- Вирусы в биотехнологии

Содержание
- 2. Субвирусные инфекционные агенты Субвирусные инфекционные агенты: 1. вироиды 2. саттелиты
- 3. вироиды Вироиды – маленькие однонитевые кольцевые молекулы РНК, не кодирующие вирусные белки, лишенные белкового капсида Термин
- 4. Вироиды Вироиды заражают высшие растения Им не нужен вирус-помощник Реплицируются в ядре или в хлоропластах с
- 5. вироиды Классификация основана а первичной нуклеотидной последовательности в РНК В настоящее время определены 30 вироидов, которые
- 6. вироиды Pospiviroidae реплицируются в ядре клетки, используя клеточные ДНК-зависимую РНК-полимеразу, а также эндонуклеазу и лигазу .
- 7. вироиды Патогенность вироидов связывают с влиянием вироидной РНК на синтез белка. Вироиды, реплицирующиеся в ядре влияют
- 8. Саттелиты Саттелиты –субвирусные агенты, неспособные заражать хозяйские клетки без вируса-помощника Саттелиты реплицируются на матрице своей собственной
- 9. саттелиты Саттелиты встречаются у вирусов растений, грибов, бактерий, животных. Они обладают рядом общих свойств: 1. Генетичекий
- 10. вирусоиды ВИРУСОИДЫ – имеют кольцевые молекулы РНК, размером 350 нуклеотидов, которые одеваются вместе с вирусной РНК
- 11. Прионы Прионы – инфекционные агенты белковой природы( Prions – Proteinaceous Infection Particles) PrPc. PrPsc вызывающие у
- 12. ПРИОНЫ 1732 в Англии была описана болезнь овец скрепи (scrape- скоблить, тереться), при которой овцы начинали
- 13. Прионы
- 14. Прионы В 1982 году Стенли Прузинер выдвинул гипотезу, что причиной губкообразных трансмиссивных энцефалопатий является белок. Он
- 15. прионы Синтезируется главным образом в нейронах. Обнаружен в селезенке, лимфатических узлах, коже, ЖКТ, фолликулярных дендритных клетках,
- 16. прионы Молекула нормального приона состоит из 4 альфа-спиральных доменов, стабилизированных междоменными электростатическими взаимодействиями и S-S1 –
- 17. Прионы Накопление конформационно измененного белка сопровождается: его агрегацией, образованием высоко упорядочных фибрилл (амелоидов), приводит к гибели
- 18. прионы
- 19. прионы Именно С-терминальный участок конформационно измененной формы, PrPsc , становится резистентным к протеазе Превращение нормального белка
- 20. прионы
- 21. прионы Измененные прионы устойчивы : 1. к протеолизу 2. к излучениям 3. к высокой температуре 4.
- 23. Прионы дрожжей Наличие прионов в дрожжах было установлено в 80х годах Представляют конформационные варианты обычных клеточных
- 24. Прионы дрожжей Дрожжевой транскрипционный ко-репрессор Ure-2p может существовать в 2 стабильных конформационных формах: 1. активной как
- 25. Бактерии рода Bartonella имеют гемин-связывающий белок (Hbp), входящий в мембрану клетки. Является порин-подобным белком, обладающим спосоюностью
- 26. Прионы дрожжей Прионы дрожжей не приводят к гибели клеток. Они повышают выживаемость в неблагоприятных условиях (
- 27. Морфологические формы фагов
- 29. Умеренный бактериофаг
- 30. Механизм лизогении Продукт гена С I является репрессором, подавляющим работу промотеров PI и Prm . Он
- 31. Механизм лизогении Продукт гена С I является репрессором, подавляющим работу промотеров PI и Prm . Он
- 32. Механизм установления лизогении
- 33. Механизм лизогении В лизогенной бактериальной клетке на низком уровне происходит синтез репрессора СI Если под действием
- 34. Механизм лизогении В «голодной» бактериальной клетке находится высокий уровень ц-АМФ, который активирует ген СII , продукт
- 35. Вирулентный фаг Умеренный фаг Форма бляшек
- 36. Лизогенная (фаговая) конверсия
- 37. Лизогенная (фаговая) конверсия Фаговая конверсия имеет значение в медицине: лизогенными бактериями вырабатываются эритрогенный токсин возбудителя скарлатины;
- 38. Роль фагов в биотехнологии Благодаря концентрации больших количеств микробных клеток обеспечение стабильности биотехнологических производств зависит от
- 39. Роль фагов в биотехнологии При использовании нового штамма-продуцента предварительно необходимо проводить его тестирование на лизогенность Для
- 40. Роль фагов в биотехнологии Для производства важно определение характера бактеритофагов, лизирующих производственную культуру, а также источник
- 41. Роль фагов в биотехнологии Источником инфицирования производства бактериофагами может быть и сырье, например- молоко при получении
- 42. Практическое использование бактериофагов Бактериофаги могут быть использованы для диагностики, идентификации выделенных бактерий На чашку со средой,
- 43. фаготипирование
- 44. Практическое использование бактериофагов Наличие фагов кишечной палочки и возбудителей кишечных инфекций в водоисточниках является показателем их
- 45. Бактериофаги –антибактериальные препараты Лечебно-профилактические бактериофаги – антимикробные препараты, которые содержат высоковирулентные бактериальные фаги и обладают селективным
- 46. Лечебно-профилактические бактериофаги – антимикробные препараты, которые содержат высоковирулентные бактериальные фаги и обладают селективным антибактериальным действием Преимущества:
- 47. Технология производства препаратов бактериофагов 1. Подбор активных природных фаговых клонов, не способных лизогенировать бактерию и обладающих
- 48. КОСМИДА cosmid vector - космида.Векторная плазмида , содержащая cos-участок ДНК фага лямбда, который является местом замыкания
- 52. Рга
- 53. РГА РГА позволяет обнаружить (провести индикацию )вирус: в зараженном курином эмбрионе культуральной жидкости зараженной вирусом культуре
- 55. Метод бляшек
- 56. Вирусы и нанотехнология Вирионы могут быть реконструированы в пробирке из белка оболочки и нуклеиновой кислоты. Получают
- 57. Вирусы и нанотехнология Белок оболочки бактериофага М13 связали с аморфным фосфатом железа, способным обратимо присоединять и
- 59. Скачать презентацию

Субвирусные инфекционные агенты
Субвирусные инфекционные агенты:
1. вироиды
2. саттелиты
Субвирусные инфекционные агенты
Субвирусные инфекционные агенты:
1. вироиды
2. саттелиты

вироиды
Вироиды – маленькие однонитевые кольцевые молекулы РНК, не кодирующие вирусные белки,
вироиды
Вироиды – маленькие однонитевые кольцевые молекулы РНК, не кодирующие вирусные белки,

Вироиды
Вироиды заражают высшие растения
Им не нужен вирус-помощник
Реплицируются в ядре или в
Вироиды
Вироиды заражают высшие растения
Им не нужен вирус-помощник
Реплицируются в ядре или в

вироиды
Классификация основана а первичной нуклеотидной последовательности в РНК
В настоящее время
вироиды
Классификация основана а первичной нуклеотидной последовательности в РНК
В настоящее время

вироиды
Pospiviroidae реплицируются в ядре клетки, используя клеточные ДНК-зависимую РНК-полимеразу, а также
вироиды
Pospiviroidae реплицируются в ядре клетки, используя клеточные ДНК-зависимую РНК-полимеразу, а также

вироиды
Патогенность вироидов связывают с влиянием вироидной РНК на синтез белка.
Вироиды, реплицирующиеся
вироиды
Патогенность вироидов связывают с влиянием вироидной РНК на синтез белка.
Вироиды, реплицирующиеся

Саттелиты
Саттелиты –субвирусные агенты, неспособные заражать хозяйские клетки без вируса-помощника
Саттелиты реплицируются на
Саттелиты
Саттелиты –субвирусные агенты, неспособные заражать хозяйские клетки без вируса-помощника
Саттелиты реплицируются на

саттелиты
Саттелиты встречаются у вирусов растений, грибов, бактерий, животных. Они обладают рядом
саттелиты
Саттелиты встречаются у вирусов растений, грибов, бактерий, животных. Они обладают рядом

вирусоиды
ВИРУСОИДЫ – имеют кольцевые молекулы РНК, размером 350 нуклеотидов, которые одеваются
вирусоиды
ВИРУСОИДЫ – имеют кольцевые молекулы РНК, размером 350 нуклеотидов, которые одеваются

Прионы
Прионы – инфекционные агенты белковой природы( Prions – Proteinaceous Infection Particles)
Прионы
Прионы – инфекционные агенты белковой природы( Prions – Proteinaceous Infection Particles)

ПРИОНЫ
1732 в Англии была описана болезнь овец скрепи (scrape- скоблить, тереться),
ПРИОНЫ
1732 в Англии была описана болезнь овец скрепи (scrape- скоблить, тереться),

Прионы
Прионы

Прионы
В 1982 году Стенли Прузинер выдвинул гипотезу, что причиной губкообразных трансмиссивных
Прионы
В 1982 году Стенли Прузинер выдвинул гипотезу, что причиной губкообразных трансмиссивных

прионы
Синтезируется главным образом в нейронах.
Обнаружен в селезенке, лимфатических узлах, коже,
прионы
Синтезируется главным образом в нейронах.
Обнаружен в селезенке, лимфатических узлах, коже,

прионы
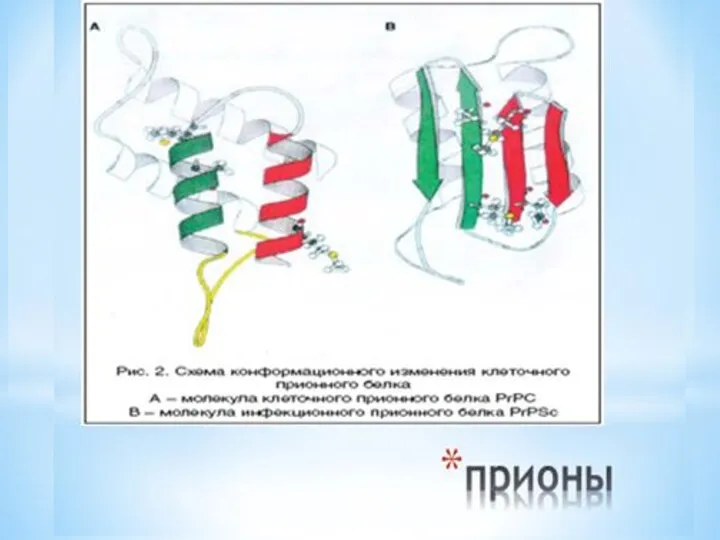
Молекула нормального приона состоит из 4 альфа-спиральных доменов, стабилизированных междоменными электростатическими
прионы
Молекула нормального приона состоит из 4 альфа-спиральных доменов, стабилизированных междоменными электростатическими

Прионы
Накопление конформационно измененного белка сопровождается:
его агрегацией,
образованием высоко упорядочных фибрилл
Прионы
Накопление конформационно измененного белка сопровождается:
его агрегацией,
образованием высоко упорядочных фибрилл

прионы
прионы

прионы
Именно С-терминальный участок конформационно измененной формы, PrPsc , становится резистентным к
прионы
Именно С-терминальный участок конформационно измененной формы, PrPsc , становится резистентным к
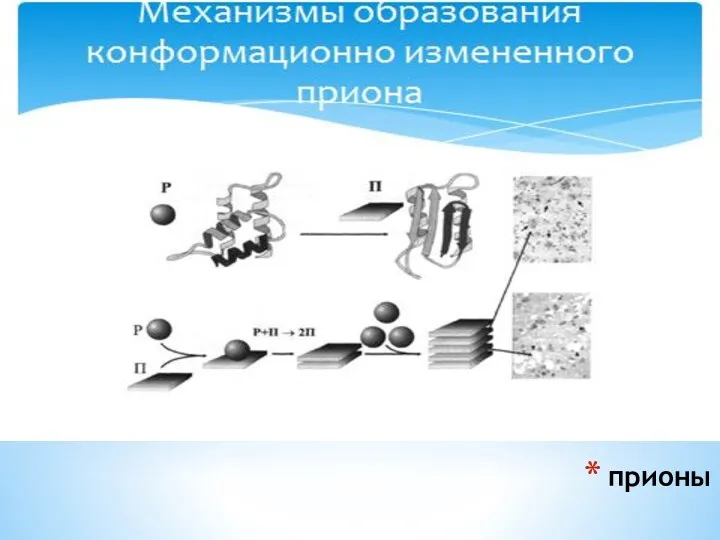
прионы
прионы

прионы
Измененные прионы устойчивы :
1. к протеолизу
2. к излучениям
3. к высокой
прионы
Измененные прионы устойчивы :
1. к протеолизу
2. к излучениям
3. к высокой

Прионы дрожжей
Наличие прионов в дрожжах было установлено в 80х годах
Представляют конформационные
Прионы дрожжей
Наличие прионов в дрожжах было установлено в 80х годах
Представляют конформационные

Прионы дрожжей
Дрожжевой транскрипционный ко-репрессор Ure-2p может существовать в 2 стабильных конформационных
Прионы дрожжей
Дрожжевой транскрипционный ко-репрессор Ure-2p может существовать в 2 стабильных конформационных

Бактерии рода Bartonella имеют гемин-связывающий белок (Hbp), входящий в мембрану клетки.
Является
Бактерии рода Bartonella имеют гемин-связывающий белок (Hbp), входящий в мембрану клетки.
Является

Прионы дрожжей
Прионы дрожжей не приводят к гибели клеток.
Они повышают выживаемость
Прионы дрожжей
Прионы дрожжей не приводят к гибели клеток.
Они повышают выживаемость
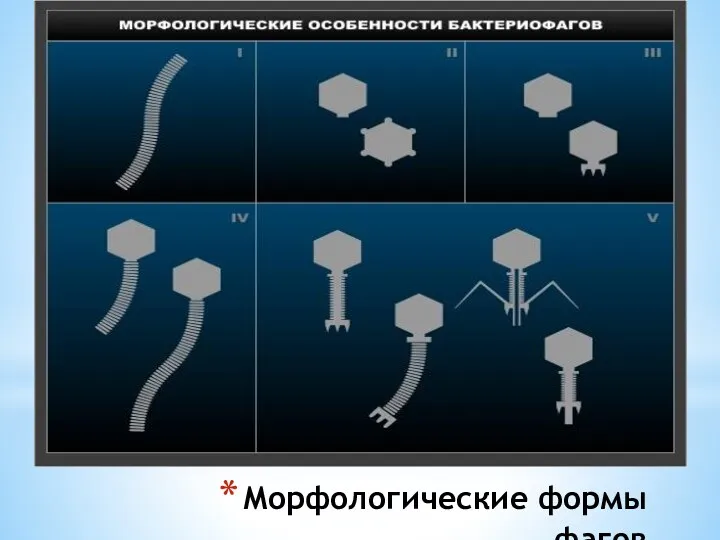
Морфологические формы фагов
Морфологические формы фагов

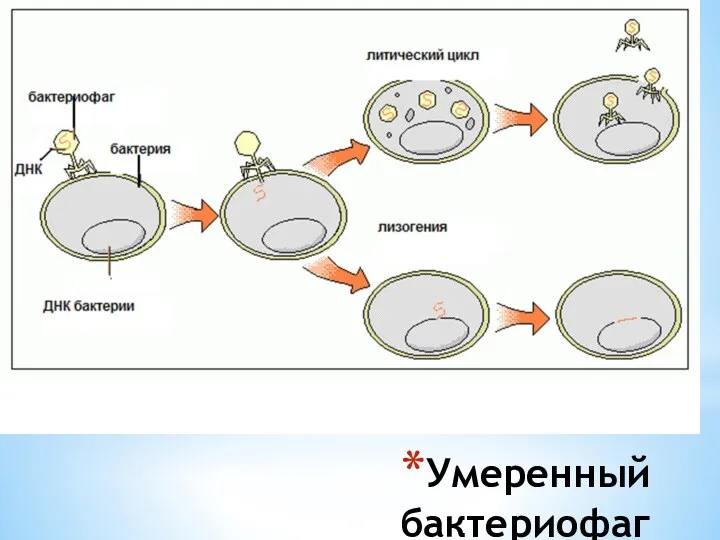
Умеренный бактериофаг
Умеренный бактериофаг
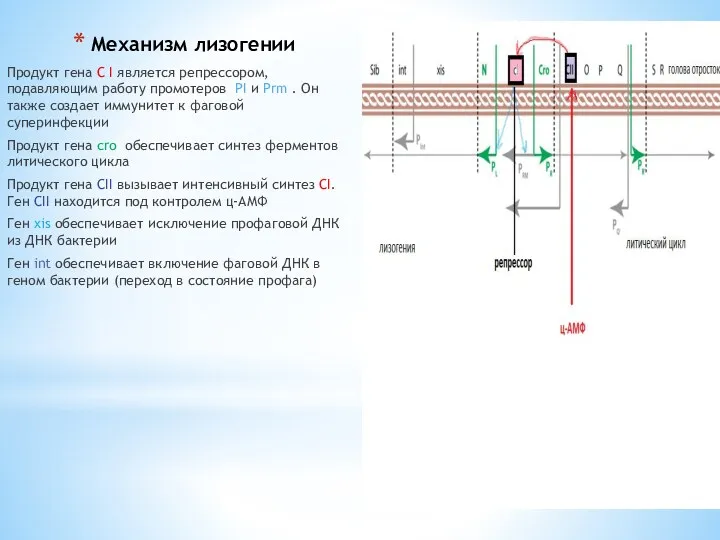
Механизм лизогении
Продукт гена С I является репрессором, подавляющим работу промотеров PI
Механизм лизогении
Продукт гена С I является репрессором, подавляющим работу промотеров PI

Механизм лизогении
Продукт гена С I является репрессором, подавляющим работу промотеров PI
Механизм лизогении
Продукт гена С I является репрессором, подавляющим работу промотеров PI
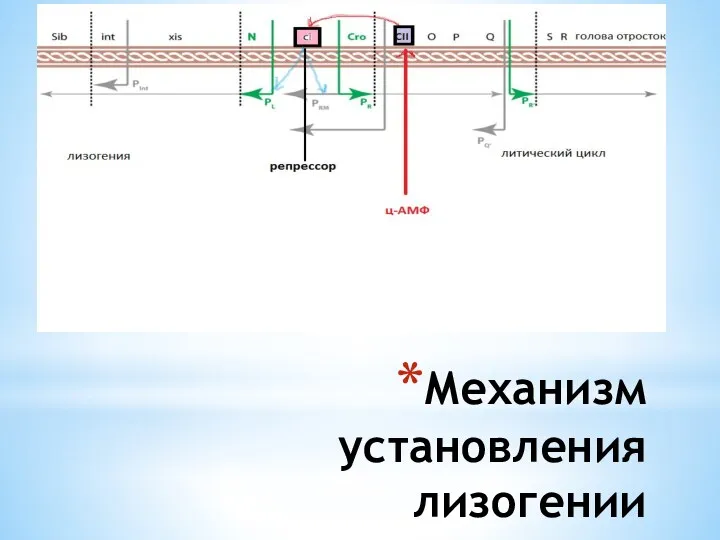
Механизм установления лизогении
Механизм установления лизогении

Механизм лизогении
В лизогенной бактериальной клетке на низком уровне происходит синтез репрессора
Механизм лизогении
В лизогенной бактериальной клетке на низком уровне происходит синтез репрессора

Механизм лизогении
В «голодной» бактериальной клетке находится высокий уровень ц-АМФ, который активирует
Механизм лизогении
В «голодной» бактериальной клетке находится высокий уровень ц-АМФ, который активирует
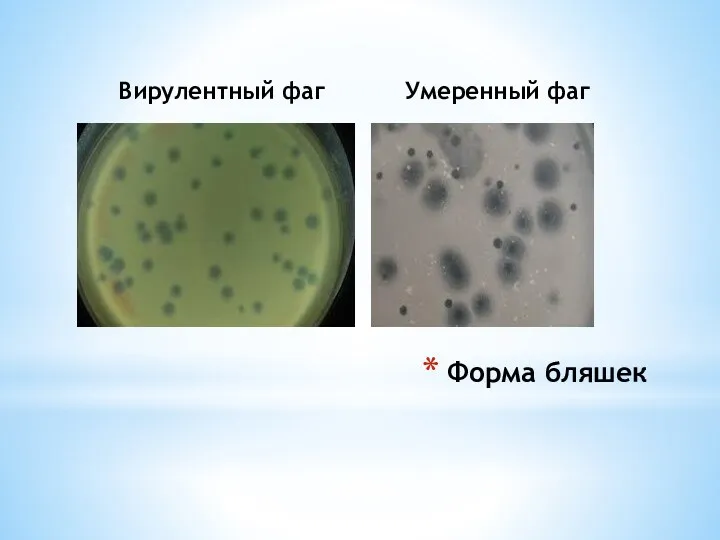
Вирулентный фаг
Умеренный фаг
Форма бляшек
Вирулентный фаг
Умеренный фаг
Форма бляшек
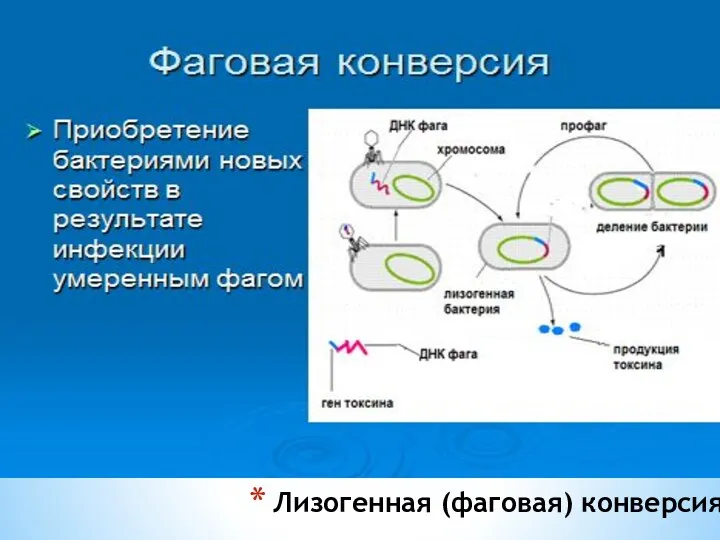
Лизогенная (фаговая) конверсия
Лизогенная (фаговая) конверсия

Лизогенная (фаговая) конверсия
Фаговая конверсия имеет значение в медицине: лизогенными бактериями вырабатываются
Лизогенная (фаговая) конверсия
Фаговая конверсия имеет значение в медицине: лизогенными бактериями вырабатываются

Роль фагов в биотехнологии
Благодаря концентрации больших количеств микробных клеток обеспечение стабильности
Роль фагов в биотехнологии
Благодаря концентрации больших количеств микробных клеток обеспечение стабильности

Роль фагов в биотехнологии
При использовании нового штамма-продуцента предварительно необходимо проводить его
Роль фагов в биотехнологии
При использовании нового штамма-продуцента предварительно необходимо проводить его

Роль фагов в биотехнологии
Для производства важно определение характера бактеритофагов, лизирующих производственную
Роль фагов в биотехнологии
Для производства важно определение характера бактеритофагов, лизирующих производственную

Роль фагов в биотехнологии
Источником инфицирования производства бактериофагами может быть и сырье,
Роль фагов в биотехнологии
Источником инфицирования производства бактериофагами может быть и сырье,

Практическое использование бактериофагов
Бактериофаги могут быть использованы для диагностики, идентификации выделенных бактерий
На
Практическое использование бактериофагов
Бактериофаги могут быть использованы для диагностики, идентификации выделенных бактерий
На

фаготипирование
фаготипирование

Практическое использование бактериофагов
Наличие фагов кишечной палочки и возбудителей кишечных инфекций в
Практическое использование бактериофагов
Наличие фагов кишечной палочки и возбудителей кишечных инфекций в

Бактериофаги –антибактериальные препараты
Лечебно-профилактические бактериофаги – антимикробные препараты, которые содержат высоковирулентные бактериальные
Бактериофаги –антибактериальные препараты
Лечебно-профилактические бактериофаги – антимикробные препараты, которые содержат высоковирулентные бактериальные

Лечебно-профилактические бактериофаги – антимикробные препараты, которые содержат высоковирулентные бактериальные фаги и
Лечебно-профилактические бактериофаги – антимикробные препараты, которые содержат высоковирулентные бактериальные фаги и

Технология производства препаратов бактериофагов
1. Подбор активных природных фаговых клонов, не способных
Технология производства препаратов бактериофагов
1. Подбор активных природных фаговых клонов, не способных

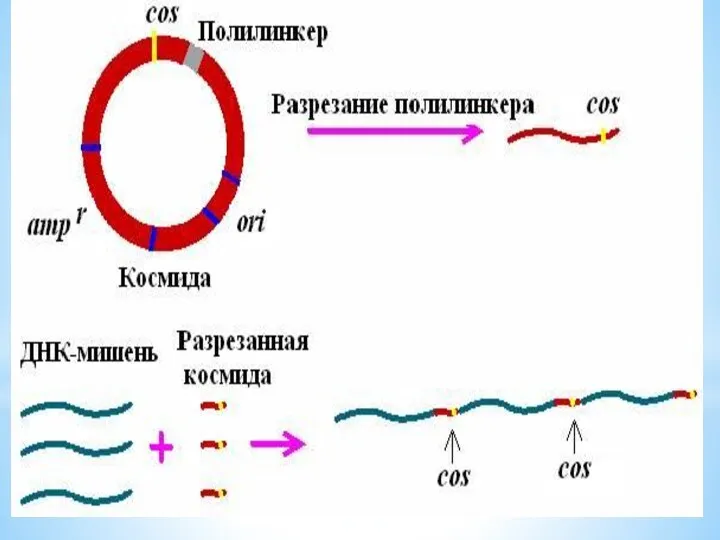
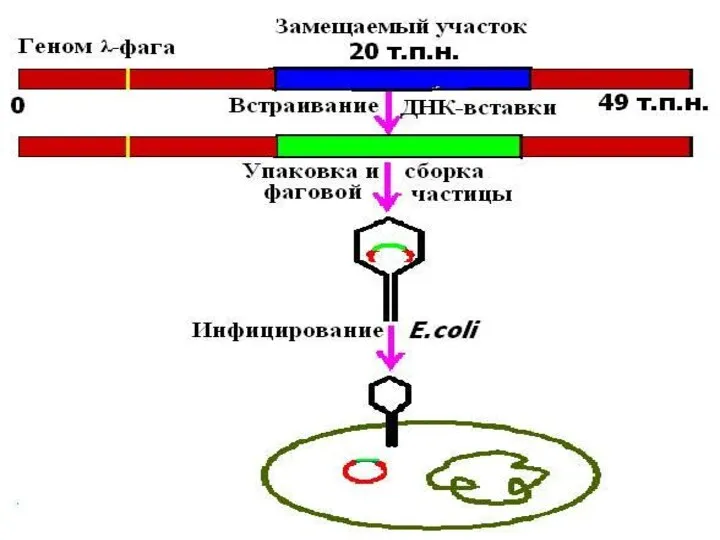
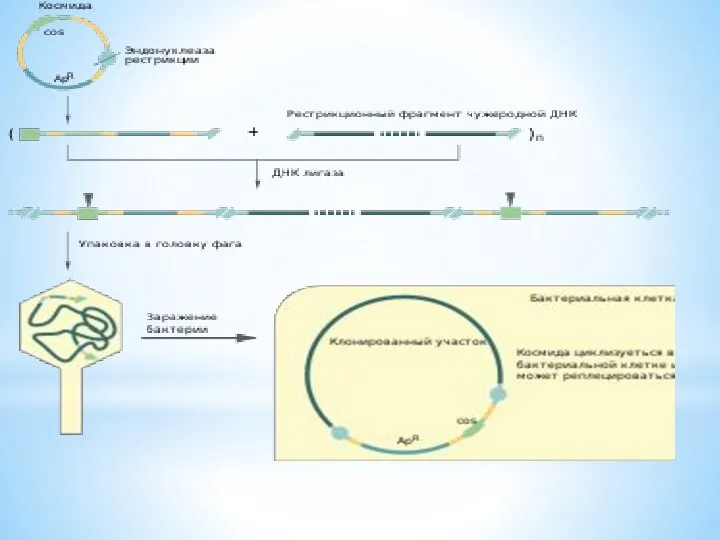
КОСМИДА
cosmid vector - космида.Векторная плазмида , содержащая cos-участок ДНК фага лямбда,
КОСМИДА
cosmid vector - космида.Векторная плазмида
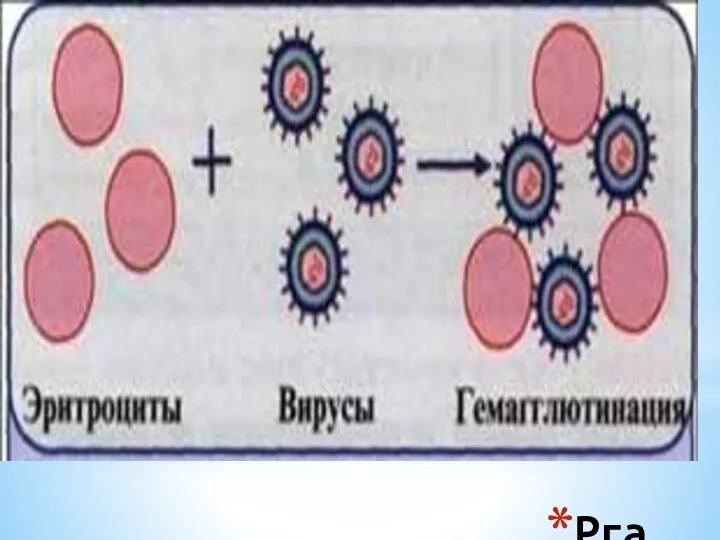
Рга
Рга

РГА
РГА позволяет обнаружить (провести индикацию )вирус:
в зараженном курином эмбрионе
РГА
РГА позволяет обнаружить (провести индикацию )вирус:
в зараженном курином эмбрионе
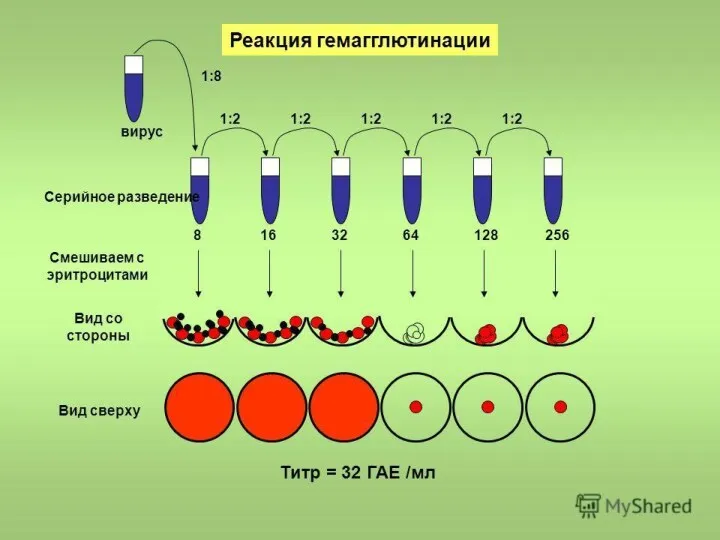

Метод бляшек
Метод бляшек

Вирусы и нанотехнология
Вирионы могут быть реконструированы в пробирке из белка оболочки
Вирусы и нанотехнология
Вирионы могут быть реконструированы в пробирке из белка оболочки

Вирусы и нанотехнология
Белок оболочки бактериофага М13 связали с аморфным фосфатом
Вирусы и нанотехнология
Белок оболочки бактериофага М13 связали с аморфным фосфатом
 Процесс фотосинтеза
Процесс фотосинтеза Микробиология зерновых продуктов
Микробиология зерновых продуктов ПРЕЗЕНТАЦИЯ ДЛЯ ИНТЕРАКТИВНОЙ ДОСКИ. Состав Солнечной системы. 5класс (В пяти частях)
ПРЕЗЕНТАЦИЯ ДЛЯ ИНТЕРАКТИВНОЙ ДОСКИ. Состав Солнечной системы. 5класс (В пяти частях) Анатомия тройничного нерва
Анатомия тройничного нерва Солнце, растение и мы с вами
Солнце, растение и мы с вами Дельфины
Дельфины Поле и его обитатели
Поле и его обитатели Органы цветковых растений.6 класс
Органы цветковых растений.6 класс Кровообращение, лимфообращение
Кровообращение, лимфообращение Орган зрения
Орган зрения Поширення плодів та насіння у природі
Поширення плодів та насіння у природі Ткани растений и животных
Ткани растений и животных Кислородсодержащие органические вещества. Углеводы
Кислородсодержащие органические вещества. Углеводы Влияние факторов внешней среды на онтогенез
Влияние факторов внешней среды на онтогенез Тайская кошка
Тайская кошка Природное и общественное в человеке (Человек как результат биологической и социокультурной эволюции)
Природное и общественное в человеке (Человек как результат биологической и социокультурной эволюции) Паразитические плоские черви. Верно ли утверждение?
Паразитические плоские черви. Верно ли утверждение? Урок-игра Органы цветкового растения
Урок-игра Органы цветкового растения Урок биологии в 5 кл. Красная книга Чувашской Республики
Урок биологии в 5 кл. Красная книга Чувашской Республики Классификация углеводов
Классификация углеводов Атавистическое, нормальное и патологическое поведение собаки и лекарственная коррекция отклоняющегося поведения
Атавистическое, нормальное и патологическое поведение собаки и лекарственная коррекция отклоняющегося поведения Этапы эволюции животного мира
Этапы эволюции животного мира Презентация к уроку Наука биология
Презентация к уроку Наука биология Звери и следы
Звери и следы Общий обзор пищеварительной системы
Общий обзор пищеварительной системы Хвороби бджіл
Хвороби бджіл Кабинет биологии, фото
Кабинет биологии, фото Екологічні групи птахів. Практична робота №3
Екологічні групи птахів. Практична робота №3