- Клеточный цикл и деление клетки

Содержание
- 2. Интерфаза – фаза жизни между делениями клетки и М-фаза – фаза митоза 2 периода жизни клетки
- 3. Компактизация (конденсация) хромосом Формирование веретена и выравнивание хромосом в плоскости экватора клетки Расхождение сестринских хроматид к
- 4. А. фаза М – митотический цикл, который длится обычно 30-60 минут, М-фаза завершается разделением клетки на
- 5. В. Фаза S или синтетическая фаза – период подготовки к митозу, во время которого происходит синтез
- 6. Когда клетки вступают в G1, последовательность молекулярных процессов «обучает» клетку для того, чтобы приступить к вхождению
- 7. Если осуществление клеточного цикла в фазе G1 замедляется, клетка может экспрессировать белки, которые используются не для
- 8. Клетки костного мозгу и слизистой ЖКТ почти непрерывно делятся и редко входят в фазу G0 Биохимические
- 9. Во время мейоза количество хромосом уменьшается в два раза с помощью 2 мейотических делений На 1
- 10. В результате 2-ого мейотического деления появляются дочерние клетки 2 типов: 1-я дочерняя клетка – полярное тельце,
- 11. Фактор стимуляции митоза (МРF) – этот фактор стимулирует клетки, находящиеся в фазе G2, к вступлению в
- 12. Наиболее изучен циклин В, увеличение и снижение концентрации которого точно совпадает с процессами клеточного цикла Белки
- 13. Циклин В включает 2 субъединицы 1. киназный домен, способный к фосфорилированию специфических веществ в клетках 2.
- 14. Синтез циклина В начинается в фазе G1, достигает максимального уровня в конце ранней профазы и резко
- 15. Киназа циклина В фосфорилирует специфические белки, изменяя таким образом их функции и приводя к компактизации хроматина,
- 16. МРF запускает биохимические процессы, в результате которых экспрессируются белки, участвующие в определенных этапах цикла Высокая концентрация
- 17. Циклины содержат специфическую последовательность из 8-10 аминокислот, называемую блоком разрушения Когда белок, содержащий такой участок, должен
- 18. После этого 3-й белок, убиквитинлигаза, присоединяет к остаткам лизина, расположенным вблизи участка блока разрушения, несколько копий
- 19. Белок с прикрепленными остатками убиквитина направляетс к большему РНК-белковому комплексу, называемому протеосомой, в котором находятся протеазы
- 20. Предполагается, что белок, распознающий блок разрушения, активируется путем фосфорилирования Во время накопления МРF (циклина В) фосфатаза
- 21. В результате распознающий белок связывается с блоком разрушения циклина В, приводя таким образом к его разрушению
- 22. Вхождение в митоз представляет собой последовательность фосфорилирования специфических белков циклином В (МРF-киназой) Белковая структура, которая стабилизирует
- 23. В ранней профазе МРF фосфорилирует сериновые и треониновые остатки ламинов и вызывает их деполимеризацию, приводящую к
- 24. Цитокинез блокируется в раннем митозе благодаря тому, что МРF фософрилирует участок легкой цепи миозина, ингибирующий ее
- 25. Итак, комплекс МРF содержит каталитическую киназную субъединицу и регуляторную циклиновую субъединицу. Эти субъединицы являются основными молекулами,
- 26. В клетках млекопитающих существует семейство циклинзависимых киназ (Сdk), которые пронумерованы с 1 по 5 в порядке
- 27. Пока факторы роста стимулируют клетку, Сdk2-циклин D находится в активном состоянии мРНК и белки циклина D
- 28. Сdk2-циклин Е появляется в фазе G1 и достигает максимальной концентрации на границе фаз G1 и S,
- 29. В конце G2-фазы начинается образование комплекса Сdk2-циклина В. Концентрация этого комплекса увеличивается на границе G2 и
- 30. Большинство генов, регулируемых Сdk2-циклинами, делятся на 2 категории: Гены быстрого ответа (кодируют факторы транскрипции, которые участвуют
- 31. А. регуляторная точка фазы G1 – если в фазе G1 обнаруживается повреждение ДНК, белок р53 выступает
- 32. Задержка в фазе G 1 предотвращает копирование поврежденных оснований и тормозит мутацию ДНК. Нестабильный р53 обычно
- 33. Б. регуляторная точка S – фазы функционирует в фазе S, когда реплицируется ДНК. Если произошли ошибки
- 34. 1. изменение одного основания – Депуринизация Замена цитозина на урацил Замена аденина на гипоксантин Алкилирование оснований
- 35. 2. изменение двух оснований Индуцируемое УФ образование димеров тимина Поперечная связь с бифункциональным алкилирующим агентом 3.
- 36. 4. поперечные связи Между основаниями одной нити или двух параллельных нитей Между ДНК и белковыми молекулами
- 37. Существует 2 основных семейства ингибиторов: Семейство р21 Семейство р15 и р16 Белки, ингибирующие комплексы Сdk2-циклин
- 38. Семейство р21 включает 3 белка, кодируемых разными генами: р21, р27, р57 Эти белки содержат гомологичные N-концевые
- 39. р21 ингибирует множество различных комплексов Сdk-циклин, блокируя их взаимодействие с субстратом (экспрессия р21 регулируется с помощью
- 40. Р15 и р16 являются медиаторами задержки пролиферации, вызванной антиростовыми факторами (например, трансформирующий фактор роста- TGF-β) Р15
- 41. Тромбоцитарный фактор роста Эпидермальный фактор роста Фактор роста фибробластов Фактор роста нервов Эритропоэтин М ИЛ-2, ИЛ-3
- 42. Большинство клеток млекопитающих находятся в особой фазе клеточного цикла G0, это удлиненная фаза G1 Гены, кодирующие
- 43. В большинстве клеток гены пролиферации могут быть включены снова; в клетках сердечной мышцы и нейронах Сdk
- 44. Примеры онкогенов
- 45. Гены-онкосупрессоры
- 46. Основным ферментом тирозинкиназного каскада является Ras-белок – небольшая (21кДа), связанная с мембраной ГТФаза, Ras-белок играет роль
- 47. Ras находится в активном состоянии, контактируя с ГТФ, и в неактивном состоянии, контактируя с ГДФ. Активация
- 48. Фактор роста связывается с RPTK-рецептором Димеризация рецептора и самофосфорилирование рецепторных остатков тирозина SH2-домен Grb2-белка связывается с
- 49. Установлено, что Ras инициирует серию реакций, в результате которых активируются другие внутриклеточные киназы, вызывающие клеточное деление
- 50. Активированный Ras взаимодействует с внутриклеточной серин-треонин-киназой Raf-1 (первый компонент ферментного каскада) Активированный Raf-1 активирует другую киназу,
- 51. Далее активируются киназы, контролирующие циклины, и циклинзависимые киназы Ras - Raf-1 – МАР2К Белок МАР2К униклен
- 52. МАРК-семейтсво включает 3 изоформы (МРК, Erk-1, Erk-2), которые могут активироваться в ответ на воздействие многих факторов
- 54. Скачать презентацию

Интерфаза – фаза жизни между делениями клетки и М-фаза – фаза
Интерфаза – фаза жизни между делениями клетки и М-фаза – фаза

Компактизация (конденсация) хромосом
Формирование веретена и выравнивание хромосом в плоскости экватора клетки
Расхождение
Компактизация (конденсация) хромосом
Формирование веретена и выравнивание хромосом в плоскости экватора клетки
Расхождение

А. фаза М – митотический цикл, который длится обычно 30-60 минут,
А. фаза М – митотический цикл, который длится обычно 30-60 минут,

В. Фаза S или синтетическая фаза – период подготовки к митозу,
В. Фаза S или синтетическая фаза – период подготовки к митозу,

Когда клетки вступают в G1, последовательность молекулярных процессов «обучает» клетку для
Когда клетки вступают в G1, последовательность молекулярных процессов «обучает» клетку для

Если осуществление клеточного цикла в фазе G1 замедляется, клетка может экспрессировать
Если осуществление клеточного цикла в фазе G1 замедляется, клетка может экспрессировать

Клетки костного мозгу и слизистой ЖКТ почти непрерывно делятся и редко
Клетки костного мозгу и слизистой ЖКТ почти непрерывно делятся и редко

Во время мейоза количество хромосом уменьшается в два раза с помощью
Во время мейоза количество хромосом уменьшается в два раза с помощью

В результате 2-ого мейотического деления появляются дочерние клетки 2 типов: 1-я
В результате 2-ого мейотического деления появляются дочерние клетки 2 типов: 1-я

Фактор стимуляции митоза (МРF) – этот фактор стимулирует клетки, находящиеся в
Фактор стимуляции митоза (МРF) – этот фактор стимулирует клетки, находящиеся в

Наиболее изучен циклин В, увеличение и снижение концентрации которого точно совпадает
Наиболее изучен циклин В, увеличение и снижение концентрации которого точно совпадает

Циклин В включает 2 субъединицы
1. киназный домен, способный к фосфорилированию специфических
Циклин В включает 2 субъединицы
1. киназный домен, способный к фосфорилированию специфических

Синтез циклина В начинается в фазе G1, достигает максимального уровня в
Синтез циклина В начинается в фазе G1, достигает максимального уровня в

Киназа циклина В фосфорилирует специфические белки, изменяя таким образом их функции
Киназа циклина В фосфорилирует специфические белки, изменяя таким образом их функции

МРF запускает биохимические процессы, в результате которых экспрессируются белки, участвующие в
МРF запускает биохимические процессы, в результате которых экспрессируются белки, участвующие в

Циклины содержат специфическую последовательность из 8-10 аминокислот, называемую блоком разрушения
Когда белок,
Циклины содержат специфическую последовательность из 8-10 аминокислот, называемую блоком разрушения
Когда белок,

После этого 3-й белок, убиквитинлигаза, присоединяет к остаткам лизина, расположенным вблизи
После этого 3-й белок, убиквитинлигаза, присоединяет к остаткам лизина, расположенным вблизи

Белок с прикрепленными остатками убиквитина направляетс к большему РНК-белковому комплексу, называемому
Белок с прикрепленными остатками убиквитина направляетс к большему РНК-белковому комплексу, называемому

Предполагается, что белок, распознающий блок разрушения, активируется путем фосфорилирования
Во время накопления
Предполагается, что белок, распознающий блок разрушения, активируется путем фосфорилирования
Во время накопления

В результате распознающий белок связывается с блоком разрушения циклина В, приводя
В результате распознающий белок связывается с блоком разрушения циклина В, приводя

Вхождение в митоз представляет собой последовательность фосфорилирования специфических белков циклином В
Вхождение в митоз представляет собой последовательность фосфорилирования специфических белков циклином В

В ранней профазе МРF фосфорилирует сериновые и треониновые остатки ламинов и
В ранней профазе МРF фосфорилирует сериновые и треониновые остатки ламинов и

Цитокинез блокируется в раннем митозе благодаря тому, что МРF фософрилирует участок
Цитокинез блокируется в раннем митозе благодаря тому, что МРF фософрилирует участок

Итак, комплекс МРF содержит каталитическую киназную субъединицу и регуляторную циклиновую субъединицу.
Эти
Итак, комплекс МРF содержит каталитическую киназную субъединицу и регуляторную циклиновую субъединицу.
Эти

В клетках млекопитающих существует семейство циклинзависимых киназ (Сdk), которые пронумерованы с
В клетках млекопитающих существует семейство циклинзависимых киназ (Сdk), которые пронумерованы с

Пока факторы роста стимулируют клетку, Сdk2-циклин D находится в активном состоянии
мРНК
Пока факторы роста стимулируют клетку, Сdk2-циклин D находится в активном состоянии
мРНК

Сdk2-циклин Е появляется в фазе G1 и достигает максимальной концентрации на
Сdk2-циклин Е появляется в фазе G1 и достигает максимальной концентрации на

В конце G2-фазы начинается образование комплекса Сdk2-циклина В. Концентрация этого комплекса
В конце G2-фазы начинается образование комплекса Сdk2-циклина В. Концентрация этого комплекса

Большинство генов, регулируемых Сdk2-циклинами, делятся на 2 категории:
Гены быстрого ответа (кодируют
Большинство генов, регулируемых Сdk2-циклинами, делятся на 2 категории:
Гены быстрого ответа (кодируют

А. регуляторная точка фазы G1 – если в фазе G1 обнаруживается
А. регуляторная точка фазы G1 – если в фазе G1 обнаруживается

Задержка в фазе G 1 предотвращает копирование поврежденных оснований и тормозит
Задержка в фазе G 1 предотвращает копирование поврежденных оснований и тормозит

Б. регуляторная точка S – фазы функционирует в фазе S, когда
Б. регуляторная точка S – фазы функционирует в фазе S, когда

1. изменение одного основания –
Депуринизация
Замена цитозина на урацил
Замена аденина на
1. изменение одного основания –
Депуринизация
Замена цитозина на урацил
Замена аденина на

2. изменение двух оснований
Индуцируемое УФ образование димеров тимина
Поперечная связь с бифункциональным
2. изменение двух оснований
Индуцируемое УФ образование димеров тимина
Поперечная связь с бифункциональным

4. поперечные связи
Между основаниями одной нити или двух параллельных нитей
Между ДНК
4. поперечные связи
Между основаниями одной нити или двух параллельных нитей
Между ДНК

Существует 2 основных семейства ингибиторов:
Семейство р21
Семейство р15 и р16
Белки, ингибирующие комплексы
Существует 2 основных семейства ингибиторов:
Семейство р21
Семейство р15 и р16
Белки, ингибирующие комплексы

Семейство р21 включает 3 белка, кодируемых разными генами: р21, р27, р57
Эти
Семейство р21 включает 3 белка, кодируемых разными генами: р21, р27, р57
Эти

р21 ингибирует множество различных комплексов Сdk-циклин, блокируя их взаимодействие с субстратом
р21 ингибирует множество различных комплексов Сdk-циклин, блокируя их взаимодействие с субстратом

Р15 и р16 являются медиаторами задержки пролиферации, вызванной антиростовыми факторами (например,
Р15 и р16 являются медиаторами задержки пролиферации, вызванной антиростовыми факторами (например,

Тромбоцитарный фактор роста
Эпидермальный фактор роста
Фактор роста фибробластов
Фактор роста нервов
Эритропоэтин
М
ИЛ-2, ИЛ-3 и
Тромбоцитарный фактор роста
Эпидермальный фактор роста
Фактор роста фибробластов
Фактор роста нервов
Эритропоэтин
М
ИЛ-2, ИЛ-3 и

Большинство клеток млекопитающих находятся в особой фазе клеточного цикла G0, это
Большинство клеток млекопитающих находятся в особой фазе клеточного цикла G0, это

В большинстве клеток гены пролиферации могут быть включены снова; в клетках
В большинстве клеток гены пролиферации могут быть включены снова; в клетках

Примеры онкогенов
Примеры онкогенов

Гены-онкосупрессоры
Гены-онкосупрессоры

Основным ферментом тирозинкиназного каскада является Ras-белок – небольшая (21кДа), связанная с
Основным ферментом тирозинкиназного каскада является Ras-белок – небольшая (21кДа), связанная с

Ras находится в активном состоянии, контактируя с ГТФ, и в неактивном
Ras находится в активном состоянии, контактируя с ГТФ, и в неактивном

Фактор роста связывается с RPTK-рецептором
Димеризация рецептора и самофосфорилирование рецепторных остатков тирозина
SH2-домен
Фактор роста связывается с RPTK-рецептором
Димеризация рецептора и самофосфорилирование рецепторных остатков тирозина
SH2-домен

Установлено, что Ras инициирует серию реакций, в результате которых активируются другие
Установлено, что Ras инициирует серию реакций, в результате которых активируются другие

Активированный Ras взаимодействует с внутриклеточной серин-треонин-киназой Raf-1 (первый компонент ферментного каскада)
Активированный
Активированный Ras взаимодействует с внутриклеточной серин-треонин-киназой Raf-1 (первый компонент ферментного каскада)
Активированный

Далее активируются киназы, контролирующие циклины, и циклинзависимые киназы
Ras - Raf-1 –
Далее активируются киназы, контролирующие циклины, и циклинзависимые киназы
Ras - Raf-1 –

МАРК-семейтсво включает 3 изоформы (МРК, Erk-1, Erk-2), которые могут активироваться в
МАРК-семейтсво включает 3 изоформы (МРК, Erk-1, Erk-2), которые могут активироваться в
 Соединительная ткань и ее виды
Соединительная ткань и ее виды Моллюски: общая характеристика типа
Моллюски: общая характеристика типа Учение об анализаторах. Орган зрения
Учение об анализаторах. Орган зрения Цветок адениум
Цветок адениум Патрин А
Патрин А Задания линии 2 и 22 по экспериментам в КИМ ЕГЭ по биологии 2022
Задания линии 2 и 22 по экспериментам в КИМ ЕГЭ по биологии 2022 Лимфатическая система
Лимфатическая система Психология и социобиология половых различий
Психология и социобиология половых различий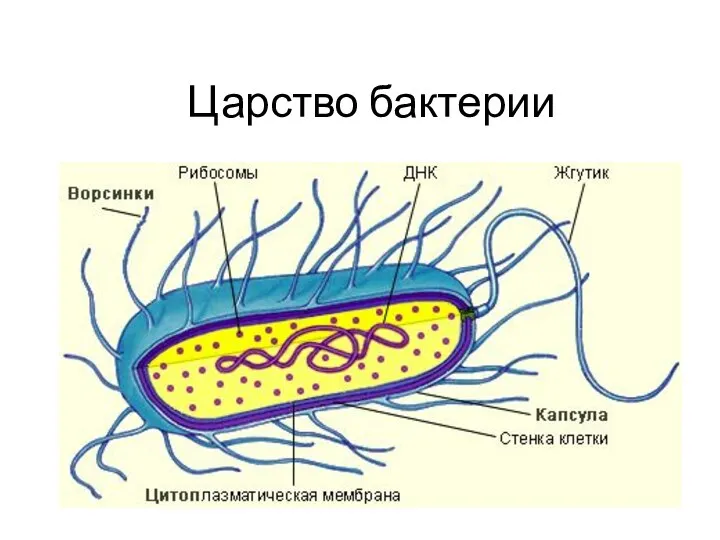 Презентация ц.бактерии 6кл
Презентация ц.бактерии 6кл Опасное растение борщевик
Опасное растение борщевик Внутривидовые и межвидовые взаимодействия у грибов, водорослей, высших растений. (Часть 4)
Внутривидовые и межвидовые взаимодействия у грибов, водорослей, высших растений. (Часть 4) Главные направления эволюции органического мира
Главные направления эволюции органического мира Тварина Південної Америки - вікунья
Тварина Південної Америки - вікунья Презентация по биологии на тему Класс Пресмыкающиеся. Особенности внешнего и внутреннего строения. Многообразие. Происхождение
Презентация по биологии на тему Класс Пресмыкающиеся. Особенности внешнего и внутреннего строения. Многообразие. Происхождение Черепные нервы. Обонятельные нервы
Черепные нервы. Обонятельные нервы Дезоксирибонуклеиновая кислота (ДНК)
Дезоксирибонуклеиновая кислота (ДНК) Строение клетки. Ткани
Строение клетки. Ткани Красная книга Белгородской области
Красная книга Белгородской области Физиология вегетативной нервной системы
Физиология вегетативной нервной системы Види сполучних тканин
Види сполучних тканин Эволюционная теория Ч. Дарвина
Эволюционная теория Ч. Дарвина Методы изучения наследственности человека
Методы изучения наследственности человека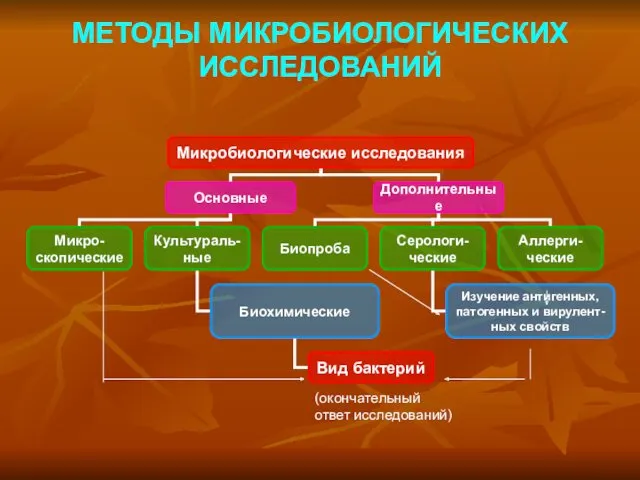 Методы микробиологических исследований
Методы микробиологических исследований Мужская половая система
Мужская половая система Действия и взаимодействия в системе Я и другой: нейробиологические аспекты
Действия и взаимодействия в системе Я и другой: нейробиологические аспекты Электрические свойства тканей организма. (Лекция 6)
Электрические свойства тканей организма. (Лекция 6) Кожа – наружный покровный орган. 8 класс
Кожа – наружный покровный орган. 8 класс В мире животных
В мире животных