- Механизм действия ферментов. Этапы ферментативного катализа

Содержание
- 2. Выдвинутая в 1913 году Л. Михаэлисом и М. Ментен общая теория ферментативного катализа постулировала, что фермент
- 3. Структурно-функциональная организация ферментов. Схема Активные центры Якорные площадки Субстратный субстрат продукт Центры регуляции + и -
- 4. Структурно-функциональная организация ферментов. Активный (субстратный) центр - это совокупность функциональных групп, расположенных в разных участках ПП
- 5. Активный центр включает в себя: Каталитический участок или центр, непосредственно взаимодействующий с субстратом, осуществляющий катализ. Контактная,
- 6. 1 этап: постепенное «причаливание» S к «якорной» площадке F. 2 этап: напряжение и деформация: индуцированное соответствие
- 7. Химические связи, действующие при этом: 1. Силы Ван дер Ваальса 2. Электростатическое взаимодействие 3. Водородные связи
- 8. Якорный участок трипсина представлен длинным узким карманом с отрицательно заряженный Асп в глубине кармана. В этой
- 9. В гидрофобном кармане, образованном радикалами Гли, Три и Лей химотрипсина располагается боковая цепь с ароматическим кольцом
- 10. В основе химических реакций лежит образование и разрыв химических связей По характеру разрыва ковалентных связей различают
- 11. Биологически важными нуклеофилами являются аминогруппы, гидроксильные группы, имидазольные группы и сульфгидрильные группы аминокислот. Нуклеофильные формы этих
- 12. Электрофильные реагенты : Наиболее известными электрофилами в биохимических реакциях являются Н+, ионы металлов, углерод карбонильной группы.
- 13. По направлению реакций с учетом конечного результата можно выделить следующие типы реакций 1. Окислительно-восстановительные. Многие окислительно-
- 14. 2. Реакции кислотно- основного взаимодействия 3.Реакции замещения 4.Реакции отщепления 5.Реакции перегруппировки 6.реакции, сопровождающиеся образованием двойной связи
- 15. Факторы, определяющие активность ферментов [E], [S], [P], Km. Влияние pH, [P], tº, ионной силы на активность
- 16. Существенное влияние на активность ферментов оказывает реакция среды. Для проявления их оптимального действия чаще всего существует
- 17. В некоторых случаях сдвиг pH на единицу снижает активность на 80%. Поэтому в экспериментальных условиях работы
- 18. Оптимум рН
- 19. Фермент pH Липаза (подж.железа) 8.0 Липаза (желудок) 4.0-5.0 Липаза(касторовое масло) 4.7 Пепсин 1.5-1.6 Трипсин 8-8.77 Уреаза
- 20. Влияние температуры
- 21. Так как все ферменты являются белками, а белки при температуре выше 40-500 С в большинстве своем
- 22. Активность фермента повышается при повышении температуры. Начиная с определенной температуры, совпадающей с началом денатурации белка, активность
- 23. Специфичность ферментов Специфичность у разных ферментов может проявляться по-разному. Ферменты как белки, построены из L-аминокислот и
- 24. Правда, лишь небольшая часть ферментов обладает абсолютной специфичностью, т.е. катализирует превращение только одного субстрата. Чаще всего
- 25. Регуляция активности ферментов Регуляция активности ферментов бывает пассивная (с помощью изменения условий среды) т. е. есть
- 26. Активная регуляция: изостерическая;( изос- равный) регуляция с помощью субстрата и продукта, Р и S аллостерическая регуляция(
- 27. Регуляция путем изменения количества фермента. У бактерий хорошо изучен феномен индуцированного синтеза ферментов при выращивании на
- 28. Замена глюкозы на лактозу приводит к индуцированному синтезу фермента галактозидазы, расщепляющей лактозу на глюкозу и галактозу.
- 29. В животных тканях подобный быстрый синтез ферментов наблюдается реже, однако при поступлении в организм некоторых ядов,
- 30. С другой стороны, иногда под действием этих гидроксилаз чужеродные вещества превращаются в более токсичные продукты (летальный
- 31. Регуляция активности по принципу обратной связи. Допустим в клетке есть многоступенчатый биосинтетический процесс, каждая стадия которого
- 32. Аллостерическая регуляция. Аллостерические ферменты - это ферменты, располагающиеся в начале метаболического потока или на его узловых
- 33. Свойства аллостерических ферментов: 1. Являются олигомерами состоящими из протомеров. 2. Имеют как минимум два центра: активный
- 34. Существует 2 вида веществ (эффекторы), которые оказывают на фермент двоякое действие: 1)активаторы; 2) ингибиторы. Аллостерический фермент
- 35. При взаимодействии аллостерического фермента с аллостерическим активатором резко возрастает степень сродства фермента к субстрату, точнее возрастает
- 36. Кинетика ферментативных реакций
- 37. Имеется реакция: S → P + Q Представим эту реакцию в виде отдельных новых стадий: S
- 38. Основы термодинамики катализа Д. Кошланд предположил, что с термодинамической точки зрения ферменты ускоряют химические реакции за
- 39. Энергия активации Энергия активации - энергия, необходимая для перевода всех молекул моля вещества в активное состояние
- 40. Термодинамика ферментативных реакций Энергетический барьер G2 G1 ΔG = G1 – G2 GA1 GA2 Энергия активации
- 41. Термодинамика ферментативных реакций Энергетический барьер G2 G1 ΔG = G2 – G1 GA1 GA2 Энергия активации
- 42. Кинетика ферментативных реакций. Концентрация фермента. Чем выше концентрация E, тем выше скорость реакции. [E] V
- 43. Кинетика ферментативных реакций. pH Для каждого фермента существует оптимальная область pH (6,9 – 7,0 для большинства
- 44. Кинетика ферментативных реакций. Температура С увеличением температуры на 10°C скорость реакции возрастает в 2 раза (правило
- 45. Активность фермента зависит от концентрации субстратов. Исследование зависимости скорости ферментативных реакций от концентрации реагирующих веществ стало
- 46. Кинетика ферментативных реакций
- 47. Если концентрация субстрата [S] очень низкая, ограничивающим скорость реакции становится этап образования комплекса ES (связывание) и
- 48. Если концентрация субстрата [S] высокая, количество образующегося комплекса ES зависит от количества доступного фермента, и скорость
- 49. Ограничивающим скорость реакции становится этап образования продукта (катализ). В реакции, катализируемой ферментом имеется верхний предел скорости
- 50. Кинетика ферментативных реакций. Концентрация субстрата Для простых ферментов график имеет вид гиперболы и описывается уравнением Михаэлиса-Ментен.
- 51. [ES]max = [Et]общ Общее количество фермента в системе, равно сумме [E] (концентрация свободного фермента), и [ES]
- 52. Важным для правильной оценки результатов исследования зависимости скорости реакции от концентрации является измерение начальной скорости реакции.
- 53. При взаимодействии фермента и субстрата очень быстро наступает равновесие между скоростью образования и скоростью распада фермент
- 54. В реакции, катализируемой ферментом можно выделить четыре реакции, каждая из которых характеризуется собственной константой скорости. Однако,
- 55. Важной качественной характеристикой фермента является константа Михаэлиса Воспользовавшись предположениями, высказанными Генри, Михаэлисом и Ментен, а также
- 56. Основная гипотеза: этапом, ограничивающим скорость ферментативной реакции является ( ES → E + P ) отсюда
- 57. тогда, количество свободного фермента: [Et] - [ES] так как [S] >> [Et] , [S]связ [этап 1]
- 58. [этап 2] гипотеза: Образование фермент-субстратного комплекса самая быстрая реакция, результатом которой является возникновение динамического равновесия между
- 59. [Этап 3] k1 [Et] [S] - k1 [ES] [S] = [ES] (k2 + k3) k1 [Et]
- 60. отсюда v0 = k3 [ES] = k3 {[Et] [S] / { [S] + Km } =
- 61. Уравнение Михаэлиса и Ментен графически – прямоугольная гипербола Если мы простроим график зависимости скорости реакции V
- 63. Каково физическое значение Km? Уравнение Михаэлиса-Ментен можно преобразовать к такому виду
- 64. Из этого уравнения легко показать, что при [S] =10 Kmv/Vmax = 0.91при [S] = Kmv/ Vmax
- 65. При условии, что k3
- 66. Значение Km дает также некоторые представления относительно эффективности катализа и регуляции. Если [S]>> 10 Km, реакция
- 67. Если [S]
- 68. Наиболее удобное сочетание эффективности и контроля соблюдается при условии, если концентрация субстрата одного порядка со значениями
- 69. фермента или в клинической лаборатории или исследовательской лаборатории, следует насыщать фермент субстратом. Знание Км позволит Вам
- 70. В физиологических условиях, для эффективной работы концентрация субстрата должна быть на уровне Км этого фермента, но
- 71. Практически рассчитать значения Км и Vmax, пользуясь кривой, описываемой уравнением Михаэлис и Ментен сложно. Более удобно
- 72. Такой способ выражения позволяет более точно рассчитать значения Км и V. Пересечение линии с осью 1/[S]
- 74. Примеры использования данных кинетических исследований ферментов в медицине Некоторые люди обладают повышенной чувствительностью к этиловому спирту.
- 75. Альдегиддегидрогеназа обычно существует в двух формах, с высоким сродством (низкие значения Км) к альдегиду и с
- 76. Семейная пара с генетической предрасположенностью к болезни Неймана-Пика ожидает ребенка. Им известно, что их будущий малыш
- 77. Если сфингомиэлин не распадается должным образом, нарушается нейронная передача. Физиологические последствия болезни Наймана-Пика - олигофрения и
- 78. Семейная пара с генетической предрасположенностью к болезни Неймана-Пика ожидает ребенка. Им известно, что их будущий малыш
- 79. У беременной были получены клетки плода (путем амниоцентеза) и размножены методом тканевой культуры. Экстракт клеток был
- 81. Значения Vmax и Km определяются при экстраполяции линий до пересечения с осью абсцисс и ординат. Как
- 83. Скачать презентацию

Выдвинутая в 1913 году Л. Михаэлисом и
М. Ментен общая
Выдвинутая в 1913 году Л. Михаэлисом и
М. Ментен общая
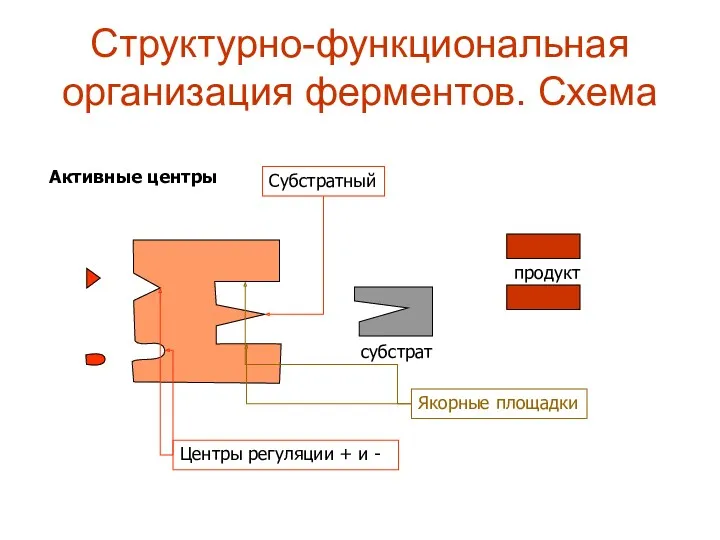
Структурно-функциональная организация ферментов. Схема
Активные центры
Якорные площадки
Субстратный
субстрат
продукт
Центры регуляции + и -
Структурно-функциональная организация ферментов. Схема
Активные центры
Якорные площадки
Субстратный
субстрат
продукт
Центры регуляции + и -

Структурно-функциональная организация ферментов.
Активный (субстратный) центр - это совокупность функциональных групп,
Структурно-функциональная организация ферментов.
Активный (субстратный) центр - это совокупность функциональных групп,

Активный центр включает в себя:
Каталитический участок или центр, непосредственно взаимодействующий с
Активный центр включает в себя:
Каталитический участок или центр, непосредственно взаимодействующий с

1 этап: постепенное «причаливание» S к «якорной» площадке F.
2 этап: напряжение
1 этап: постепенное «причаливание» S к «якорной» площадке F.
2 этап: напряжение

Химические связи, действующие при этом:
1. Силы Ван дер Ваальса
2. Электростатическое взаимодействие
3.
Химические связи, действующие при этом:
1. Силы Ван дер Ваальса
2. Электростатическое взаимодействие
3.

Якорный участок трипсина представлен длинным узким карманом с отрицательно заряженный Асп
Якорный участок трипсина представлен длинным узким карманом с отрицательно заряженный Асп

В гидрофобном кармане, образованном радикалами Гли, Три и Лей химотрипсина располагается
В гидрофобном кармане, образованном радикалами Гли, Три и Лей химотрипсина располагается

В основе химических реакций лежит образование и разрыв химических связей
По
В основе химических реакций лежит образование и разрыв химических связей
По

Биологически важными нуклеофилами являются аминогруппы, гидроксильные группы, имидазольные группы и
Биологически важными нуклеофилами являются аминогруппы, гидроксильные группы, имидазольные группы и

Электрофильные реагенты : Наиболее известными электрофилами в биохимических реакциях являются Н+,
Электрофильные реагенты : Наиболее известными электрофилами в биохимических реакциях являются Н+,

По направлению реакций с учетом конечного результата можно выделить следующие типы
По направлению реакций с учетом конечного результата можно выделить следующие типы

2. Реакции кислотно- основного взаимодействия
3.Реакции замещения
4.Реакции отщепления
5.Реакции перегруппировки
6.реакции, сопровождающиеся образованием двойной
3.Реакции замещения
4.Реакции отщепления
5.Реакции перегруппировки
6.реакции, сопровождающиеся образованием двойной
![Факторы, определяющие активность ферментов [E], [S], [P], Km. Влияние pH,](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/270654/slide-14.jpg)
Факторы, определяющие активность ферментов [E], [S], [P], Km.
Влияние pH, [P],
Влияние pH, [P],

Существенное влияние на активность ферментов оказывает реакция среды. Для проявления
Существенное влияние на активность ферментов оказывает реакция среды. Для проявления

В некоторых случаях сдвиг pH на единицу снижает активность на 80%.
В некоторых случаях сдвиг pH на единицу снижает активность на 80%.
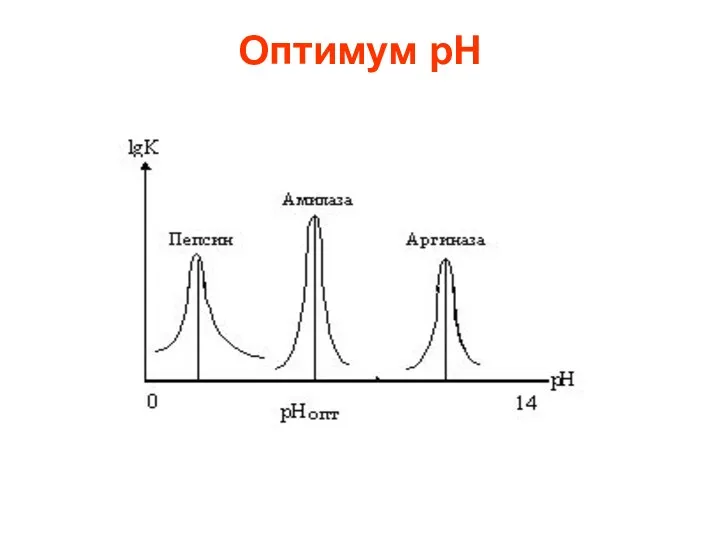
Оптимум рН
Оптимум рН

Фермент pH
Липаза (подж.железа) 8.0
Липаза (желудок) 4.0-5.0
Липаза(касторовое масло) 4.7
Пепсин 1.5-1.6
Трипсин 8-8.77
Уреаза
Фермент pH
Липаза (подж.железа) 8.0
Липаза (желудок) 4.0-5.0
Липаза(касторовое масло) 4.7
Пепсин 1.5-1.6
Трипсин 8-8.77
Уреаза
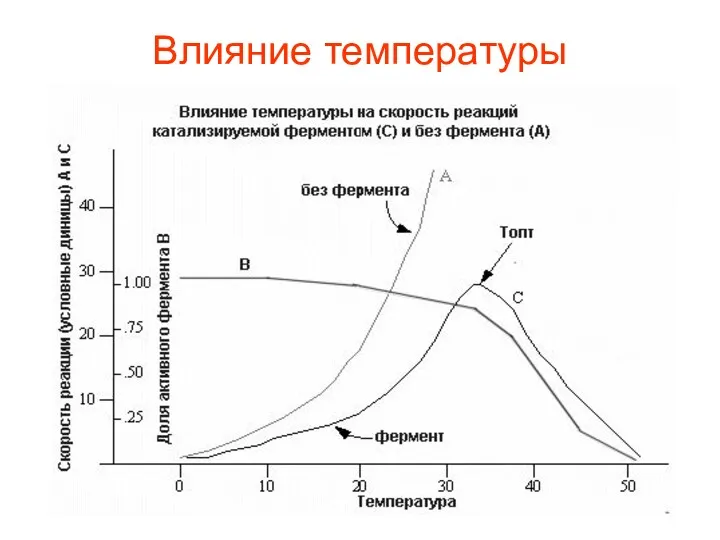
Влияние температуры
Влияние температуры

Так как все ферменты являются белками, а белки при температуре выше
Так как все ферменты являются белками, а белки при температуре выше

Активность фермента повышается при повышении температуры. Начиная с определенной температуры, совпадающей
Активность фермента повышается при повышении температуры. Начиная с определенной температуры, совпадающей

Специфичность ферментов
Специфичность у разных ферментов может проявляться по-разному. Ферменты как
Специфичность ферментов
Специфичность у разных ферментов может проявляться по-разному. Ферменты как

Правда, лишь небольшая часть ферментов обладает абсолютной специфичностью, т.е. катализирует превращение
Правда, лишь небольшая часть ферментов обладает абсолютной специфичностью, т.е. катализирует превращение

Регуляция активности ферментов
Регуляция активности ферментов бывает пассивная (с помощью изменения условий
Регуляция активности ферментов
Регуляция активности ферментов бывает пассивная (с помощью изменения условий

Активная регуляция:
изостерическая;( изос- равный) регуляция с помощью субстрата и продукта, Р
Активная регуляция:
изостерическая;( изос- равный) регуляция с помощью субстрата и продукта, Р

Регуляция путем изменения количества фермента.
У бактерий хорошо изучен феномен индуцированного синтеза
Регуляция путем изменения количества фермента.
У бактерий хорошо изучен феномен индуцированного синтеза

Замена глюкозы на лактозу приводит к индуцированному синтезу фермента галактозидазы, расщепляющей
Замена глюкозы на лактозу приводит к индуцированному синтезу фермента галактозидазы, расщепляющей

В животных тканях подобный быстрый синтез ферментов наблюдается реже, однако при
В животных тканях подобный быстрый синтез ферментов наблюдается реже, однако при

С другой стороны, иногда под действием этих гидроксилаз чужеродные вещества превращаются
С другой стороны, иногда под действием этих гидроксилаз чужеродные вещества превращаются

Регуляция активности по принципу обратной связи.
Допустим в клетке есть многоступенчатый
Регуляция активности по принципу обратной связи.
Допустим в клетке есть многоступенчатый

Аллостерическая регуляция.
Аллостерические ферменты - это ферменты, располагающиеся в начале
Аллостерическая регуляция.
Аллостерические ферменты - это ферменты, располагающиеся в начале

Свойства аллостерических ферментов:
1. Являются олигомерами состоящими из протомеров.
2. Имеют как минимум два
Свойства аллостерических ферментов:
1. Являются олигомерами состоящими из протомеров.
2. Имеют как минимум два

Существует 2 вида веществ (эффекторы), которые оказывают на фермент двоякое действие:
1)активаторы;
Существует 2 вида веществ (эффекторы), которые оказывают на фермент двоякое действие:
1)активаторы;

При взаимодействии аллостерического фермента с аллостерическим активатором резко возрастает степень сродства
При взаимодействии аллостерического фермента с аллостерическим активатором резко возрастает степень сродства

Кинетика ферментативных реакций
Кинетика ферментативных реакций

Имеется реакция:
S → P + Q
Представим эту реакцию в виде
Имеется реакция:
S → P + Q
Представим эту реакцию в виде

Основы термодинамики катализа
Д. Кошланд предположил, что с термодинамической точки зрения ферменты
Основы термодинамики катализа
Д. Кошланд предположил, что с термодинамической точки зрения ферменты

Энергия активации
Энергия активации - энергия, необходимая для перевода всех молекул моля
Энергия активации
Энергия активации - энергия, необходимая для перевода всех молекул моля
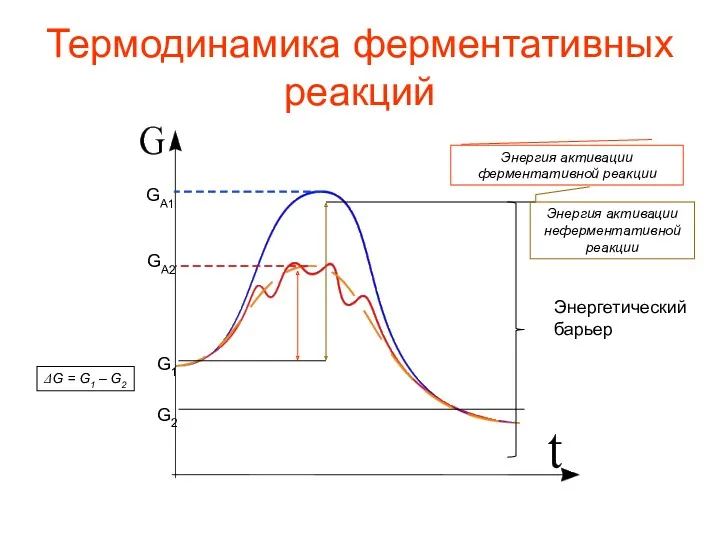
Термодинамика ферментативных реакций
Энергетический
барьер
G2
G1
ΔG = G1 – G2
GA1
GA2
Энергия активации неферментативной реакции
Энергия активации
Термодинамика ферментативных реакций
Энергетический
барьер
G2
G1
ΔG = G1 – G2
GA1
GA2
Энергия активации неферментативной реакции
Энергия активации
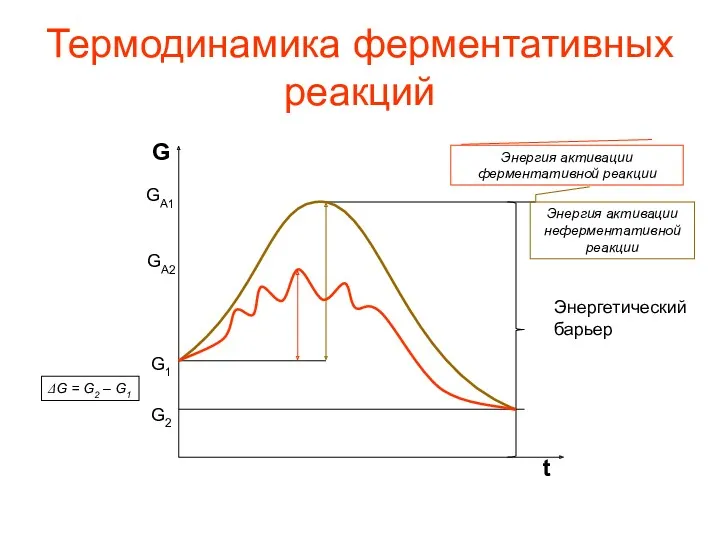
Термодинамика ферментативных реакций
Энергетический
барьер
G2
G1
ΔG = G2 – G1
GA1
GA2
Энергия активации неферментативной реакции
Энергия активации
Термодинамика ферментативных реакций
Энергетический
барьер
G2
G1
ΔG = G2 – G1
GA1
GA2
Энергия активации неферментативной реакции
Энергия активации
![Кинетика ферментативных реакций. Концентрация фермента. Чем выше концентрация E, тем выше скорость реакции. [E] V](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/270654/slide-41.jpg)
Кинетика ферментативных реакций. Концентрация фермента.
Чем выше концентрация E, тем выше скорость
Кинетика ферментативных реакций. Концентрация фермента.
Чем выше концентрация E, тем выше скорость
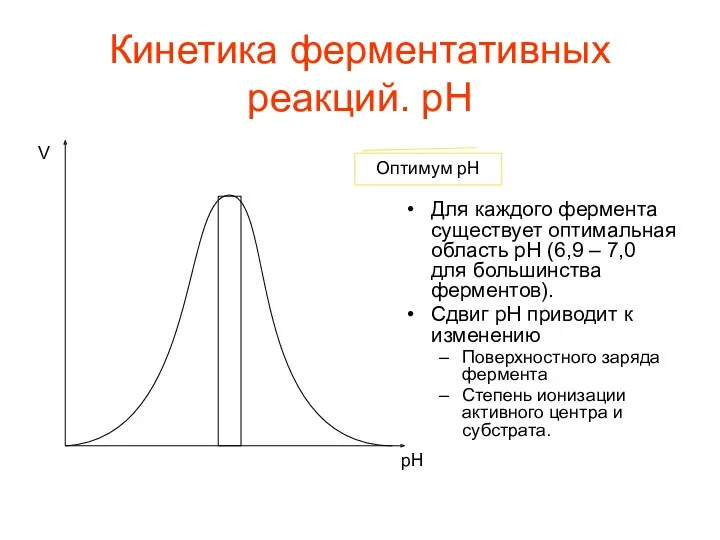
Кинетика ферментативных реакций. pH
Для каждого фермента существует оптимальная область pH (6,9
Кинетика ферментативных реакций. pH
Для каждого фермента существует оптимальная область pH (6,9
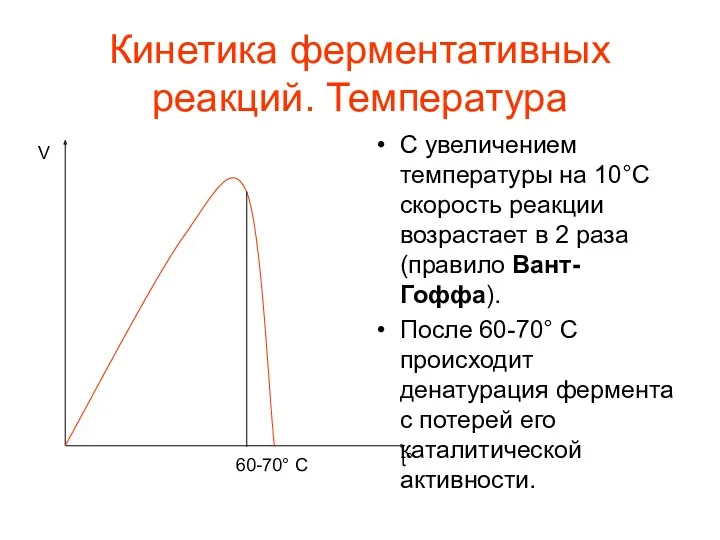
Кинетика ферментативных реакций. Температура
С увеличением температуры на 10°C скорость реакции возрастает
Кинетика ферментативных реакций. Температура
С увеличением температуры на 10°C скорость реакции возрастает

Активность фермента зависит от концентрации субстратов.
Исследование зависимости скорости ферментативных реакций
Активность фермента зависит от концентрации субстратов.
Исследование зависимости скорости ферментативных реакций
Кинетика ферментативных реакций
Кинетика ферментативных реакций
![Если концентрация субстрата [S] очень низкая, ограничивающим скорость реакции становится](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/270654/slide-46.jpg)
Если концентрация субстрата [S] очень низкая, ограничивающим скорость реакции становится этап
Если концентрация субстрата [S] очень низкая, ограничивающим скорость реакции становится этап
![Если концентрация субстрата [S] высокая, количество образующегося комплекса ES зависит](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/270654/slide-47.jpg)
Если концентрация субстрата [S] высокая, количество образующегося комплекса ES зависит
Если концентрация субстрата [S] высокая, количество образующегося комплекса ES зависит

Ограничивающим скорость реакции становится этап образования продукта (катализ). В реакции, катализируемой
Ограничивающим скорость реакции становится этап образования продукта (катализ). В реакции, катализируемой
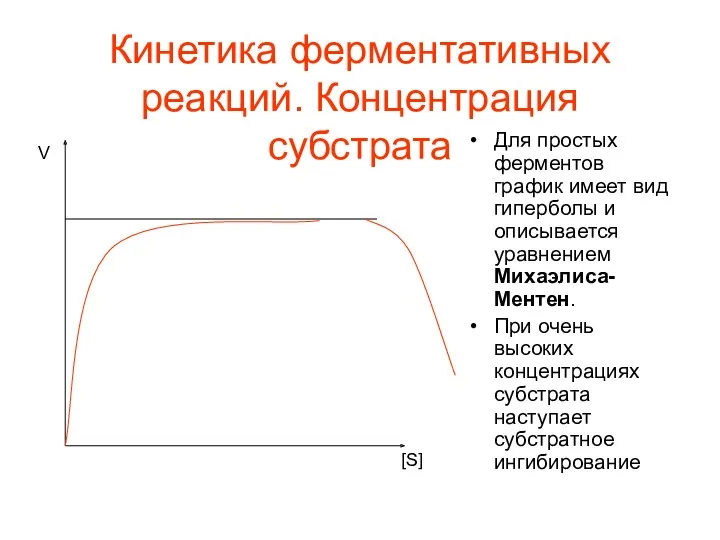
Кинетика ферментативных реакций. Концентрация субстрата
Для простых ферментов график имеет вид гиперболы
Кинетика ферментативных реакций. Концентрация субстрата
Для простых ферментов график имеет вид гиперболы
![[ES]max = [Et]общ Общее количество фермента в системе, равно сумме](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/270654/slide-50.jpg)
[ES]max = [Et]общ
Общее количество фермента в системе, равно сумме [E]
[ES]max = [Et]общ
Общее количество фермента в системе, равно сумме [E]

Важным для правильной оценки результатов исследования зависимости скорости реакции от
Важным для правильной оценки результатов исследования зависимости скорости реакции от

При взаимодействии фермента и субстрата очень быстро наступает равновесие между скоростью
При взаимодействии фермента и субстрата очень быстро наступает равновесие между скоростью

В реакции, катализируемой ферментом можно выделить четыре реакции, каждая из которых
В реакции, катализируемой ферментом можно выделить четыре реакции, каждая из которых

Важной качественной характеристикой фермента является константа Михаэлиса
Воспользовавшись предположениями, высказанными Генри,
Важной качественной характеристикой фермента является константа Михаэлиса
Воспользовавшись предположениями, высказанными Генри,

Основная гипотеза: этапом, ограничивающим скорость ферментативной реакции является ( ES →
Основная гипотеза: этапом, ограничивающим скорость ферментативной реакции является ( ES →
![тогда, количество свободного фермента: [Et] - [ES] так как [S]](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/270654/slide-56.jpg)
тогда, количество свободного фермента: [Et] - [ES]
так как [S] >>
тогда, количество свободного фермента: [Et] - [ES]
так как [S] >>
![[этап 2] гипотеза: Образование фермент-субстратного комплекса самая быстрая реакция, результатом](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/270654/slide-57.jpg)
[этап 2] гипотеза: Образование фермент-субстратного комплекса самая быстрая реакция, результатом которой является
[этап 2] гипотеза: Образование фермент-субстратного комплекса самая быстрая реакция, результатом которой является
![[Этап 3] k1 [Et] [S] - k1 [ES] [S] =](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/270654/slide-58.jpg)
[Этап 3] k1 [Et] [S] - k1 [ES] [S] = [ES]
[Этап 3] k1 [Et] [S] - k1 [ES] [S] = [ES]
![отсюда v0 = k3 [ES] = k3 {[Et] [S] /](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/270654/slide-59.jpg)
отсюда v0 = k3 [ES] = k3 {[Et] [S] / {
отсюда v0 = k3 [ES] = k3 {[Et] [S] / {
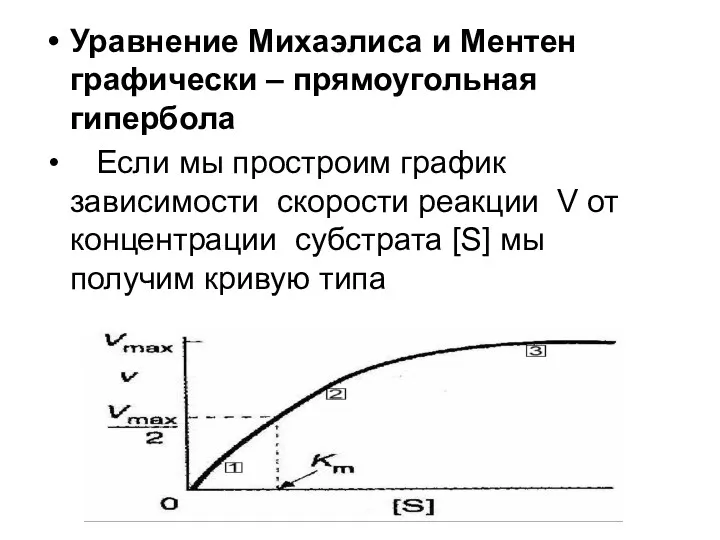
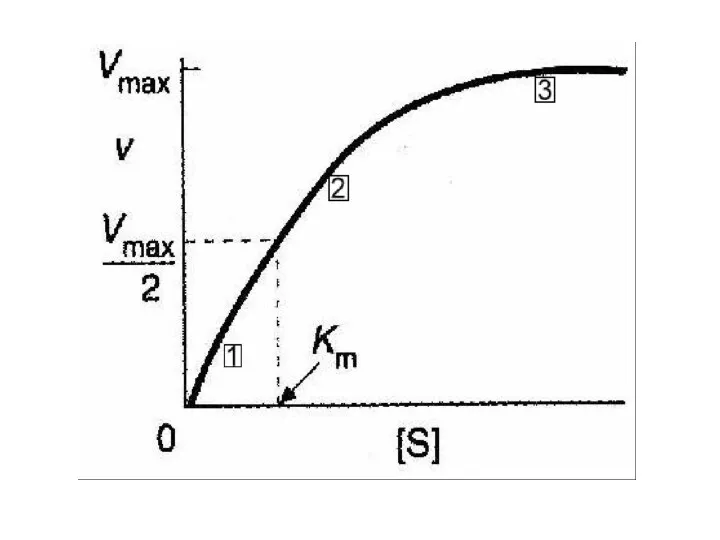
Уравнение Михаэлиса и Ментен графически – прямоугольная гипербола
Если мы простроим
Уравнение Михаэлиса и Ментен графически – прямоугольная гипербола
Если мы простроим

Каково физическое значение Km? Уравнение Михаэлиса-Ментен можно преобразовать к такому виду
Каково физическое значение Km? Уравнение Михаэлиса-Ментен можно преобразовать к такому виду
![Из этого уравнения легко показать, что при [S] =10 Kmv/Vmax](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/270654/slide-63.jpg)
Из этого уравнения легко показать, что
при [S] =10 Kmv/Vmax =
Из этого уравнения легко показать, что
при [S] =10 Kmv/Vmax =

При условии, что k3 << k2, константа Михаэлиса становится хорошим показателем
При условии, что k3 << k2, константа Михаэлиса становится хорошим показателем

Значение Km дает также некоторые представления относительно эффективности катализа и регуляции.
Значение Km дает также некоторые представления относительно эффективности катализа и регуляции.
![Если [S]](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/270654/slide-66.jpg)
Если [S] << 0.1 Km, эффективность реакции низка, но имеется хорошее
Если [S] << 0.1 Km, эффективность реакции низка, но имеется хорошее

Наиболее удобное сочетание эффективности и контроля соблюдается при условии, если концентрация
Наиболее удобное сочетание эффективности и контроля соблюдается при условии, если концентрация

фермента или в клинической лаборатории или исследовательской лаборатории, следует насыщать фермент
фермента или в клинической лаборатории или исследовательской лаборатории, следует насыщать фермент

В физиологических условиях, для эффективной работы концентрация субстрата должна быть на
В физиологических условиях, для эффективной работы концентрация субстрата должна быть на

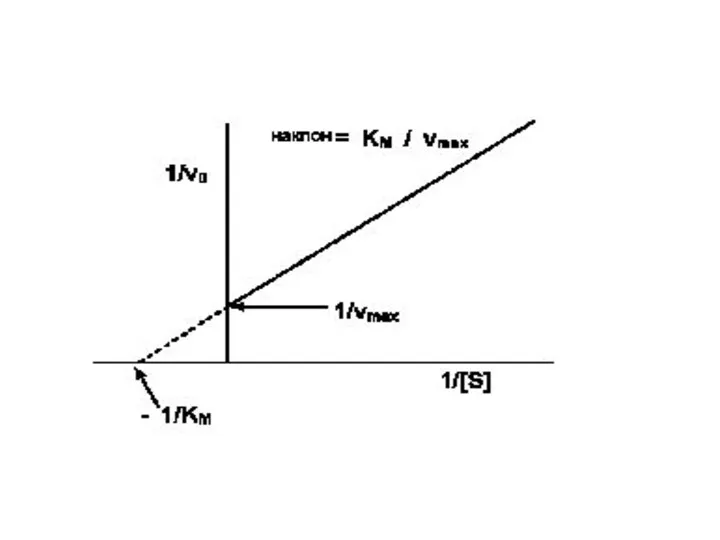
Практически рассчитать значения Км и Vmax, пользуясь кривой, описываемой уравнением
Практически рассчитать значения Км и Vmax, пользуясь кривой, описываемой уравнением

Такой способ выражения позволяет более точно рассчитать значения Км и
Такой способ выражения позволяет более точно рассчитать значения Км и

Примеры использования данных кинетических исследований ферментов в медицине
Некоторые люди обладают
Примеры использования данных кинетических исследований ферментов в медицине
Некоторые люди обладают

Альдегиддегидрогеназа обычно существует в двух формах, с высоким сродством (низкие значения
Альдегиддегидрогеназа обычно существует в двух формах, с высоким сродством (низкие значения

Семейная пара с генетической предрасположенностью к болезни Неймана-Пика ожидает ребенка. Им
Семейная пара с генетической предрасположенностью к болезни Неймана-Пика ожидает ребенка. Им

Если сфингомиэлин не распадается должным образом, нарушается нейронная передача. Физиологические последствия
Если сфингомиэлин не распадается должным образом, нарушается нейронная передача. Физиологические последствия

Семейная пара с генетической предрасположенностью к болезни Неймана-Пика ожидает ребенка. Им
Семейная пара с генетической предрасположенностью к болезни Неймана-Пика ожидает ребенка. Им

У беременной были получены клетки плода (путем амниоцентеза) и размножены методом
У беременной были получены клетки плода (путем амниоцентеза) и размножены методом
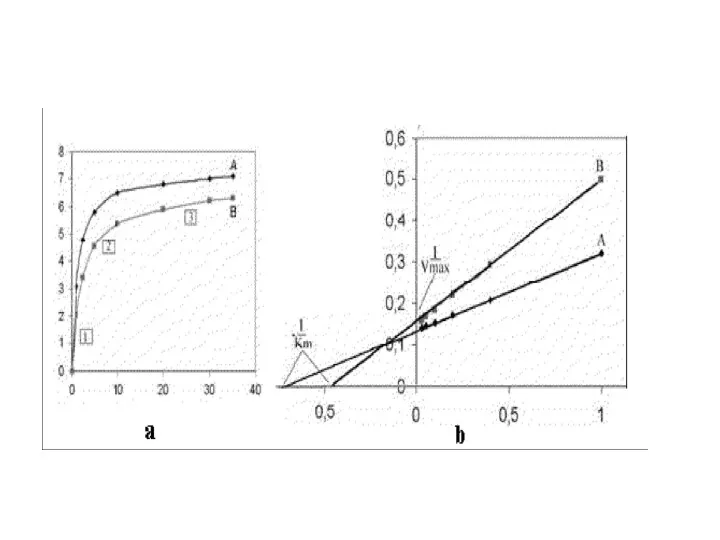

Значения Vmax и Km определяются при экстраполяции линий до пересечения с
Значения Vmax и Km определяются при экстраполяции линий до пересечения с
 Основные генетические и морфофизиологические характеристики популяции
Основные генетические и морфофизиологические характеристики популяции Строение и свойства белков.
Строение и свойства белков. Тыныс алудың организм үшін маңызы. Тыныс алу кезеңдері. Тыныс алудың реттелуі
Тыныс алудың организм үшін маңызы. Тыныс алу кезеңдері. Тыныс алудың реттелуі блок_8_Зоо_Млекопитающие
блок_8_Зоо_Млекопитающие Опытно-экспериментальная работа на учебно-опытном участке
Опытно-экспериментальная работа на учебно-опытном участке Цитоплазма. Структура еукаріотичної клітини
Цитоплазма. Структура еукаріотичної клітини Введение в иммунологию. Иммунная система
Введение в иммунологию. Иммунная система Высшие споровые растения. 7 класс
Высшие споровые растения. 7 класс Сучасні критерії виду
Сучасні критерії виду Сера и её роль в организме человека
Сера и её роль в организме человека Елементний склад живих організмів. Неорганічні речовини: вода і мінеральні солі
Елементний склад живих організмів. Неорганічні речовини: вода і мінеральні солі Презентация по теме Химический состав клетки
Презентация по теме Химический состав клетки Влияние света на проращивание семян (часть 1)
Влияние света на проращивание семян (часть 1) Проект на тему Мой домашний зоопарк
Проект на тему Мой домашний зоопарк Кільчасті черви
Кільчасті черви Движение крови и лимфы
Движение крови и лимфы Решение задач части С5. Молекулярная биология
Решение задач части С5. Молекулярная биология Бактерии. Грибы. Общая характеристика грибов
Бактерии. Грибы. Общая характеристика грибов История развития биологии
История развития биологии Возрастные особенности температурной сенсорной системы. Механизмы закаливания
Возрастные особенности температурной сенсорной системы. Механизмы закаливания Мужская половая система
Мужская половая система Скелет человека
Скелет человека Ткани растений
Ткани растений Международная классификация микроорганизмов по Берги
Международная классификация микроорганизмов по Берги Динозавры — удивительные существа
Динозавры — удивительные существа Улитки Ахатины - исследовательская работа
Улитки Ахатины - исследовательская работа Сердечно-сосудистая система. Физиологические свойства и функции сердца
Сердечно-сосудистая система. Физиологические свойства и функции сердца Мышечная система
Мышечная система