- Механорецепторы кожи

Содержание
- 2. Функции Поставляет головному мозгу информацию о положении и движении тела в пространстве, о положении его отдельных
- 3. Тип воспринимаемой энергии Прикосновение механического воздействия растяжении кожи Реагируют на скорость изменения силы на болевые, и
- 4. Типы рецепторов свободные неинкапсулированные нервные окончания (длинные) свободные нервные окончания волосяных фолликулов(длинные) диски Меркеля(короткие) тельца Руффини(короткие)
- 5. Свободные неинкапсулированные нервные окончания являются самыми распространенными рецепторами кожи. Они находятся в основном в сосочковом слое
- 6. Диски (клеточные комплексы) Меркеля представляют собой видоизмененные эпителиальные клетки, с которыми образуют синапсы дендриты афферентных нейронов.
- 7. Быстро адаптирующие рецепторы Тельца Мейснера (тактильное мейснеровое тельце) представляют собой капсулу конусовидной или овальной формы. Капсула
- 8. Таким образом, Большинство тактильных рецепторов локализовано, как правило, в глубоких слоях эпидермиса и сосочковом слое кориума.
- 9. Принцип работы механорецепторов. Независимо от того, что одни рецепторы реагируют на изменение силы, вторые – на
- 10. Пути Заднестолбовой медиальный лемнисковый проводящий путь. Сенсорная информация от мех. рец. кожи -> кора БП по
- 11. Спиноцервикальный тракт(СЦТ) и волокна дорзального канатика восходят по ипсилатеральной стороне СМ до латерального цервикального ядра заднего
- 12. Что происходит в коре? Информация от нейронов специфических ядер таламуса первоначально поступает в две проекционные соматосенсорные
- 13. S1 Первая проекционная соматосенсорная область, локализованная в постцентральной извилине (первичные зоны — 1-е и 3-е поля
- 14. S2 Во вторую соматосенсорную зону коры больших полушарий (SII), расположенную в области сильвиевой борозды вблизи от
- 15. Далее… От первичных и вторичных проекционных зон коры информация от тактильных рецепторов поступает в передние (фронтальные)
- 17. Скачать презентацию

Функции
Поставляет головному мозгу информацию о положении и движении тела в пространстве,
Функции
Поставляет головному мозгу информацию о положении и движении тела в пространстве,

Тип воспринимаемой энергии
Прикосновение
механического воздействия
растяжении кожи
Реагируют на скорость изменения силы
на болевые, и
Тип воспринимаемой энергии
Прикосновение
механического воздействия
растяжении кожи
Реагируют на скорость изменения силы
на болевые, и

Типы рецепторов
свободные неинкапсулированные нервные окончания (длинные)
свободные нервные окончания волосяных фолликулов(длинные)
диски
Типы рецепторов
свободные неинкапсулированные нервные окончания (длинные)
свободные нервные окончания волосяных фолликулов(длинные)
диски

Свободные неинкапсулированные нервные окончания являются самыми распространенными рецепторами кожи. Они находятся
Свободные неинкапсулированные нервные окончания являются самыми распространенными рецепторами кожи. Они находятся

Диски (клеточные комплексы) Меркеля представляют собой видоизмененные эпителиальные клетки, с которыми
Диски (клеточные комплексы) Меркеля представляют собой видоизмененные эпителиальные клетки, с которыми

Быстро адаптирующие рецепторы
Тельца Мейснера (тактильное мейснеровое тельце) представляют собой капсулу конусовидной или
Быстро адаптирующие рецепторы
Тельца Мейснера (тактильное мейснеровое тельце) представляют собой капсулу конусовидной или

Таким образом,
Большинство тактильных рецепторов локализовано, как правило, в глубоких слоях эпидермиса
Таким образом,
Большинство тактильных рецепторов локализовано, как правило, в глубоких слоях эпидермиса

Принцип работы механорецепторов.
Независимо от того, что одни рецепторы реагируют на изменение
Принцип работы механорецепторов.
Независимо от того, что одни рецепторы реагируют на изменение
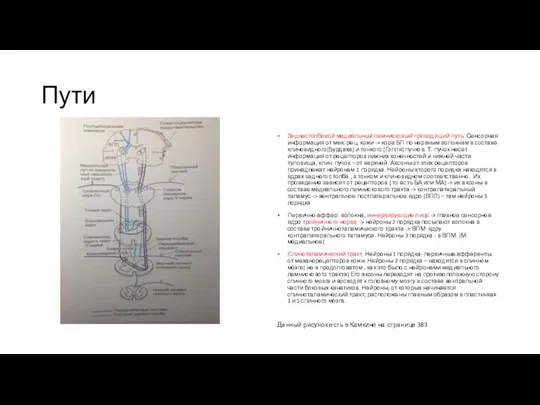
Пути
Заднестолбовой медиальный лемнисковый проводящий путь. Сенсорная информация от мех. рец. кожи
Пути
Заднестолбовой медиальный лемнисковый проводящий путь. Сенсорная информация от мех. рец. кожи

Спиноцервикальный тракт(СЦТ) и волокна дорзального канатика восходят по ипсилатеральной стороне СМ
Спиноцервикальный тракт(СЦТ) и волокна дорзального канатика восходят по ипсилатеральной стороне СМ

Что происходит в коре?
Информация от нейронов специфических ядер таламуса первоначально поступает
Что происходит в коре?
Информация от нейронов специфических ядер таламуса первоначально поступает

S1
Первая проекционная соматосенсорная область, локализованная в постцентральной извилине (первичные зоны —
S1
Первая проекционная соматосенсорная область, локализованная в постцентральной извилине (первичные зоны —

S2
Во вторую соматосенсорную зону коры больших полушарий (SII), расположенную в области
S2
Во вторую соматосенсорную зону коры больших полушарий (SII), расположенную в области

Далее…
От первичных и вторичных проекционных зон коры информация от тактильных рецепторов
Далее…
От первичных и вторичных проекционных зон коры информация от тактильных рецепторов
 General characteristics of 6 Kingdoms
General characteristics of 6 Kingdoms Биологически активные низкомолекулярные вещества
Биологически активные низкомолекулярные вещества Әр түрлі мөлшердегі минералды тыңайтқыштардың қант қызылшасының өнімділігіне әсерін анықтау барысындағы суреттер
Әр түрлі мөлшердегі минералды тыңайтқыштардың қант қызылшасының өнімділігіне әсерін анықтау барысындағы суреттер Растительная клетка. Лекция 1
Растительная клетка. Лекция 1 Актинидия. Особенности
Актинидия. Особенности Многообразие ракообразных
Многообразие ракообразных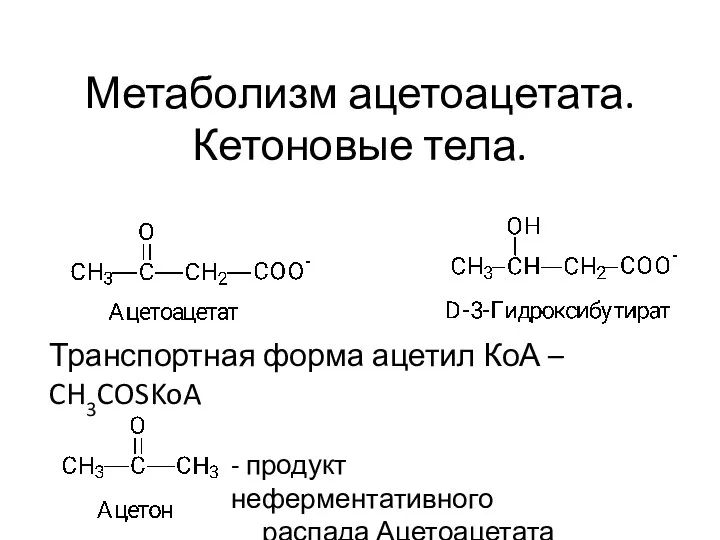 Кетоновые тела. Тема 10
Кетоновые тела. Тема 10 Лист. Морфология
Лист. Морфология Строение растительной клетки
Строение растительной клетки Генеративні органи рослини
Генеративні органи рослини Функциональная асимметрия мозга. Лекция 11
Функциональная асимметрия мозга. Лекция 11 Методы микроскопии
Методы микроскопии Самое редкое животное Африки - Окапи
Самое редкое животное Африки - Окапи Презентация, конспект урока, карточки - задания к уроку биологии для 6 класса на тему Фотосинтез.
Презентация, конспект урока, карточки - задания к уроку биологии для 6 класса на тему Фотосинтез. ПрезентацияТурнир по биологии в 8 классе.
ПрезентацияТурнир по биологии в 8 классе. Тема 12.Навоз, воздух, вода
Тема 12.Навоз, воздух, вода Лист — боковой орган побега
Лист — боковой орган побега Фотосинтез. История открытия
Фотосинтез. История открытия Миология. Мышцы тела человека
Миология. Мышцы тела человека Рослини ендеміки Австралії
Рослини ендеміки Австралії Деление клетки – митоз, мейоз
Деление клетки – митоз, мейоз Животные Республики Башкортостан
Животные Республики Башкортостан Физиология растений. Лекция 2
Физиология растений. Лекция 2 Красная книга Сызранского района
Красная книга Сызранского района В Буграх растут деревья и кустарники
В Буграх растут деревья и кустарники Сон и его значение
Сон и его значение Половое поведение животных
Половое поведение животных Ароматерапия. Химия эфирных масел
Ароматерапия. Химия эфирных масел