- Механизмы оплодотворения и морфогенеза

Содержание
- 2. Видоспецифичность оплодотворения Дистантные взаимодействия характерны для водных организмов, с наружным типом оплодотворения. При этом животные сталкиваются
- 3. Видоспецифичность оплодотворения Модель узнавания спермием и яйцом друг друга у млекопитающих (модель капацитации) (по Гилберту, 1993):
- 4. Видоспецифичность оплодотворения Молекулы байндина, покрывающие поверхность акросомального отростка сперматозоида морского ежа (сильно схематизировано). Как полагают, эти
- 5. Блокирование полиспермии Существуют механизмы защиты яйцеклетки от полиспермии: Быстрый блок полиспермии Медленный блок полиспермии Рис. 5.
- 6. Блокирование полиспермии
- 7. Блокирование полиспермии Модель распознавания гамет на поверхности zona pellucida. У мышей zona pellucida (аквамарин) состоит из
- 8. Роль ионных сдвигов в активации биосинтетических процессов в яйце Схема возможных соотношений между разными событиями в
- 9. Схематическое изображение сперматозоидов яйцо сигнализации, что приводит к последовательному Ca2 + освобождение от ER через IP3R
- 10. Генетическая неровность пронуклеусов у млекопитающих Зародыши мыши на 11-й день беременности: А - контрольный. Б -
- 11. Генетическая неровность пронуклеусов у млекопитающих male pronucleus female pronucleus
- 12. Генетическая неровность пронуклеусов у млекопитающих 5hmC и 5mC в начале пронуклеусов стадии зиготы. (А) зиготы пронуклеусов
- 13. Генетическая неровность пронуклеусов у млекопитающих Общие изменения в уровнях 5mC и 5hmC в пронуклеарной стадии зигот
- 14. Явление геномного импринтинга Геномный импринтинг - это эпигенетический процесс, который происходит в течение овогенеза и сперматогенеза
- 15. Экспрессия импринтированных генов Импринтированные гены экспрессируются с одной родительской аллели и часто собраны в кластеры. Большинство
- 16. Таблица. Функция импринтированных генов, определяемая путем инактивации генов. Особенности кластеров импринтированных генов в геноме мыши. (М)
- 17. Молекулярно-генетические механизмы морфогенеза Перемещение цитоплазматических масс в яйце асцидии Styela partita. А. До оплодотворения желтая кортикальная
- 18. Молекулярно-генетические механизмы морфогенеза
- 19. сперакт и резакт (Морские ежи) Гиногамоны Андрогомоны Глюкозил-трансфераза N-ацетил- глюкозамин
- 21. Izumo Juno
- 22. 5hmC 5mC Tet3 5mC 5mC
- 23. P1+ P2P3P4 - P + Р+ Р+
- 24. HNF-4 C-mos Notch I
- 26. Скачать презентацию

Видоспецифичность оплодотворения
Дистантные взаимодействия характерны для водных организмов, с наружным типом оплодотворения.
Видоспецифичность оплодотворения
Дистантные взаимодействия характерны для водных организмов, с наружным типом оплодотворения.
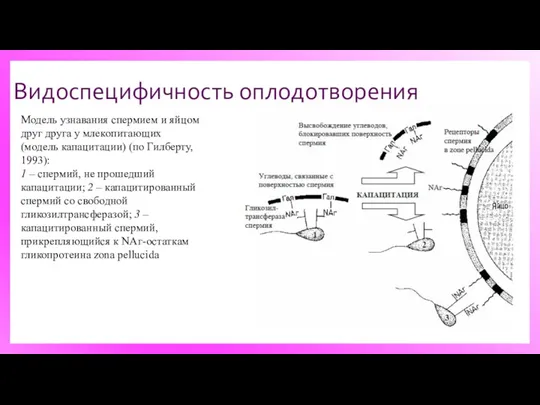
Видоспецифичность оплодотворения
Модель узнавания спермием и яйцом друг друга у млекопитающих (модель
Видоспецифичность оплодотворения
Модель узнавания спермием и яйцом друг друга у млекопитающих (модель
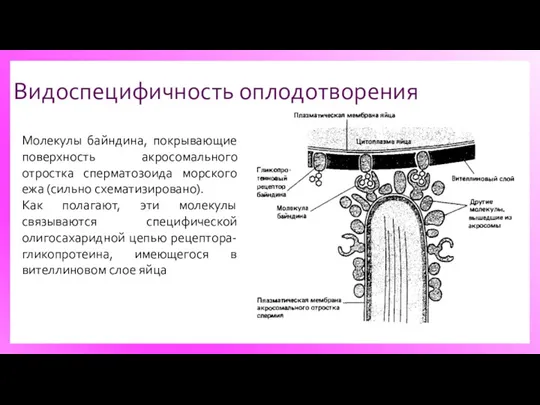
Видоспецифичность оплодотворения
Молекулы байндина, покрывающие поверхность акросомального отростка сперматозоида морского ежа (сильно
Видоспецифичность оплодотворения
Молекулы байндина, покрывающие поверхность акросомального отростка сперматозоида морского ежа (сильно
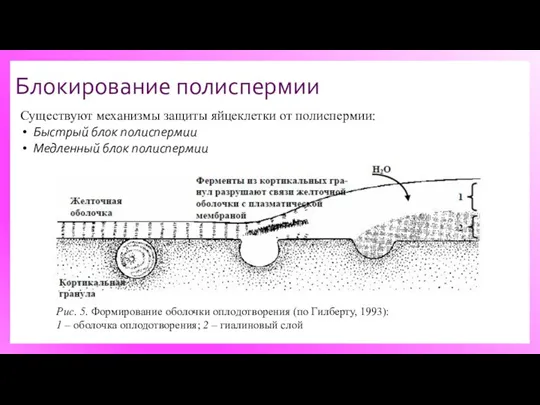
Блокирование полиспермии
Существуют механизмы защиты яйцеклетки от полиспермии:
Быстрый блок полиспермии
Медленный блок
Блокирование полиспермии
Существуют механизмы защиты яйцеклетки от полиспермии:
Быстрый блок полиспермии
Медленный блок

Блокирование
полиспермии
Блокирование
полиспермии
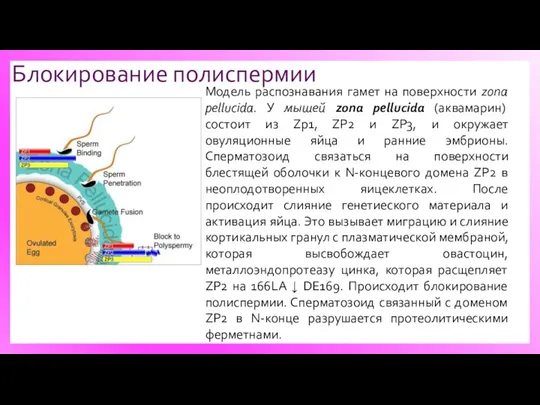
Блокирование полиспермии
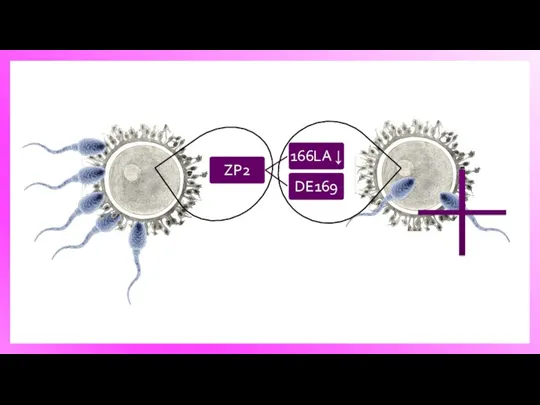
Модель распознавания гамет на поверхности zona pellucida. У мышей zona
Блокирование полиспермии
Модель распознавания гамет на поверхности zona pellucida. У мышей zona
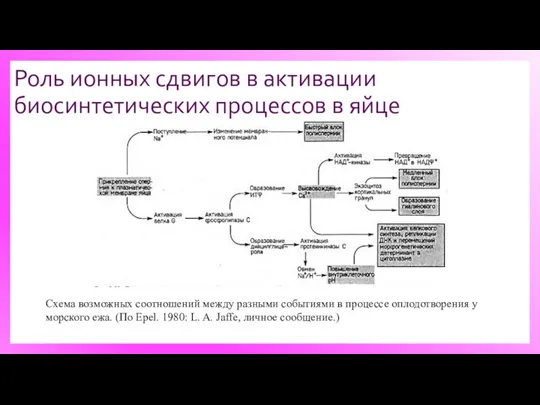
Роль ионных сдвигов в активации биосинтетических процессов в яйце
Схема возможных
Роль ионных сдвигов в активации биосинтетических процессов в яйце
Схема возможных
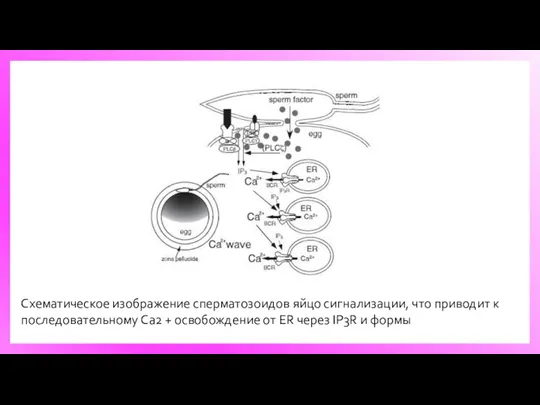
Схематическое изображение сперматозоидов яйцо сигнализации, что приводит к последовательному Ca2 +
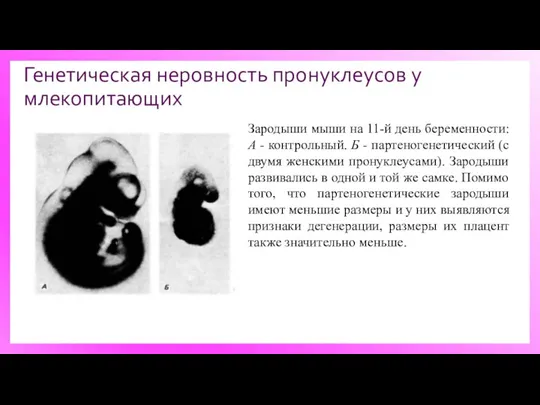
Генетическая неровность пронуклеусов у млекопитающих
Зародыши мыши на 11-й день беременности: А
Генетическая неровность пронуклеусов у млекопитающих
Зародыши мыши на 11-й день беременности: А
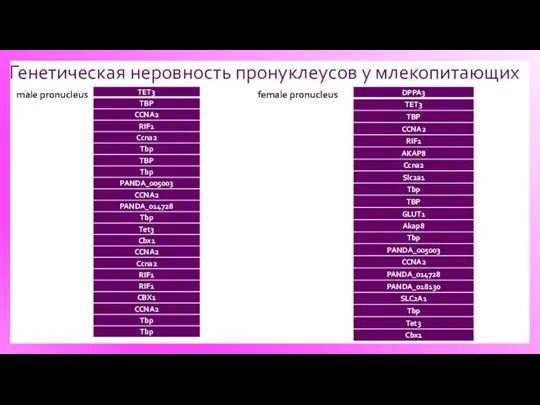
Генетическая неровность пронуклеусов у млекопитающих
male pronucleus
female pronucleus
Генетическая неровность пронуклеусов у млекопитающих
male pronucleus
female pronucleus

Генетическая неровность пронуклеусов у млекопитающих
5hmC и 5mC в начале пронуклеусов стадии
Генетическая неровность пронуклеусов у млекопитающих
5hmC и 5mC в начале пронуклеусов стадии
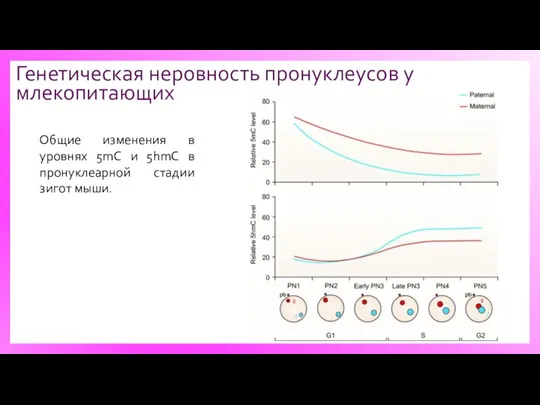
Генетическая неровность пронуклеусов у млекопитающих
Общие изменения в уровнях 5mC и 5hmC
Генетическая неровность пронуклеусов у млекопитающих
Общие изменения в уровнях 5mC и 5hmC

Явление геномного импринтинга
Геномный импринтинг - это эпигенетический процесс, который происходит
Явление геномного импринтинга
Геномный импринтинг - это эпигенетический процесс, который происходит
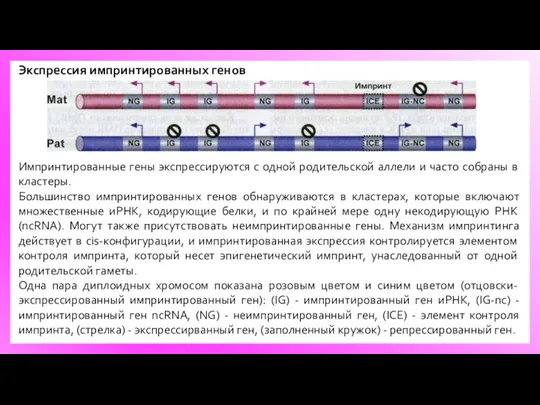
Экспрессия импринтированных генов
Импринтированные гены экспрессируются с одной родительской аллели и часто
Экспрессия импринтированных генов
Импринтированные гены экспрессируются с одной родительской аллели и часто
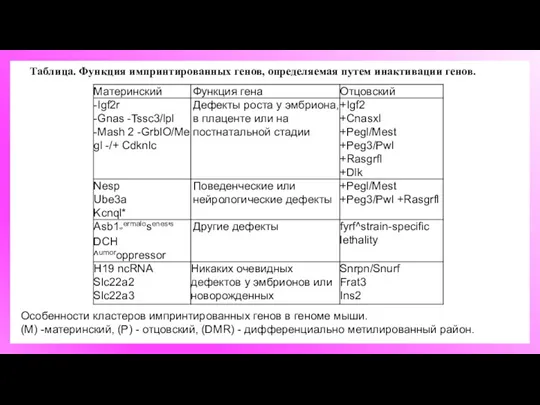
Таблица. Функция импринтированных генов, определяемая путем инактивации генов.
Особенности кластеров импринтированных
Таблица. Функция импринтированных генов, определяемая путем инактивации генов.
Особенности кластеров импринтированных
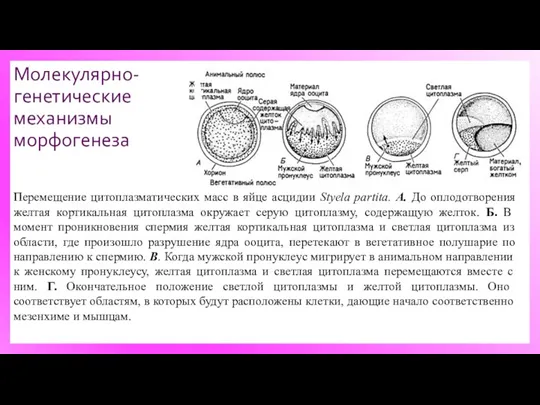
Молекулярно-генетические механизмы морфогенеза
Перемещение цитоплазматических масс в яйце асцидии Styela partita. А.
Молекулярно-генетические механизмы морфогенеза
Перемещение цитоплазматических масс в яйце асцидии Styela partita. А.

Молекулярно-генетические
механизмы морфогенеза
Молекулярно-генетические
механизмы морфогенеза
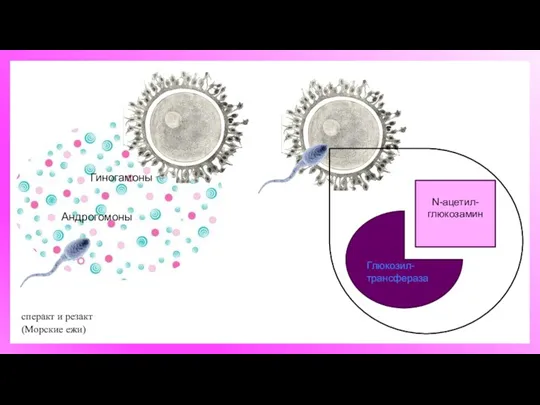
сперакт и резакт
(Морские ежи)
Гиногамоны
Андрогомоны
Глюкозил-трансфераза
N-ацетил-
глюкозамин
сперакт и резакт
(Морские ежи)
Гиногамоны
Андрогомоны
Глюкозил-трансфераза
N-ацетил-
глюкозамин
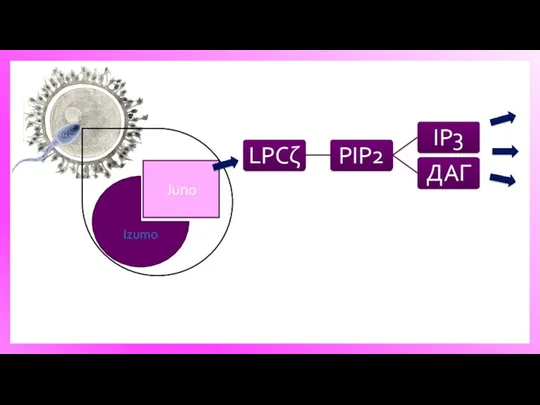
Izumo
Juno
Juno
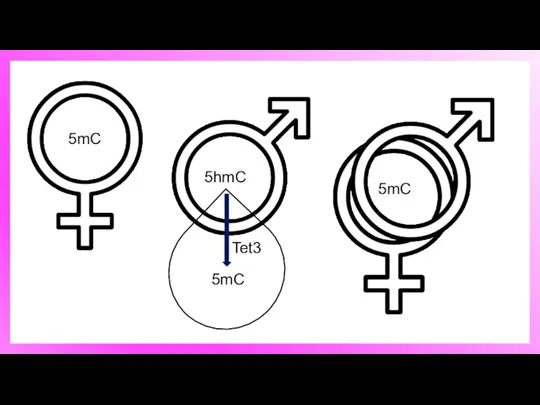
5hmC
5mC
Tet3
5mC
5mC
5hmC
5mC
Tet3
5mC
5mC
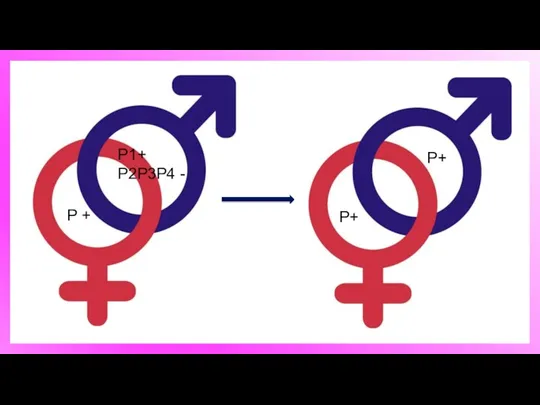
P1+
P2P3P4 -
P +
Р+
Р+
P1+
P2P3P4 -
P +
Р+
Р+
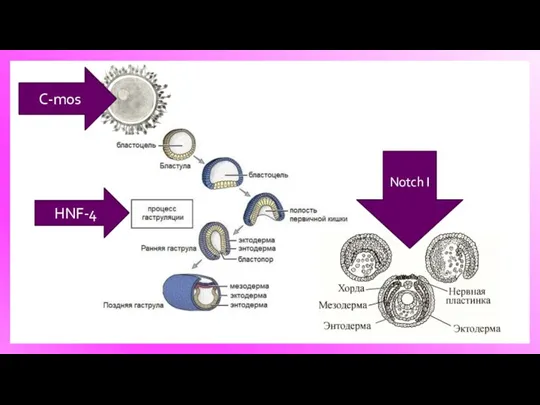
HNF-4
C-mos
Notch I
HNF-4
C-mos
Notch I
 Биология. Разбор структуры и заданий КИМ ОГЭ-2023
Биология. Разбор структуры и заданий КИМ ОГЭ-2023 Пищеварительная система и процесс пищеварения у человека
Пищеварительная система и процесс пищеварения у человека Лимфатическая система
Лимфатическая система Игра Найти лишнее. 6 класс
Игра Найти лишнее. 6 класс Клиническая анатомия висцеральных систем
Клиническая анатомия висцеральных систем Вода и ее свойства
Вода и ее свойства Внутривидовая и отдалённая гибридизация растений
Внутривидовая и отдалённая гибридизация растений Анатомия и физиология ЦНС. Свойства и принципы функционирования нервных центров. Строение СМ. Цереброспинальная жидкость
Анатомия и физиология ЦНС. Свойства и принципы функционирования нервных центров. Строение СМ. Цереброспинальная жидкость About animals
About animals Бионика
Бионика Мой питомец
Мой питомец Интересное животное Африки - оцелот
Интересное животное Африки - оцелот Введение в биологию. Свойства и признаки живых организмов
Введение в биологию. Свойства и признаки живых организмов Науки о природе. Природоведение. Конспект урока для учащихся
Науки о природе. Природоведение. Конспект урока для учащихся Химический состав растений
Химический состав растений Среды обитания живых организмов
Среды обитания живых организмов Биомембраны. Основные функции биомембран
Биомембраны. Основные функции биомембран Конспект урока: Экологические типы птиц, их роль в природе, жизни человека
Конспект урока: Экологические типы птиц, их роль в природе, жизни человека Ветеринарная биотехнология
Ветеринарная биотехнология Иллюзии зрения
Иллюзии зрения Роль собаки в жизни человека
Роль собаки в жизни человека Основные этапы становления и эволюции органического мира. Глобальные биотические события в история развития биосферы
Основные этапы становления и эволюции органического мира. Глобальные биотические события в история развития биосферы Физиология высшей нервной деятельности и сенсорные системы
Физиология высшей нервной деятельности и сенсорные системы Зачем публиковать данные в GBIF и на других открытых ресурсах?
Зачем публиковать данные в GBIF и на других открытых ресурсах? Растительноядные и хищные животные
Растительноядные и хищные животные Культурні рослини
Культурні рослини Физиология возбудимых тканей. Клеточная мембрана: строение, свойства, функции. Транспорт веществ через мембрану
Физиология возбудимых тканей. Клеточная мембрана: строение, свойства, функции. Транспорт веществ через мембрану Тайская кошка
Тайская кошка