Липидтердің алмасуы Липидтердің анаболитикалық және катаболитикалық жолдармен пайда болуы және биологиялық рөлі презентация
- Липидтердің алмасуы Липидтердің анаболитикалық және катаболитикалық жолдармен пайда болуы және биологиялық рөлі

Содержание
- 2. Липидтердің алмасуы және функциясы. липидтер күрделі (липидтер) қарапайым стеридтар холестеридтар триацилглицериндер гликолипидтар цереброзидтер ганглиозидтар сфингофосфатиттар фосфоглицериндар
- 3. Липидтердің биологиялық құрылымы: Липидтер энергия беретін материал болып табылады яғни,қауіпсіздік,пластикалық,тасымалдаушы және реттеуші қызмет атқарады. Липидтерді физиологиялық
- 4. Қаныққан май қышқылдары -пальмитин(С15Н31СООН) -стеарин ( С17Н35СООН) -арахин (С19Н39СООН). Мононенасыщенные( одна двойная связь): -олеин(С17Н35СООН) -кротон(С3Н5СООН) -пальмитоолеин(С15Н29СООН)
- 5. Насыщенные -пальмитиновая(С15Н31СООН) -стеариновая ( С17Н35СООН) -арахиновая (С19Н39СООН). Мононенасыщенные( одна двойная связь): -олеиновая(С17Н35СООН) -кротоновая(С3Н5СООН) -пальмитоолеиновая(С15Н29СООН)
- 6. Полиқанықпаған май қышқылдары- 2- және одан көп қосарланған байланыстармен: -линоль(С17Н31СООН)- 2 = байланыс -линолен(С17Н29СООН)- 3= связи
- 7. Тасымалдаушы липидтер. Тасымалдаушы липидтер-белоктық комплекстің құрамында болатын экстрацелюлярлық сұйықтық (қан,лимфа) арқылы бір-органнан екінші органга жеткізу қызметін
- 8. Хиломикрондар (ХМ) Хиломикрондар (ХМ) ішек қабырғасында түзіледі,басқа липопротеиндардан айырмашылығы құрамында өте жоғары дәрежеде триацилглицериндер (85-90%), өте
- 9. ӨТТЛП немесе пре-β-липопротеиндер ӨТТЛП немесе пре-β-липопротеиндер, бауырда түзіледі кей жағдайда аш ішектің эпителиальдық жасушасында түзілуі мүмкін.
- 10. ТТЛП немесе β- липопротеиндар. ТТЛП или β- липопротеиндар-қанда ӨТТЛП дан тұзіледі,және барлық организімдегі ондағы жасушалардағы,бауырдағы холестериннің
- 11. ЖТЛП немесе α-липопротеиндер ЖТЛПнемесе α-липопротеиндер,гепатоциттерде қалыптасып,жоғарғы дәрежедегі фосфолипидтер (25-27%), холестеридтер ( 17-20 %) ақуыздармен ( 45-49%)сипатталады,және
- 12. Липидтердің қорытылып сіңіуі. Липидтердің құрамымен әртүрлігі пайдаланып отырған өнімге байланысты. Адам тамақпен бірге бір-бірінен май қышқылдары
- 13. Өт қышқылдары. Майлардың дұрыстап қорытылуындағы ең маңызды шар өт қышқылдары әсерінен эмулгіленген күйге түсуі. Өт қышқылдары
- 17. Триацилглицеридтердің қортылуы.
- 18. CH2-O-CO-CH2-R\ CH-O-CO-CH2-R\\ CH2-O-CO-CH2-R\\\ +3HOH липаза CH2-OH CH-OH CH2OH триацилглицерид глицерин + R\-CH2-COOH R\\-CH2-COOH R\\\-CH2-COOH Май қышқылы
- 19. Май ұлпасындағы липид синтезі(липидтердің корға сақталуы)
- 20. Липидтер мобилизациясы.
- 21. Карнитинді челнок Карнитин амин қышқылдарын цитозольден митохондрияға тасымалдауы үшін керек. Ол жерде ацил-КоА бета-тотығуға ұшырайды.
- 22. Май қышқылдарының бета-тотығуы
- 23. Кейін екі көміртек атомына қысқарған ацил-КоА молекуласының тотығуы жүреді
- 24. Бета-тотығу процесінің биоэнергетикасы Амин қышқылдары тотығуының биоэнергетикасы бета-тотығу циклында туындаған молекулалар мен әр циклда синтезделетін АТФ
- 25. Май қышқылдарының биосинтезі Синтездің қайнар көзі ретінде ацетил-КоАдан түзілетін малонил-КоА болып табылады. Ацетил-КоА амин қышқылдары бета-тотығуы
- 27. Тема №7: Липидтер амасуы Негізгі сұрақтар: 1.Стероидтар алмасуы. Холестерин биосинтезі туралы түсінік. 2.Өт қышқылы синтезі. Организмнен
- 28. Холестерин биосинтезі Стероидтарға көмірсу туындысы цклопентан-пергидро-фенантрена жатады. 13 күнде(эстран) немесе 10 және 13 күйде(андростан) метилденген. Стероидтардың
- 30. Холестерин синтезінің негізгі қайнар көзі ретінде ацетил-КоА болып таблады. Бета-гидрокси-бета-метилглутарил-КоА түзілгенге дейін синтез осы жолмен жүреді.
- 31. Мевалонат киназа әсерінен 2АТФ молекулалары көмегімен мевалонилпирофосфат түзіледі. Оның декарбоксилденуі кезінде изопентилпирофосфат және диметилаллилпирофосфат түзіледі:
- 33. Фосфолипидтер мен гликолипидтер синтезі Фосфолипидтер мен гликолипидтер-жасуша мембранасының ең негізгі құрылымдық компоненттері. Олардың синтезі үшін әдеттегі
- 34. Холин синтезі кезінде метионин метилді топтардың донаторы рөлін атқарады. Метионин S-аденозилметионина түрінде жасушаларда кездеседі:
- 35. Коламин коламин фосфокиназа әсерінен, активтеліп, коламин фосфат түзеді.
- 36. Ұқсас болып фосфохолин түзіледі. Ол лецитиндер синтезі үшін қажет.
- 37. Фосфохолин түзілу жолы:
- 38. Гликолипидтер синтезі Гликолипидтер синтезі эндоплазматикалық ретикулумның мембранасында жүреді. Түзілген гликолипидтер мембрана беткейінде орналасады. Кейін Гольджи аппаратына
- 40. Сфингозин-ацитрансфераза фермент әсерінен сфингозинге пептидтік байланыспен аминқышқылы келіп жалғасады. Нәтижесінде церамид пайда болады.
- 41. Церамидке фосфохолин келіп қосылады. Одан сфингозинфосфатид сфингомиелин түзіледі.
- 42. Гликолипидтер синтезі Гликолипидтер синтезі церомидтер арқылы жүзеге асады. Цереброзидтер синтезі кезінде церамидке гликозидтік тізбек арқылы моносахаридтер
- 44. Скачать презентацию
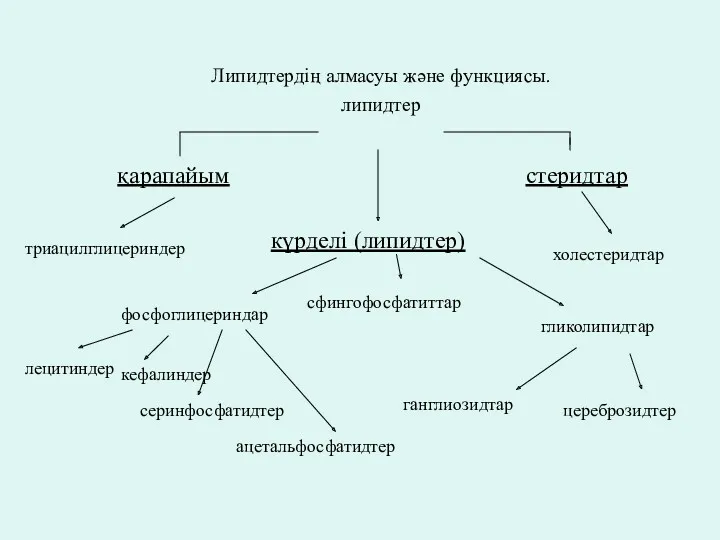
Липидтердің алмасуы және функциясы.
липидтер
күрделі (липидтер)
қарапайым
стеридтар
холестеридтар
триацилглицериндер
гликолипидтар
цереброзидтер
ганглиозидтар
сфингофосфатиттар
фосфоглицериндар
лецитиндер
кефалиндер
серинфосфатидтер
ацетальфосфатидтер
Липидтердің алмасуы және функциясы.
липидтер
күрделі (липидтер)
қарапайым
стеридтар
холестеридтар
триацилглицериндер
гликолипидтар
цереброзидтер
ганглиозидтар
сфингофосфатиттар
фосфоглицериндар
лецитиндер
кефалиндер
серинфосфатидтер
ацетальфосфатидтер

Липидтердің биологиялық құрылымы:
Липидтер энергия беретін материал болып табылады яғни,қауіпсіздік,пластикалық,тасымалдаушы және реттеуші
Липидтердің биологиялық құрылымы:
Липидтер энергия беретін материал болып табылады яғни,қауіпсіздік,пластикалық,тасымалдаушы және реттеуші

Қаныққан май қышқылдары
-пальмитин(С15Н31СООН)
-стеарин ( С17Н35СООН)
-арахин (С19Н39СООН).
Мононенасыщенные( одна двойная
Қаныққан май қышқылдары
-пальмитин(С15Н31СООН)
-стеарин ( С17Н35СООН)
-арахин (С19Н39СООН).
Мононенасыщенные( одна двойная

Насыщенные
-пальмитиновая(С15Н31СООН)
-стеариновая ( С17Н35СООН)
-арахиновая (С19Н39СООН).
Мононенасыщенные( одна двойная связь):
-олеиновая(С17Н35СООН)
-кротоновая(С3Н5СООН)
-пальмитоолеиновая(С15Н29СООН)
Насыщенные
-пальмитиновая(С15Н31СООН)
-стеариновая ( С17Н35СООН)
-арахиновая (С19Н39СООН).
Мононенасыщенные( одна двойная связь):
-олеиновая(С17Н35СООН)
-кротоновая(С3Н5СООН)
-пальмитоолеиновая(С15Н29СООН)

Полиқанықпаған май қышқылдары- 2- және одан көп қосарланған байланыстармен:
-линоль(С17Н31СООН)- 2
Полиқанықпаған май қышқылдары- 2- және одан көп қосарланған байланыстармен:
-линоль(С17Н31СООН)- 2

Тасымалдаушы липидтер.
Тасымалдаушы липидтер-белоктық комплекстің құрамында болатын экстрацелюлярлық сұйықтық (қан,лимфа) арқылы бір-органнан
Тасымалдаушы липидтер.
Тасымалдаушы липидтер-белоктық комплекстің құрамында болатын экстрацелюлярлық сұйықтық (қан,лимфа) арқылы бір-органнан

Хиломикрондар (ХМ)
Хиломикрондар (ХМ) ішек қабырғасында түзіледі,басқа липопротеиндардан айырмашылығы құрамында өте жоғары
Хиломикрондар (ХМ)
Хиломикрондар (ХМ) ішек қабырғасында түзіледі,басқа липопротеиндардан айырмашылығы құрамында өте жоғары

ӨТТЛП немесе пре-β-липопротеиндер
ӨТТЛП немесе пре-β-липопротеиндер, бауырда түзіледі кей жағдайда аш ішектің
ӨТТЛП немесе пре-β-липопротеиндер
ӨТТЛП немесе пре-β-липопротеиндер, бауырда түзіледі кей жағдайда аш ішектің

ТТЛП немесе β- липопротеиндар.
ТТЛП или β- липопротеиндар-қанда ӨТТЛП дан тұзіледі,және барлық
ТТЛП немесе β- липопротеиндар.
ТТЛП или β- липопротеиндар-қанда ӨТТЛП дан тұзіледі,және барлық

ЖТЛП немесе α-липопротеиндер
ЖТЛПнемесе α-липопротеиндер,гепатоциттерде қалыптасып,жоғарғы дәрежедегі фосфолипидтер (25-27%), холестеридтер ( 17-20
ЖТЛП немесе α-липопротеиндер
ЖТЛПнемесе α-липопротеиндер,гепатоциттерде қалыптасып,жоғарғы дәрежедегі фосфолипидтер (25-27%), холестеридтер ( 17-20

Липидтердің қорытылып сіңіуі.
Липидтердің құрамымен әртүрлігі пайдаланып отырған өнімге байланысты.
Адам тамақпен бірге
Липидтердің қорытылып сіңіуі.
Липидтердің құрамымен әртүрлігі пайдаланып отырған өнімге байланысты.
Адам тамақпен бірге

Өт қышқылдары.
Майлардың дұрыстап қорытылуындағы ең маңызды шар өт қышқылдары әсерінен эмулгіленген
Өт қышқылдары.
Майлардың дұрыстап қорытылуындағы ең маңызды шар өт қышқылдары әсерінен эмулгіленген
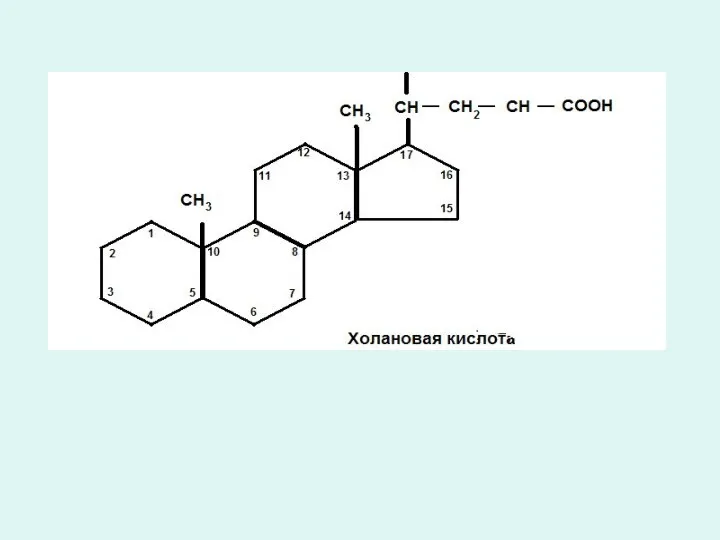
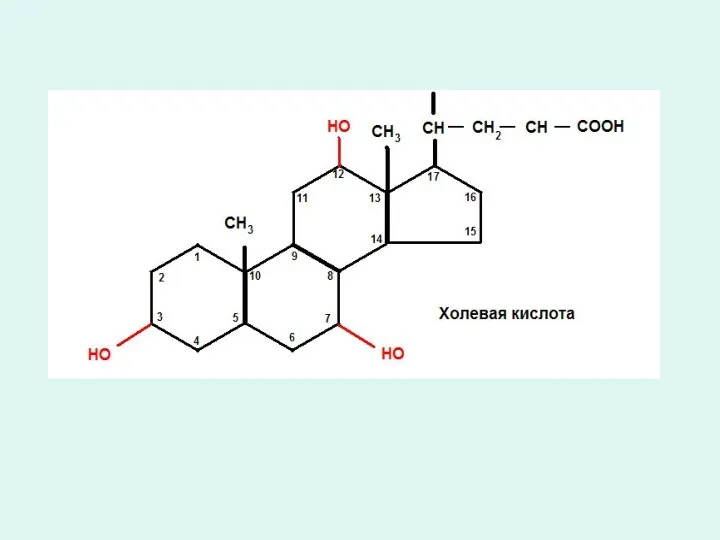
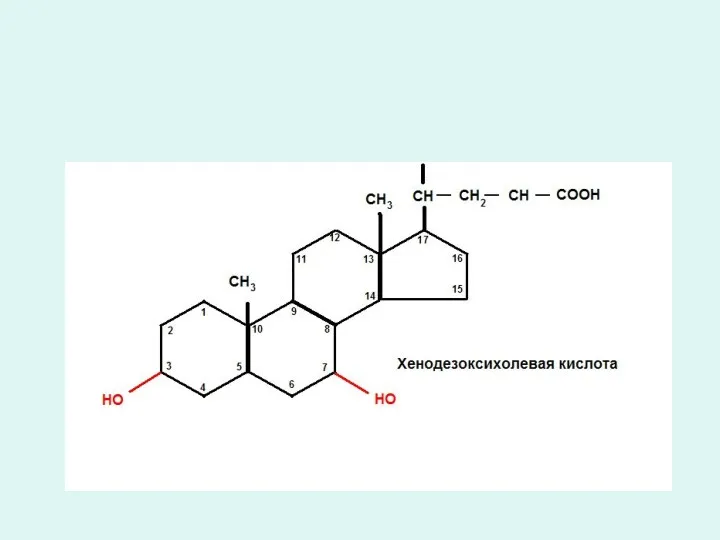

Триацилглицеридтердің қортылуы.
Триацилглицеридтердің қортылуы.
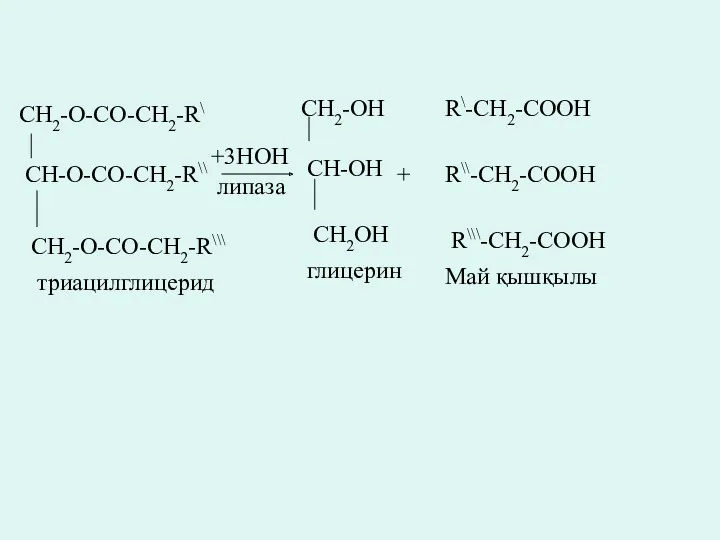
CH2-O-CO-CH2-R\
CH-O-CO-CH2-R\\
CH2-O-CO-CH2-R\\\
+3HOH
липаза
CH2-OH
CH-OH
CH2OH
триацилглицерид
глицерин
+
R\-CH2-COOH
R\\-CH2-COOH
R\\\-CH2-COOH
Май қышқылы
CH2-O-CO-CH2-R\
CH-O-CO-CH2-R\\
CH2-O-CO-CH2-R\\\
+3HOH
липаза
CH2-OH
CH-OH
CH2OH
триацилглицерид
глицерин
+
R\-CH2-COOH
R\\-CH2-COOH
R\\\-CH2-COOH
Май қышқылы

Май ұлпасындағы липид синтезі(липидтердің корға сақталуы)
Май ұлпасындағы липид синтезі(липидтердің корға сақталуы)
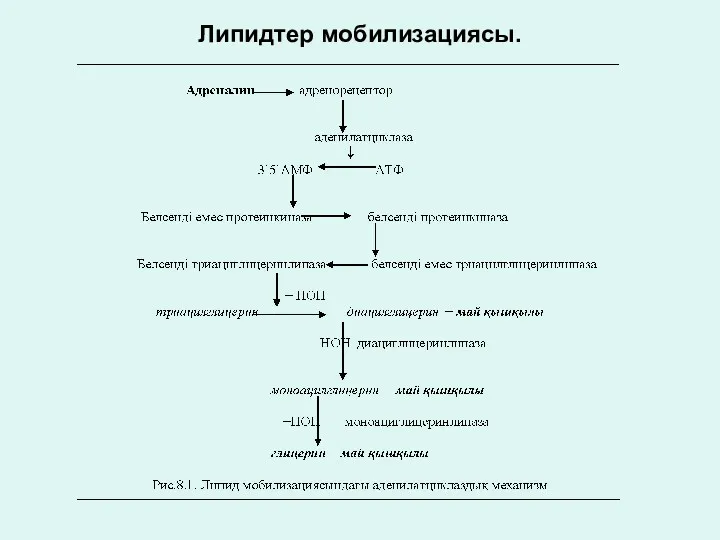
Липидтер мобилизациясы.
Липидтер мобилизациясы.

Карнитинді челнок
Карнитин амин қышқылдарын цитозольден митохондрияға тасымалдауы үшін керек. Ол жерде
Карнитинді челнок
Карнитин амин қышқылдарын цитозольден митохондрияға тасымалдауы үшін керек. Ол жерде

Май қышқылдарының бета-тотығуы
Май қышқылдарының бета-тотығуы

Кейін екі көміртек атомына қысқарған ацил-КоА молекуласының тотығуы жүреді
Кейін екі көміртек атомына қысқарған ацил-КоА молекуласының тотығуы жүреді

Бета-тотығу процесінің биоэнергетикасы
Амин қышқылдары тотығуының биоэнергетикасы бета-тотығу циклында туындаған молекулалар мен
Бета-тотығу процесінің биоэнергетикасы
Амин қышқылдары тотығуының биоэнергетикасы бета-тотығу циклында туындаған молекулалар мен

Май қышқылдарының биосинтезі
Синтездің қайнар көзі ретінде ацетил-КоАдан түзілетін малонил-КоА болып табылады.
Май қышқылдарының биосинтезі
Синтездің қайнар көзі ретінде ацетил-КоАдан түзілетін малонил-КоА болып табылады.

Тема №7: Липидтер амасуы
Негізгі сұрақтар:
1.Стероидтар алмасуы. Холестерин биосинтезі туралы түсінік.
Тема №7: Липидтер амасуы
Негізгі сұрақтар:
1.Стероидтар алмасуы. Холестерин биосинтезі туралы түсінік.

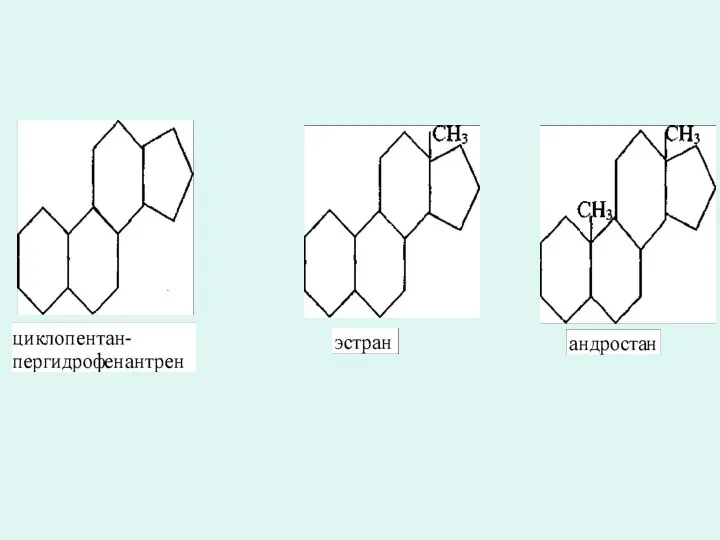
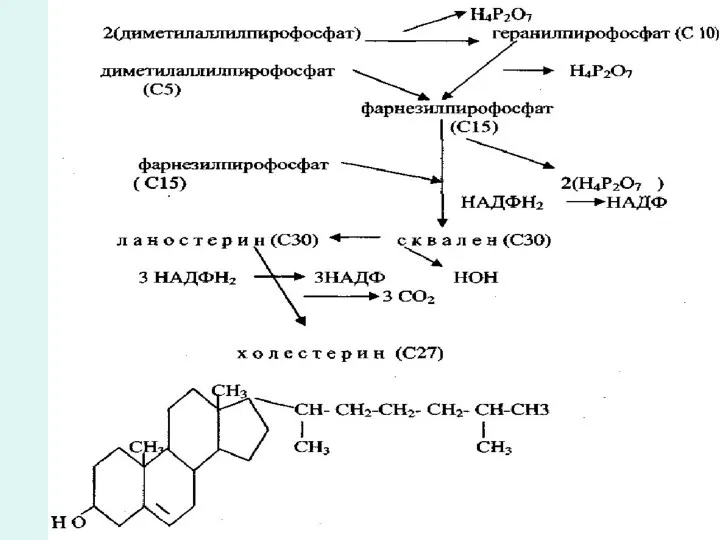
Холестерин биосинтезі
Стероидтарға көмірсу туындысы цклопентан-пергидро-фенантрена жатады. 13 күнде(эстран) немесе 10 және
Холестерин биосинтезі
Стероидтарға көмірсу туындысы цклопентан-пергидро-фенантрена жатады. 13 күнде(эстран) немесе 10 және
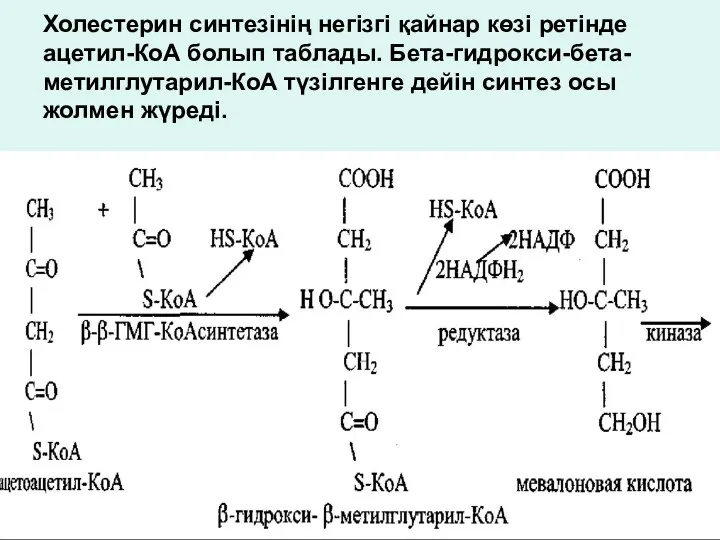
Холестерин синтезінің негізгі қайнар көзі ретінде ацетил-КоА болып таблады. Бета-гидрокси-бета-метилглутарил-КоА түзілгенге
Холестерин синтезінің негізгі қайнар көзі ретінде ацетил-КоА болып таблады. Бета-гидрокси-бета-метилглутарил-КоА түзілгенге
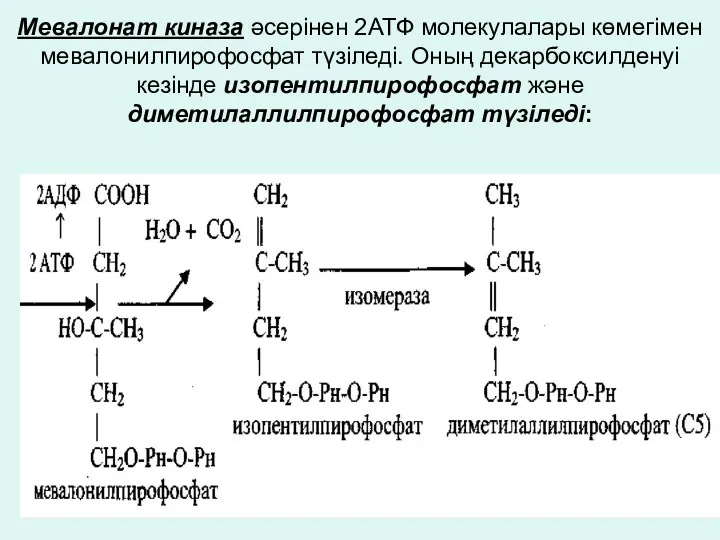
Мевалонат киназа әсерінен 2АТФ молекулалары көмегімен мевалонилпирофосфат түзіледі. Оның декарбоксилденуі кезінде
Мевалонат киназа әсерінен 2АТФ молекулалары көмегімен мевалонилпирофосфат түзіледі. Оның декарбоксилденуі кезінде

Фосфолипидтер мен гликолипидтер синтезі
Фосфолипидтер мен гликолипидтер-жасуша мембранасының ең негізгі құрылымдық компоненттері.
Фосфолипидтер мен гликолипидтер синтезі
Фосфолипидтер мен гликолипидтер-жасуша мембранасының ең негізгі құрылымдық компоненттері.

Холин синтезі кезінде метионин метилді топтардың донаторы рөлін атқарады. Метионин S-аденозилметионина
Холин синтезі кезінде метионин метилді топтардың донаторы рөлін атқарады. Метионин S-аденозилметионина
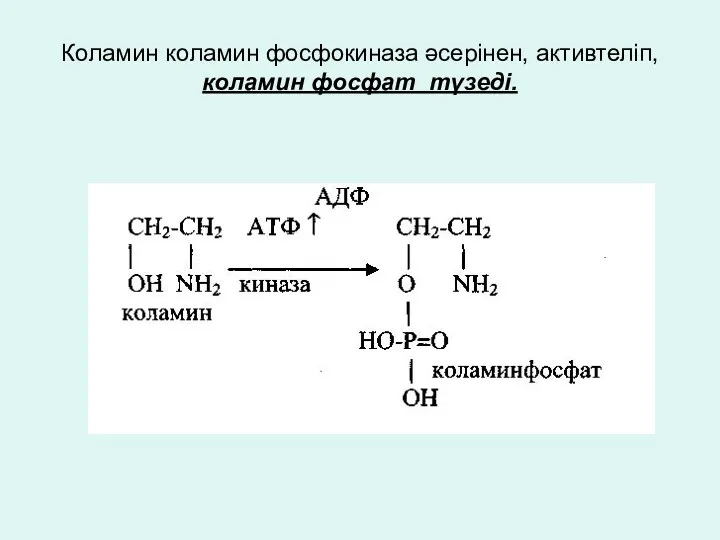
Коламин коламин фосфокиназа әсерінен, активтеліп, коламин фосфат түзеді.
Коламин коламин фосфокиназа әсерінен, активтеліп, коламин фосфат түзеді.
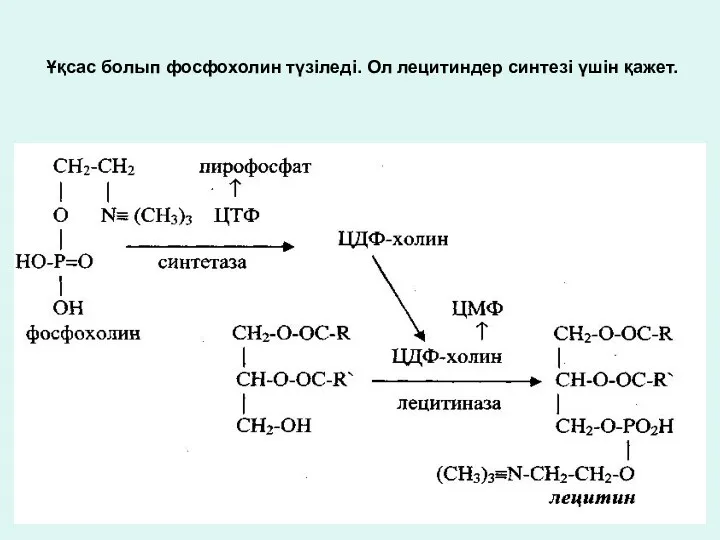
Ұқсас болып фосфохолин түзіледі. Ол лецитиндер синтезі үшін қажет.
Ұқсас болып фосфохолин түзіледі. Ол лецитиндер синтезі үшін қажет.
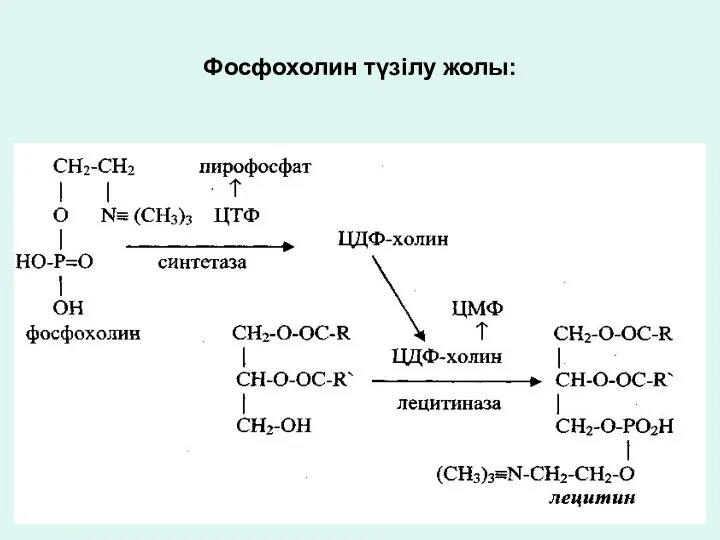
Фосфохолин түзілу жолы:
Фосфохолин түзілу жолы:

Гликолипидтер синтезі
Гликолипидтер синтезі эндоплазматикалық ретикулумның мембранасында жүреді.
Түзілген гликолипидтер мембрана беткейінде
Гликолипидтер синтезі
Гликолипидтер синтезі эндоплазматикалық ретикулумның мембранасында жүреді.
Түзілген гликолипидтер мембрана беткейінде
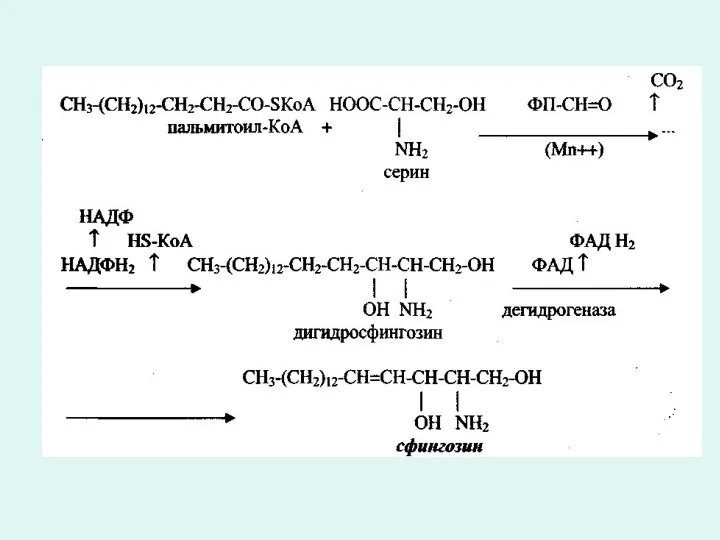
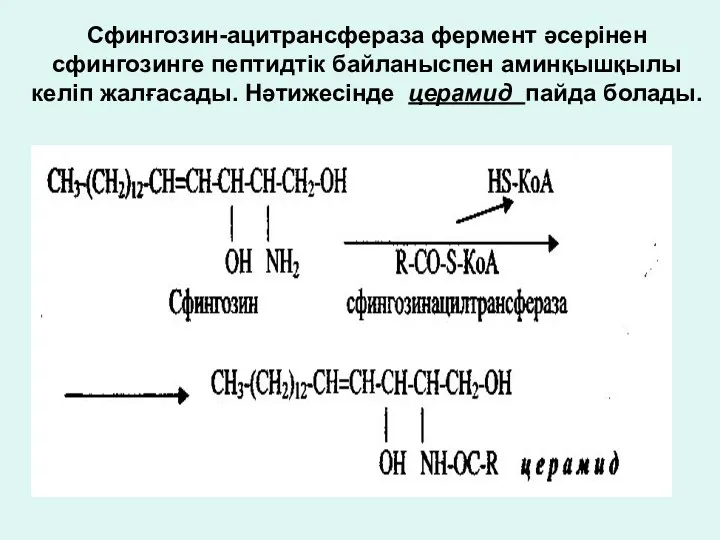
Сфингозин-ацитрансфераза фермент әсерінен сфингозинге пептидтік байланыспен аминқышқылы келіп жалғасады. Нәтижесінде церамид
Сфингозин-ацитрансфераза фермент әсерінен сфингозинге пептидтік байланыспен аминқышқылы келіп жалғасады. Нәтижесінде церамид
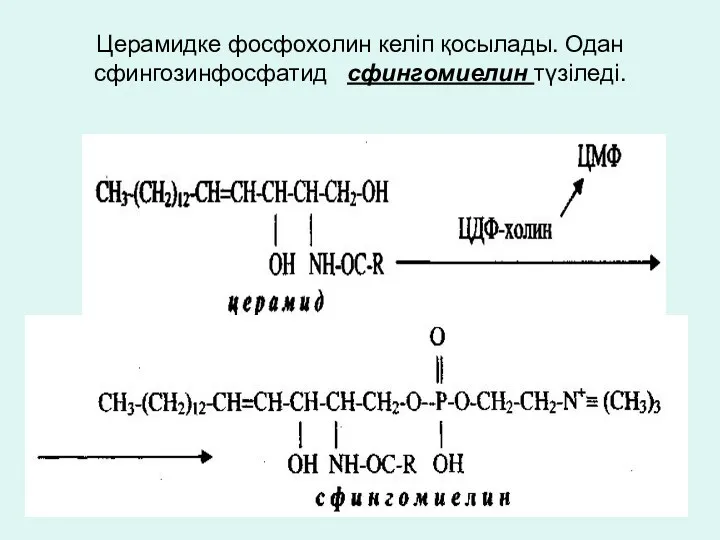
Церамидке фосфохолин келіп қосылады. Одан сфингозинфосфатид сфингомиелин түзіледі.
Церамидке фосфохолин келіп қосылады. Одан сфингозинфосфатид сфингомиелин түзіледі.

Гликолипидтер синтезі
Гликолипидтер синтезі церомидтер арқылы жүзеге асады.
Цереброзидтер синтезі кезінде церамидке
Гликолипидтер синтезі
Гликолипидтер синтезі церомидтер арқылы жүзеге асады.
Цереброзидтер синтезі кезінде церамидке
 Презентация к уроку биологии 9 класс на тему: Система органов пищеварения
Презентация к уроку биологии 9 класс на тему: Система органов пищеварения Презентации к урокам
Презентации к урокам Царство грибов
Царство грибов Размножение живых организмов
Размножение живых организмов Общая характеристика бактерий и грибов, их строение, жизнедеятельность и многообразие
Общая характеристика бактерий и грибов, их строение, жизнедеятельность и многообразие Растения в домашней аптечке
Растения в домашней аптечке Эндокринная система рыб. Лекция 3
Эндокринная система рыб. Лекция 3 Новый вид симбиоза
Новый вид симбиоза Вид. Структура и критерии вида
Вид. Структура и критерии вида Internal medium of organism (Blood)
Internal medium of organism (Blood) Эволюция органического мира
Эволюция органического мира Выступление на всероссийском семинаре Личностно- ориентированнный подход при создании проектных работ.
Выступление на всероссийском семинаре Личностно- ориентированнный подход при создании проектных работ. Селекция яблок
Селекция яблок Хризантема овощная
Хризантема овощная Пищеварение в кишечнике
Пищеварение в кишечнике История первобытного общества: социогенез
История первобытного общества: социогенез Определение активной реакции воды (рН)
Определение активной реакции воды (рН) Систематика
Систематика Помидор – полезный овощ. 2 класс
Помидор – полезный овощ. 2 класс Физиология дыхания
Физиология дыхания Біологічно активні речовини
Біологічно активні речовини Генная регуляция поддержания популяции стволовых клеток в организме млекопитающих
Генная регуляция поддержания популяции стволовых клеток в организме млекопитающих Состав и функции крови
Состав и функции крови Не ходите дети в Африку гулять... или самые необычные растения Африки
Не ходите дети в Африку гулять... или самые необычные растения Африки Стати собак
Стати собак Класс однодольные
Класс однодольные Царство Грибы
Царство Грибы Bacillus. General Characteristics of Bacillus
Bacillus. General Characteristics of Bacillus