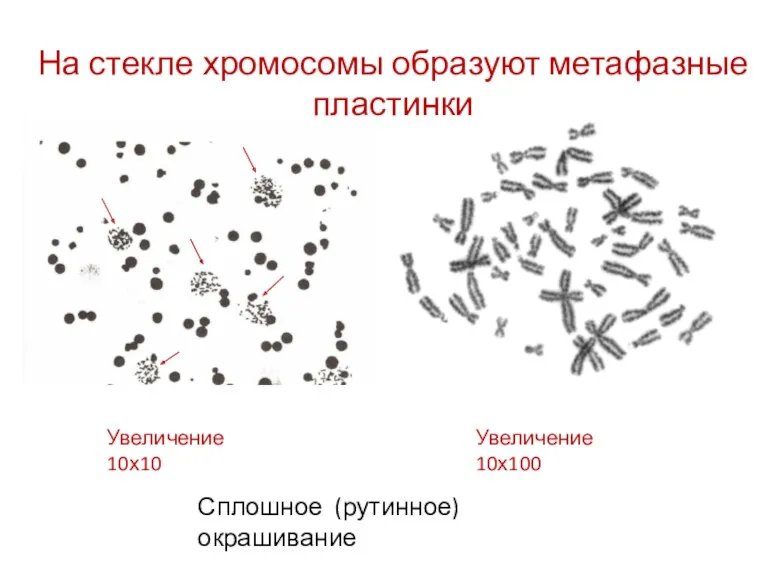
На стекле хромосомы образуют метафазные пластинки
Сплошное (рутинное) окрашивание
На стекле хромосомы образуют метафазные пластинки
Сплошное (рутинное) окрашивание

Виды окраски хромосом
Рутинная, появилась в 50-х годах ХХ века. (Денверская классификация
Виды окраски хромосом
Рутинная, появилась в 50-х годах ХХ века. (Денверская классификация
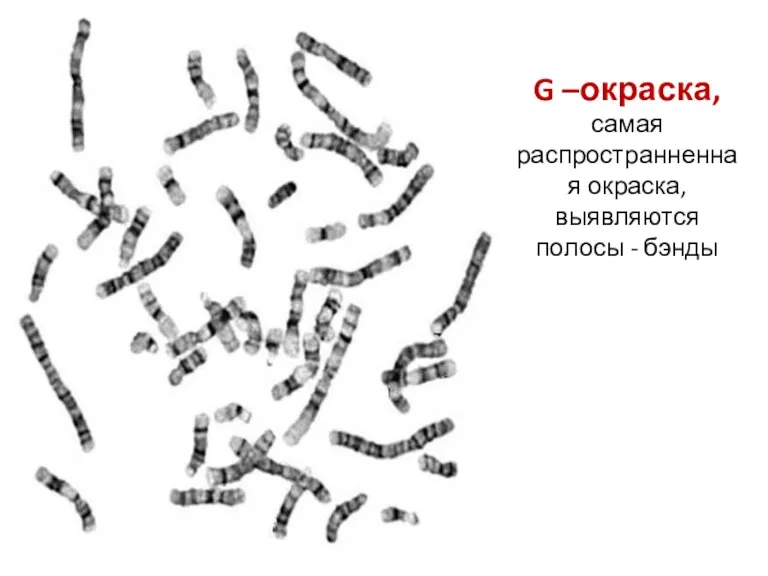
G –окраска, самая распространненная окраска, выявляются полосы - бэнды
G –окраска, самая распространненная окраска, выявляются полосы - бэнды
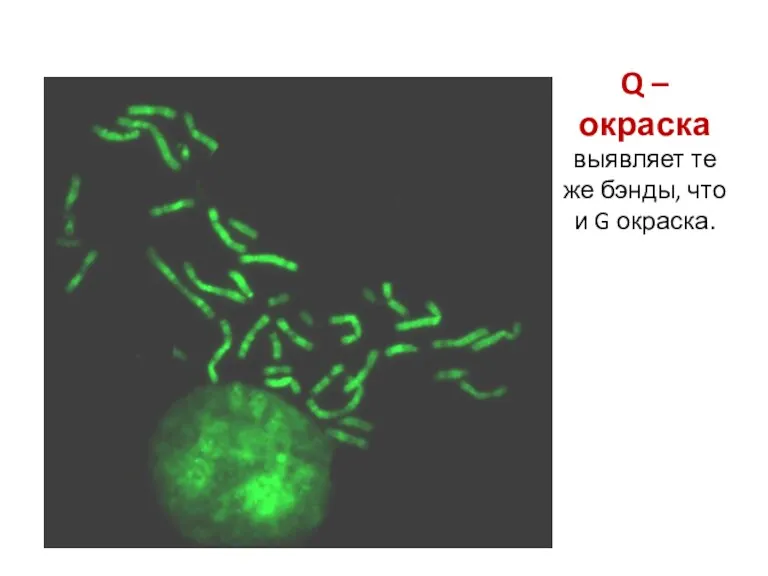
Q –окраска выявляет те же бэнды, что и G окраска.
Q –окраска выявляет те же бэнды, что и G окраска.

C –окраска
выявляет конститутивный гетерохроматин
C –окраска
выявляет конститутивный гетерохроматин

Окраска азотнокислым серебром
выявляет ядрышко-образующие районы хромосом
Окраска азотнокислым серебром
выявляет ядрышко-образующие районы хромосом

Ядрышко-образующие районы хромосом
Ядрышко-образующие районы хромосом (ЯОР) локализованы в коротких плечах акроцентрических
Ядрышко-образующие районы хромосом
Ядрышко-образующие районы хромосом (ЯОР) локализованы в коротких плечах акроцентрических
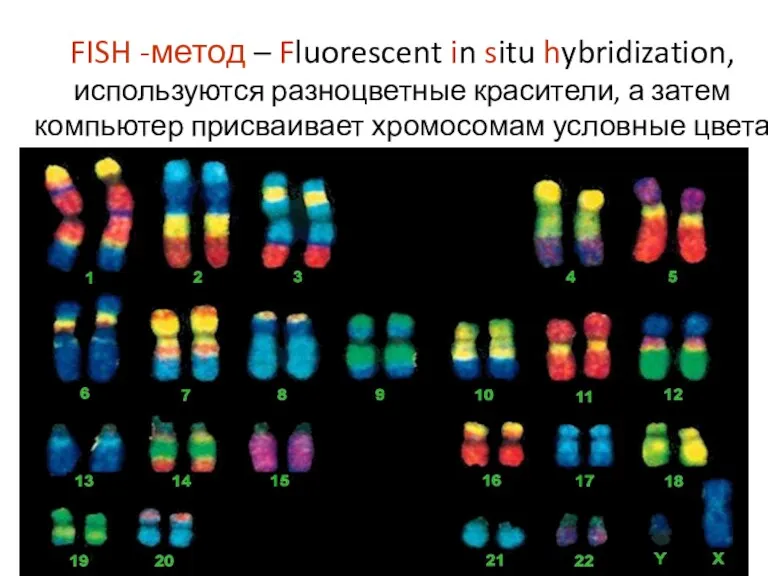
FISH -метод – Fluorescent in situ hybridization, используются разноцветные красители, а
FISH -метод – Fluorescent in situ hybridization, используются разноцветные красители, а
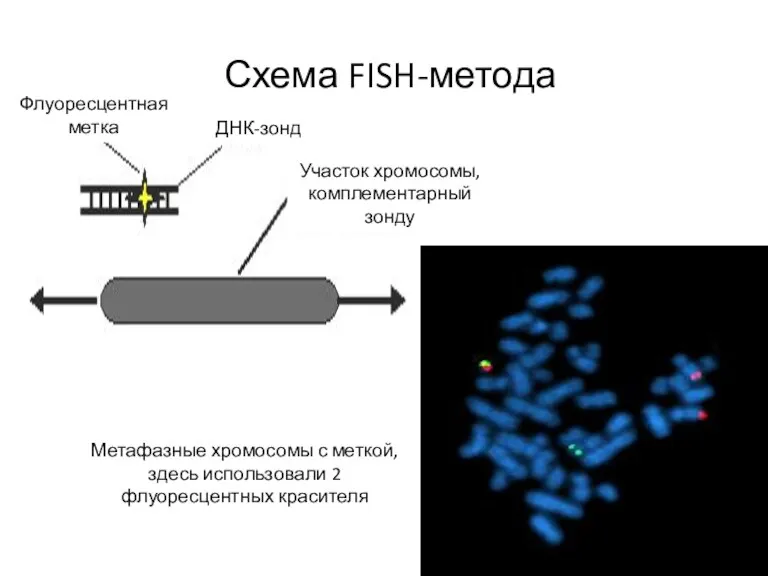
Схема FISH-метода
Флуоресцентная метка
ДНК-зонд
Участок хромосомы, комплементарный зонду
Метафазные хромосомы с меткой, здесь использовали
Схема FISH-метода
Флуоресцентная метка
ДНК-зонд
Участок хромосомы, комплементарный зонду
Метафазные хромосомы с меткой, здесь использовали
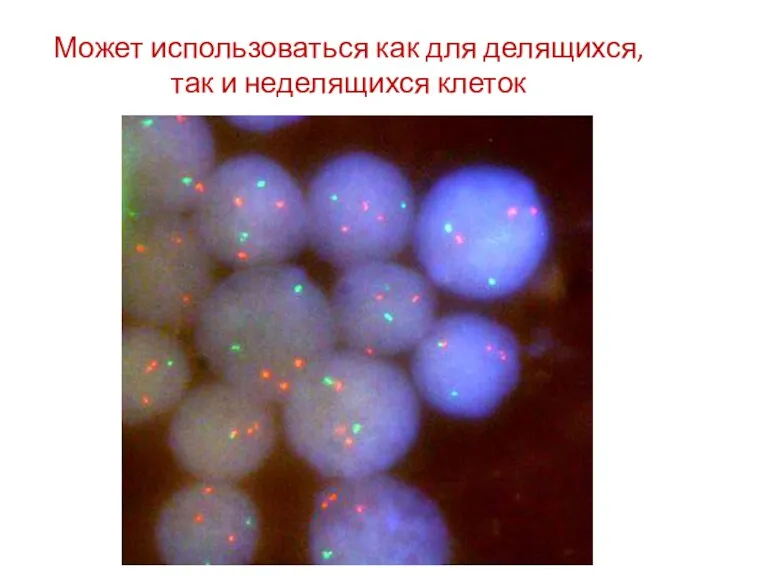
Может использоваться как для делящихся, так и неделящихся клеток
Может использоваться как для делящихся, так и неделящихся клеток
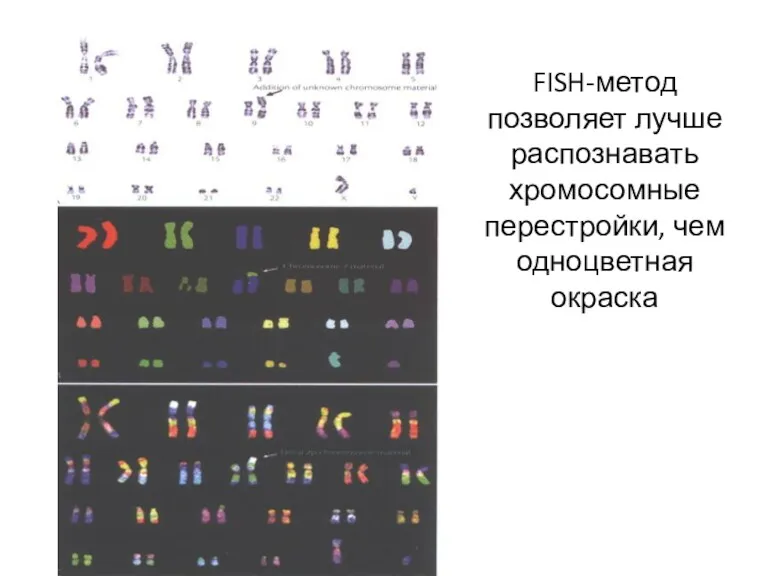
FISH-метод позволяет лучше распознавать хромосомные перестройки, чем одноцветная окраска
FISH-метод позволяет лучше распознавать хромосомные перестройки, чем одноцветная окраска

Классификация (номенклатура) хромосом
Классификация (номенклатура) хромосом
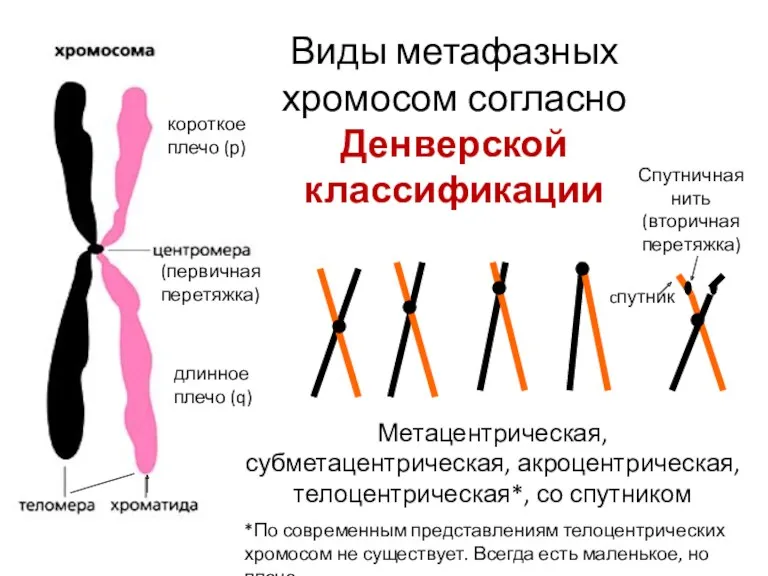
Виды метафазных хромосом согласно Денверской классификации
Метацентрическая, субметацентрическая, акроцентрическая, телоцентрическая*, со спутником
*По
Виды метафазных хромосом согласно Денверской классификации
Метацентрическая, субметацентрическая, акроцентрическая, телоцентрическая*, со спутником
*По

Хромосомы человека по Денверской классификации делят на 7 групп
Хромосомы человека по Денверской классификации делят на 7 групп

Парижская классификация дополняет Денверскую и основана на дифференциальной окраске (чаще всего
Парижская классификация дополняет Денверскую и основана на дифференциальной окраске (чаще всего
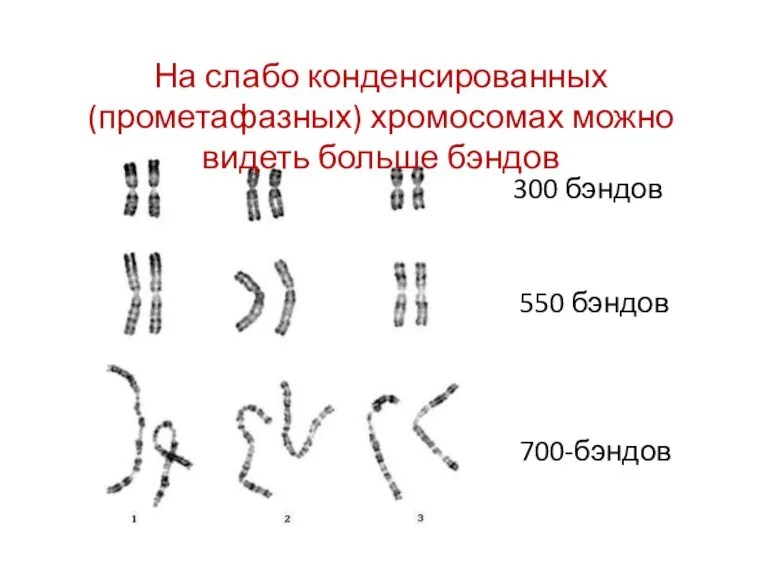
300 бэндов
550 бэндов
700-бэндов
На слабо конденсированных (прометафазных) хромосомах можно видеть больше бэндов
300 бэндов
550 бэндов
700-бэндов
На слабо конденсированных (прометафазных) хромосомах можно видеть больше бэндов
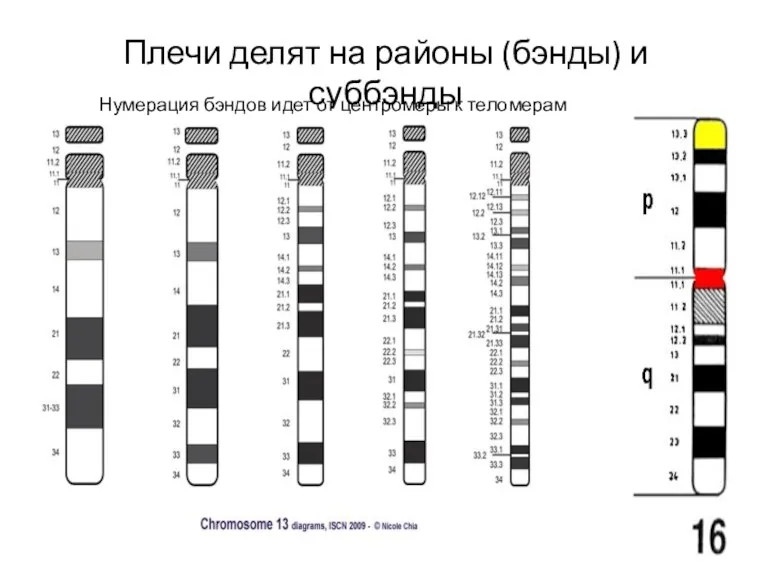
Плечи делят на районы (бэнды) и суббэнды
Нумерация бэндов идет от центромеры
Плечи делят на районы (бэнды) и суббэнды
Нумерация бэндов идет от центромеры
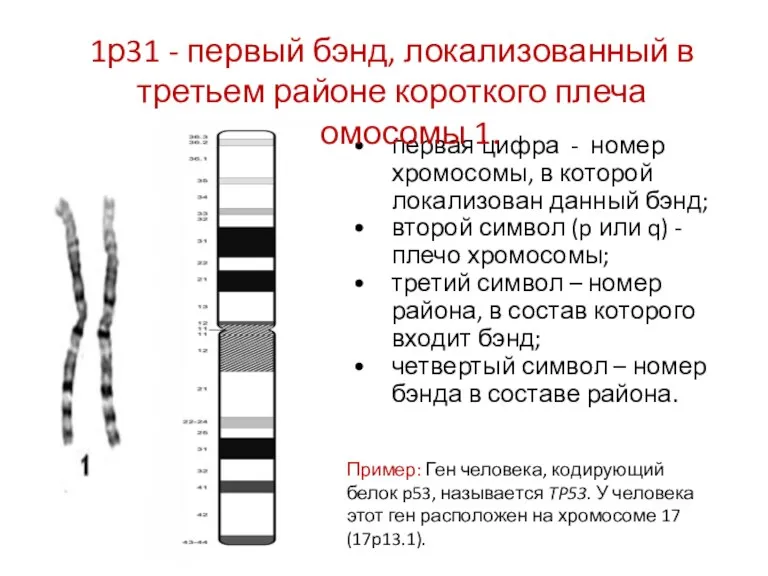
первая цифра - номер хромосомы, в которой локализован данный бэнд;
второй символ
первая цифра - номер хромосомы, в которой локализован данный бэнд;
второй символ
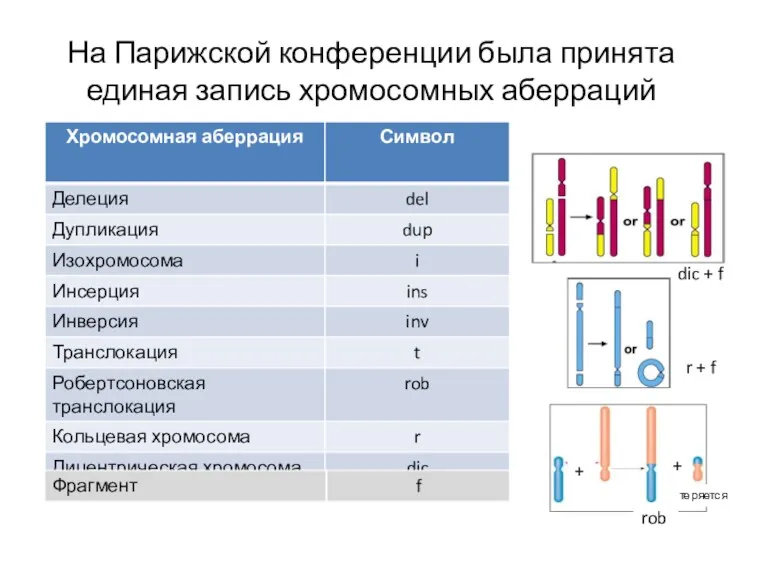
На Парижской конференции была принята единая запись хромосомных аберраций
Фрагмент f
+
+
теряется
rob
r
На Парижской конференции была принята единая запись хромосомных аберраций
Фрагмент f
+
+
теряется
rob
r
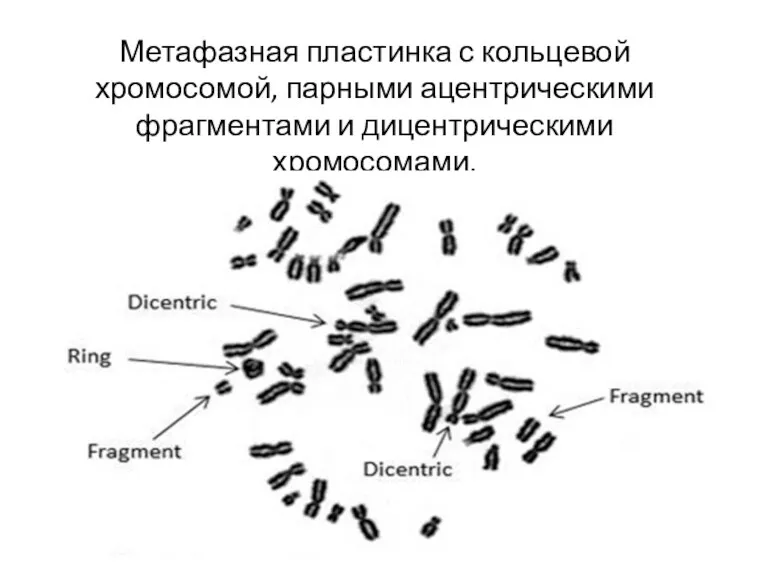
Метафазная пластинка с кольцевой хромосомой, парными ацентрическими фрагментами и дицентрическими хромосомами.
Метафазная пластинка с кольцевой хромосомой, парными ацентрическими фрагментами и дицентрическими хромосомами.

Трисомия записывается знаком «+»
Трисомия записывается знаком «+»
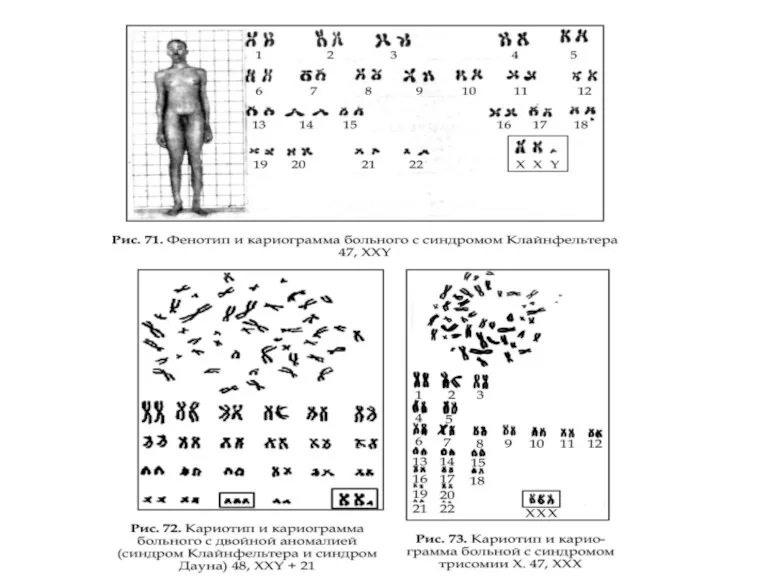

Примеры записи хромосомного диагноза
46,XX обычная женщина
46,XY обычный мужчина
69,XXY
Примеры записи хромосомного диагноза
46,XX обычная женщина
46,XY обычный мужчина
69,XXY

V Международный конгресс по генетике человека (Мехико, 1972г.) оформил официальную номенклатуру
V Международный конгресс по генетике человека (Мехико, 1972г.) оформил официальную номенклатуру

Необычные хромосомы
Необычные хромосомы

Необычные виды хромосом
«ламповые щетки». Впервые хромосомы типа ламповых щёток были описаны
Необычные виды хромосом
«ламповые щетки». Впервые хромосомы типа ламповых щёток были описаны
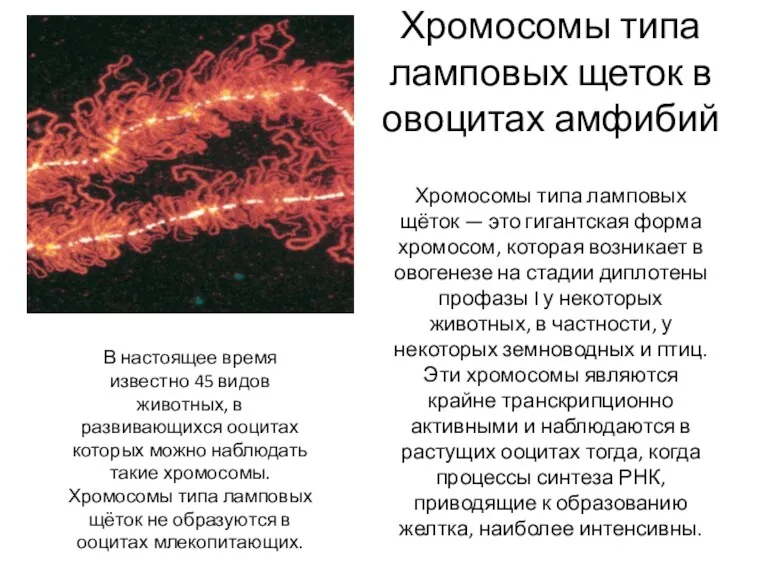
Хромосомы типа ламповых щеток в овоцитах амфибий
Хромосомы типа ламповых щёток — это
Хромосомы типа ламповых щеток в овоцитах амфибий
Хромосомы типа ламповых щёток — это
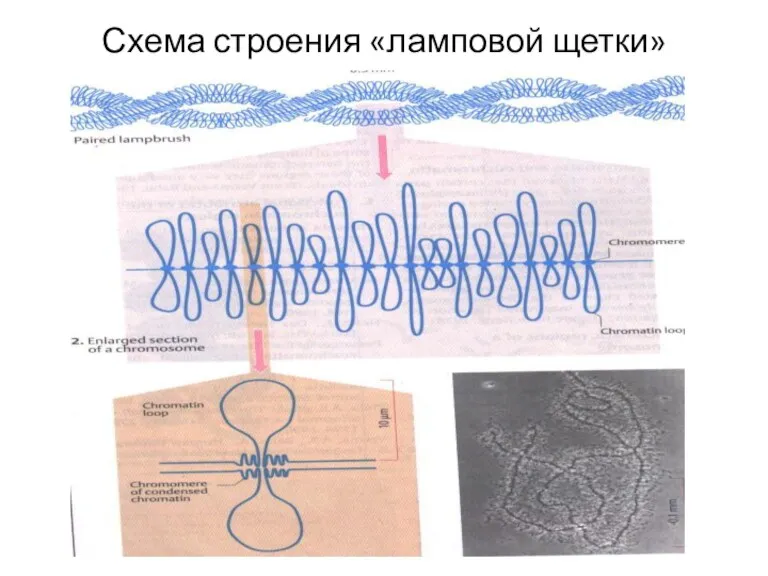
Схема строения «ламповой щетки»
Схема строения «ламповой щетки»
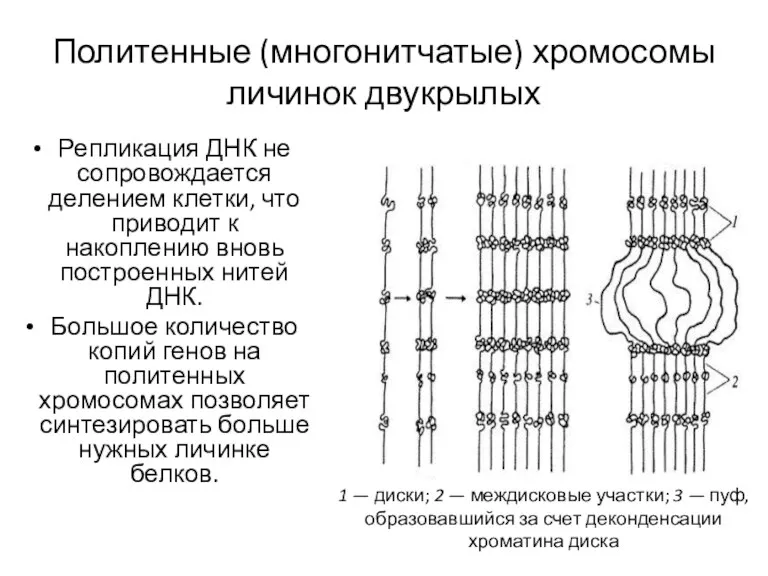
Политенные (многонитчатые) хромосомы личинок двукрылых
Репликация ДНК не сопровождается делением клетки, что
Политенные (многонитчатые) хромосомы личинок двукрылых
Репликация ДНК не сопровождается делением клетки, что
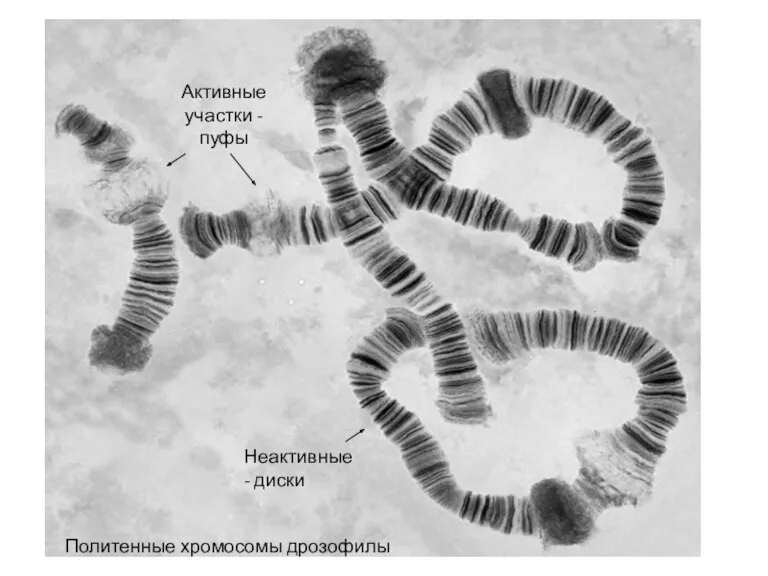
Активные участки -пуфы
Неактивные - диски
Политенные хромосомы дрозофилы
Активные участки -пуфы
Неактивные - диски
Политенные хромосомы дрозофилы

Хромосомные карты
Хромосомные карты

Хромосомные карты делят на
Генетические – показывают, где лежит какой ген
Цитологические –
Хромосомные карты делят на
Генетические – показывают, где лежит какой ген
Цитологические –

Основные методы составления генетических (хромосомных) карт
На основе скрещиваний - не у
Основные методы составления генетических (хромосомных) карт
На основе скрещиваний - не у

Расстояние генов В и V – 17 морганид
Первым организмом, для которого
Расстояние генов В и V – 17 морганид
Первым организмом, для которого
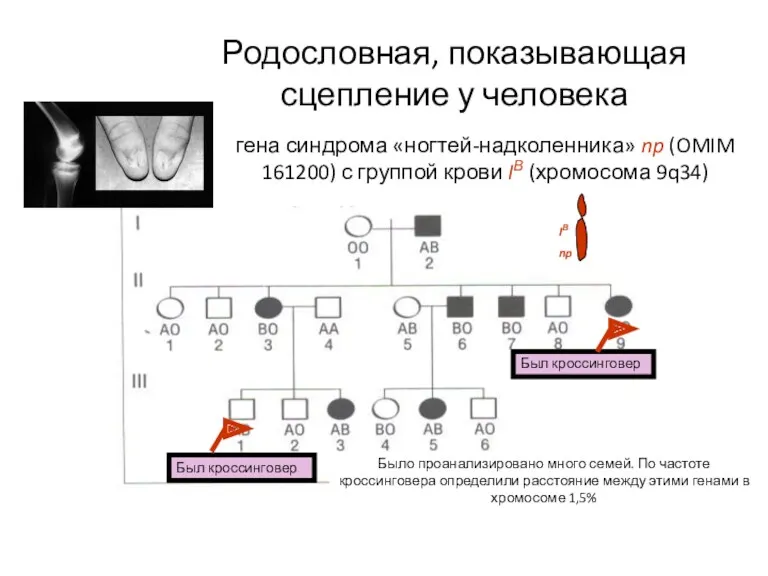
Родословная, показывающая сцепление у человека
Был кроссинговер
Был кроссинговер
Было проанализировано много семей. По
Родословная, показывающая сцепление у человека
Был кроссинговер
Был кроссинговер
Было проанализировано много семей. По

Метод изучения генетики соматических клеток
Клетки выращивают в культуре.
Этим методом удалось
Метод изучения генетики соматических клеток
Клетки выращивают в культуре.
Этим методом удалось
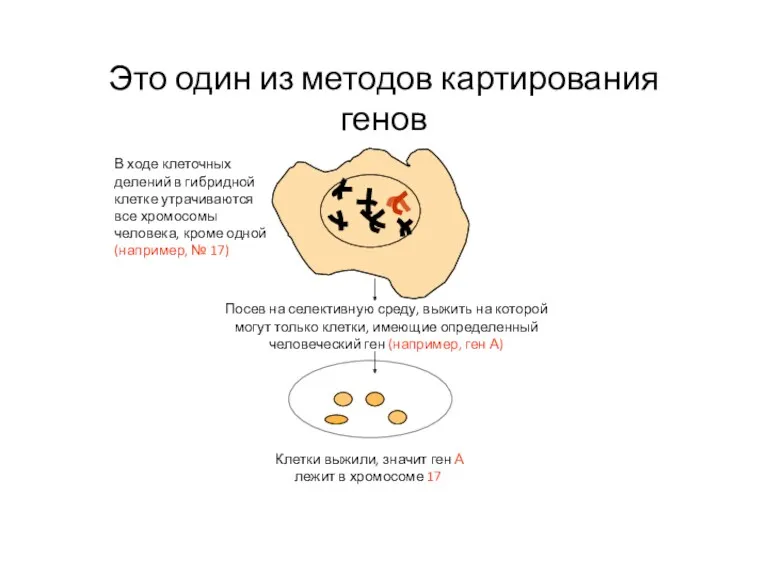
В ходе клеточных делений в гибридной клетке утрачиваются все хромосомы человека,
В ходе клеточных делений в гибридной клетке утрачиваются все хромосомы человека,
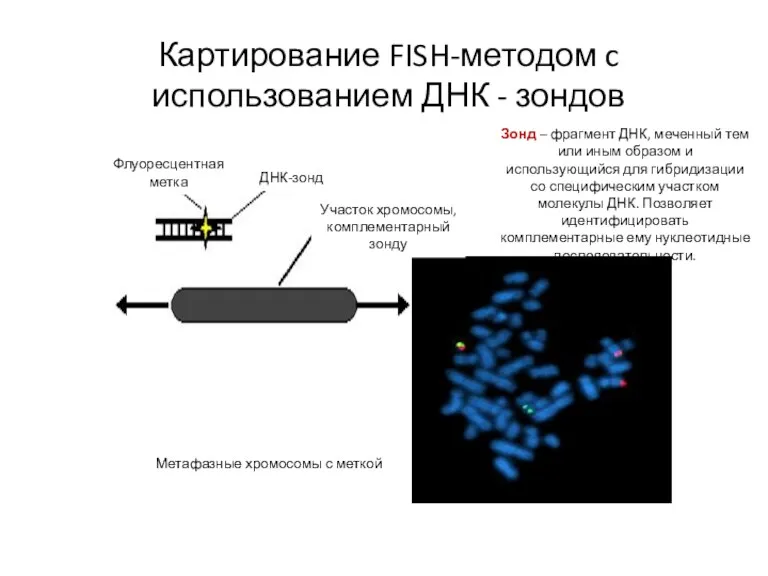
Картирование FISH-методом c использованием ДНК - зондов
Флуоресцентная метка
ДНК-зонд
Участок хромосомы, комплементарный зонду
Метафазные
Картирование FISH-методом c использованием ДНК - зондов
Флуоресцентная метка
ДНК-зонд
Участок хромосомы, комплементарный зонду
Метафазные

Карта хромосомы 9 – пример комбинированной карты
Карта хромосомы 9 – пример комбинированной карты
 Продукты питания, ядовитые для собак и кошек
Продукты питания, ядовитые для собак и кошек Энергетический обмен
Энергетический обмен Проект Мои домашние питомцы
Проект Мои домашние питомцы Корисні бактерії
Корисні бактерії Вирусы в биотехнологии
Вирусы в биотехнологии Биологическая роль железа
Биологическая роль железа Coleoptera
Coleoptera История зоологии. Общие сведения о животном мире
История зоологии. Общие сведения о животном мире Удивительные и необычные деревья
Удивительные и необычные деревья Формы размножения организмов
Формы размножения организмов Плоды, семена
Плоды, семена Органы пищеварения
Органы пищеварения Щелочные и щелочноземельные металлы и их роль в организме человека
Щелочные и щелочноземельные металлы и их роль в организме человека Tetracyclines, Chloramphenicol, Aminoglycosides
Tetracyclines, Chloramphenicol, Aminoglycosides Адаптация организмов к условиям обитания
Адаптация организмов к условиям обитания Біологічна продуктивність водойм. Охорона водойм від забруднення
Біологічна продуктивність водойм. Охорона водойм від забруднення Биотестирование. Компоненты современной системы оценки антропогенного влияния на окружающую среду
Биотестирование. Компоненты современной системы оценки антропогенного влияния на окружающую среду Презентация Отдел Голосеменные
Презентация Отдел Голосеменные Морфология и ультраструктура бактерий
Морфология и ультраструктура бактерий Көмірсулардың катаболиттік және анаболиттік жолдарын айырбастау және көмірсулардың биологиялық рөлі туралы түсінік беру
Көмірсулардың катаболиттік және анаболиттік жолдарын айырбастау және көмірсулардың биологиялық рөлі туралы түсінік беру Экспрессия генов. Транскрипция, трансляция
Экспрессия генов. Транскрипция, трансляция Глаукус. Строение
Глаукус. Строение Музей Квітки Цісик
Музей Квітки Цісик Дагестанский заповедник у Кизлярского залива
Дагестанский заповедник у Кизлярского залива Отряд Чешуйчатые. Подотряд Ящерицы
Отряд Чешуйчатые. Подотряд Ящерицы Молекулярные основы наследственности
Молекулярные основы наследственности Введение в анатомию. Общая анатомия скелета
Введение в анатомию. Общая анатомия скелета Северный олень
Северный олень