- Метаболизм белков, пептидов, аминокислот. Лекция 19-21

Содержание
- 2. Общая характеристика биосинтеаз белка В биосинтезе белка участвует очень большое число белков, иРНК, тРНК, рРНК в
- 3. Для расшифровки нуклеотидной последовательности необходим генетический код Он устанавливает соответствие между нуклеотидной последовательностью иРНК и полипептидной
- 4. Каждому кодону (61 из 64) соответствует строго определенная аминокислота
- 5. Трансляция иРНК начинается с инициирующего триплета – АУГ Далее расшифровывается каждый последующий триплет в направлении от
- 6. 20 стандартным аминокислотам соответствует 61 кодон Таким образом, почти каждой стандартной аминокислоте соответствует несколько кодонов Т.о.
- 7. Генетический код
- 8. Свойства генетического кода Триплетность Кодон состоит из последовательности, состоящей из трех нуклеотидов. Специфичность (однозначность) Каждому кодону
- 9. Вырожденность Каждый из 61 смысловых триплетов кодирует включение в белок одну из 20 стандартных аминокислот. Включение
- 10. Неперекрываемость кода Триплеты читаются последовательно не перекрываясь без каких либо пропусков.
- 11. Универсальность кода Код универсален, т.е. смысл кодов одинаков для всех организмов Существует исключение – митохондриальная мРНК
- 12. Стадии биосинтеза белка (трансляция) Активация аминокислот На этой стадии каждая из 20 аминокислот присоединяется к определенной
- 14. Суммарная реакция активации аминокислоты выглядит так:
- 15. Аминоацил-тРНК-синтетазы обладают корректирующей активностью – способны вырезать неправильно присоединенный остаток аминокислоты.
- 16. Инициация белкового синтеза Процессы трансляции эукариотической иРНК и прокариотической иРНК в общих чертах сходны. Инициирующей аа-тРНК
- 17. Инициация начинается с присоединения к малой субъединице рибосомы иРНК и первой аа-тРНК, антикодон которой комплементарен инициирующему
- 19. Для осуществления инициации трансляции необходима энергия. Ее поставляет ГТФ. Энергия высвобождается при гидролизе ГТФ до ГДФ
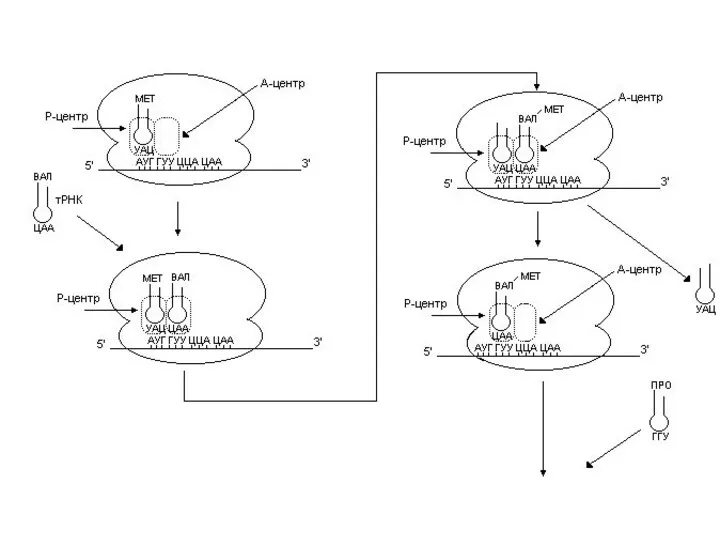
- 20. Элонгация белкового синтеза В процессе элонгации происходит наращивание полипептидной цепи. В А-центре происходит связывание аа-тРНК, антикодон
- 21. Далее рибосома перемещается на один кодон в направлении от 5’- к 3’-концу мРНК. Дипептидил-тРНК из А-центра
- 22. Теперь рибосома готова к новому циклу элонгации. Дипептид с дипептидил-тРНК переносится на следующую аминоацил-тРНК, находящуюся в
- 24. Пептидилтрансферазная реакция:
- 25. Терминация Терминация белового синтеза наступает, как только в А-центре окажется один из терминирующих кодонов: УАГ, УГА,
- 26. ПОСТТРАНСЛЯЦИОННЫЕ МОДИФИКАЦИИ БЕЛКОВ В процессе трансляции участвуют 20 протеиногенных аминокислот. К Несмотря на это в составе
- 27. В настоящее время известно более двухсот различных вариантов ПТМ белков. Среди них выделяют: модификации функциональных групп
- 28. химические перестройки основы аминокислотных остатков (формирование межцепочечных сшивок в молекулах коллагена и др.); ограниченный (частичный) протеолиз
- 29. Фосфорилирование и дефосфорилирование радикалов аминокислотных остатков. ПК – протеинкиназа ФПФазы –фосфопротеинфосфатаза.
- 30. Реакциями фосфорилирования и дефосфорилирования регулируются многочисленные ферменты, участвующие в метаболизме различных вещест, контролируются функции белков цитоскелета,
- 31. Гликозилирование белков. В процессе гликозилирования белков к ним ковалентно присоединяется углеводный компонент. Гликозилированные белки называются гликопротеинами.
- 32. Протеолитические ферменты (протеазы, протеиназы) Протеазы — ферменты класса гидролаз, расщепляющие пептидные связи между аминокислотами в белках
- 33. Протеазы разделяют на шесть групп, в зависимости от строения активного центра: сериновые; в активном центре этих
- 34. ОГРАНИЧЕННЫЙ ПРОТЕОЛИЗ БЕЛКОВ Ограниченный протеолиз – расщепление одной или нескольких пептидных связей в молекуле белка посредством
- 35. Предшественником пепсина является пепсиноген.
- 36. Образование активной формы химотрипсина из химотрипсиногена:
- 37. Процессинг инсулина
- 38. Диссимиляция белков При окислении 1 г белка выделяется 17,2 кДж (4,1 ккал) энергии. Белки редко использует
- 40. Бывают - незаменимые аминокислоты (лейцин, изолейцин, валин, фенилаланин, триптофан, треонин, лизин, метионин), - заменимые аминокислоты. -
- 41. Катаболизм аминокислот Катаболизм большинства аминокислот начинается с отщепления α-аминогруппы. Аминокислота теряет аминогруппу в результате дезаминирования или
- 42. Дезаминирование аминокислот
- 43. Переаминирование аминокислот Переаминирование или трансаминирование —ферментативная реакция обратимого переноса аминогруппы с аминокислоты на кетокислоту . Ферменты,
- 45. Аминокислоты могут подвергаться и другим превращениям. 10 аминокислот (ала, тре, гли, сер, цис, фен, три, тир,
- 46. 2 аминокислоты (фен, тир) – в фумарат 2 аминокислоты (асп, асн) – в оксалоацетат Все конечные
- 47. Биосинтез аминокислот в тканях. В организме человека возможен синтез заменимых аминокислот. Основными путями образования заменимых аминокислот
- 48. 2) восстановительное аминирование α-кетокислот, 3) синтез с участием незаменимых аминокислот. Заменимая аминокислота тирозин может образоваться из
- 49. Цикл мочевины Деградация аминокислот происходит преимущественно в печени. При этом освобождается аммиак. Значительные количества аммиака образуются
- 50. У наземных позвоночных аммиак превращается в мочевину (уреотелические организмы) Живущие в воде животные выделяют аммиак непосредственно
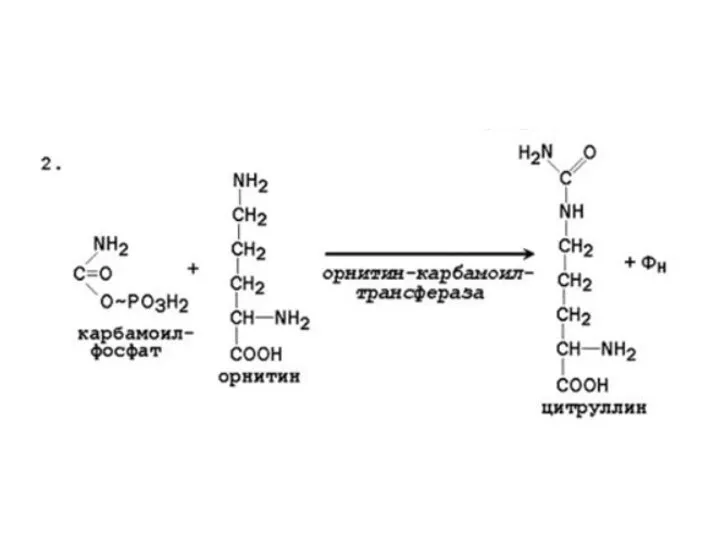
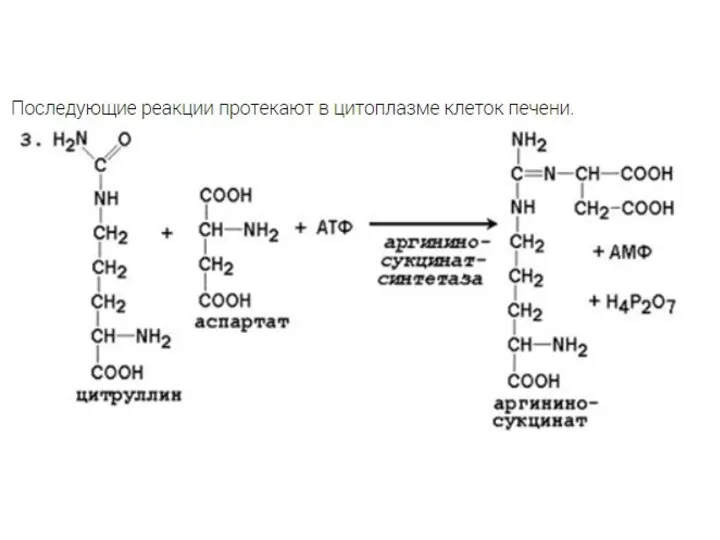
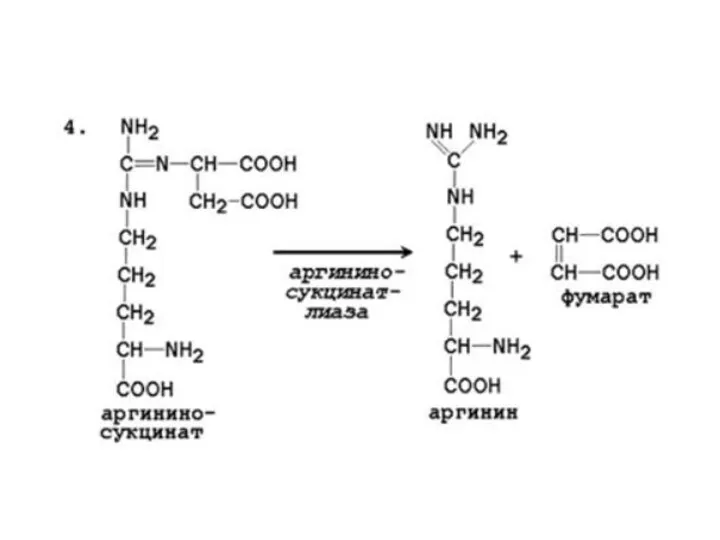
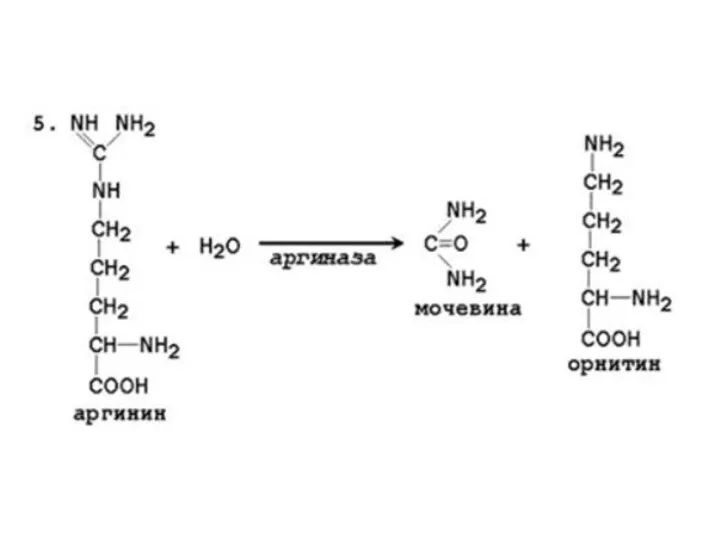
- 51. Мочевина синтезируется в орнитиновом цикле, протекающем в клетках печени. Две начальные реакции биосинтеза мочевины происходят в
- 57. Скачать презентацию

Общая характеристика биосинтеаз белка
В биосинтезе белка участвует очень большое число
Общая характеристика биосинтеаз белка
В биосинтезе белка участвует очень большое число

Для расшифровки нуклеотидной последовательности необходим генетический код
Он устанавливает соответствие между нуклеотидной
Для расшифровки нуклеотидной последовательности необходим генетический код
Он устанавливает соответствие между нуклеотидной

Каждому кодону (61 из 64) соответствует строго определенная аминокислота

Трансляция иРНК начинается с инициирующего триплета – АУГ
Далее расшифровывается каждый
Трансляция иРНК начинается с инициирующего триплета – АУГ
Далее расшифровывается каждый

20 стандартным аминокислотам соответствует 61 кодон
Таким образом, почти каждой стандартной аминокислоте
20 стандартным аминокислотам соответствует 61 кодон
Таким образом, почти каждой стандартной аминокислоте
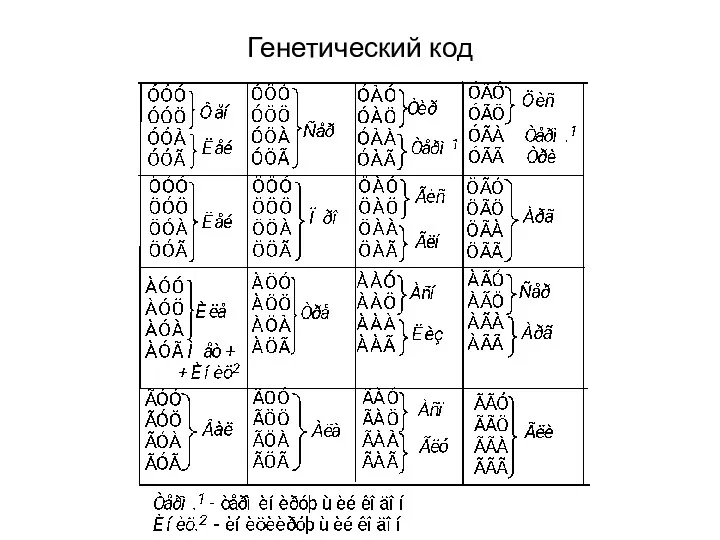
Генетический код
Генетический код

Свойства генетического кода
Триплетность
Кодон состоит из последовательности, состоящей из трех нуклеотидов.
Специфичность (однозначность)
Свойства генетического кода
Триплетность
Кодон состоит из последовательности, состоящей из трех нуклеотидов.
Специфичность (однозначность)

Вырожденность
Каждый из 61 смысловых триплетов кодирует включение в белок одну из
Вырожденность
Каждый из 61 смысловых триплетов кодирует включение в белок одну из

Неперекрываемость кода
Триплеты читаются последовательно не перекрываясь без каких либо пропусков.
Неперекрываемость кода
Триплеты читаются последовательно не перекрываясь без каких либо пропусков.

Универсальность кода
Код универсален, т.е. смысл кодов одинаков для всех организмов
Существует
Универсальность кода
Код универсален, т.е. смысл кодов одинаков для всех организмов
Существует

Стадии биосинтеза белка (трансляция)
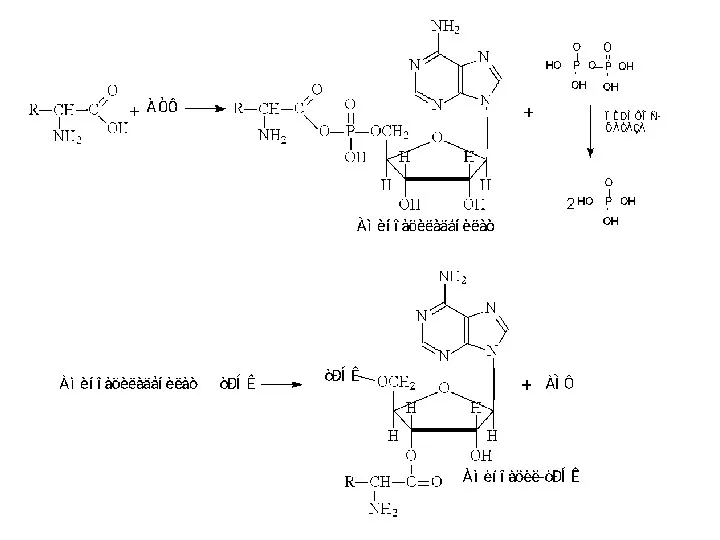
Активация аминокислот
На этой стадии каждая из 20 аминокислот
Стадии биосинтеза белка (трансляция)
Активация аминокислот
На этой стадии каждая из 20 аминокислот
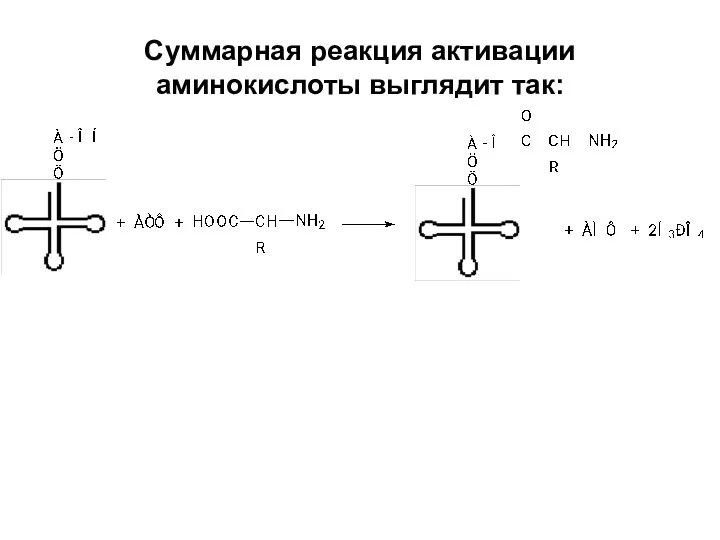
Суммарная реакция активации аминокислоты выглядит так:
Суммарная реакция активации аминокислоты выглядит так:

Аминоацил-тРНК-синтетазы обладают корректирующей активностью – способны вырезать неправильно присоединенный остаток аминокислоты.
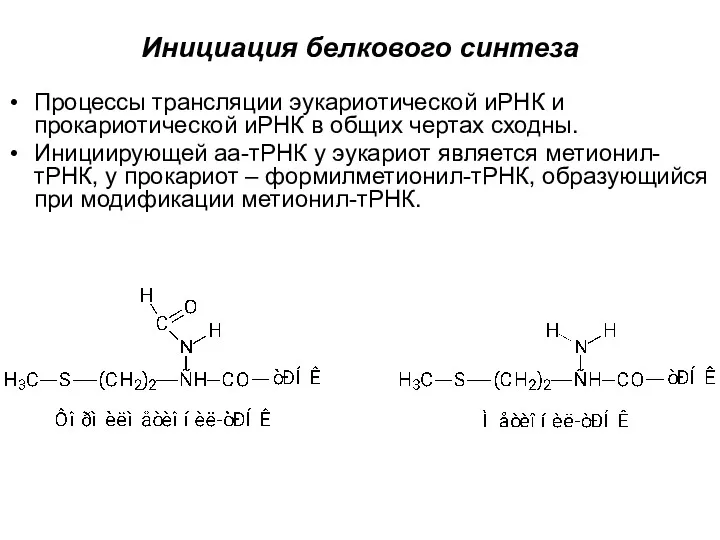
Инициация белкового синтеза
Процессы трансляции эукариотической иРНК и прокариотической иРНК в общих
Инициация белкового синтеза
Процессы трансляции эукариотической иРНК и прокариотической иРНК в общих

Инициация начинается с присоединения к малой субъединице рибосомы иРНК и первой
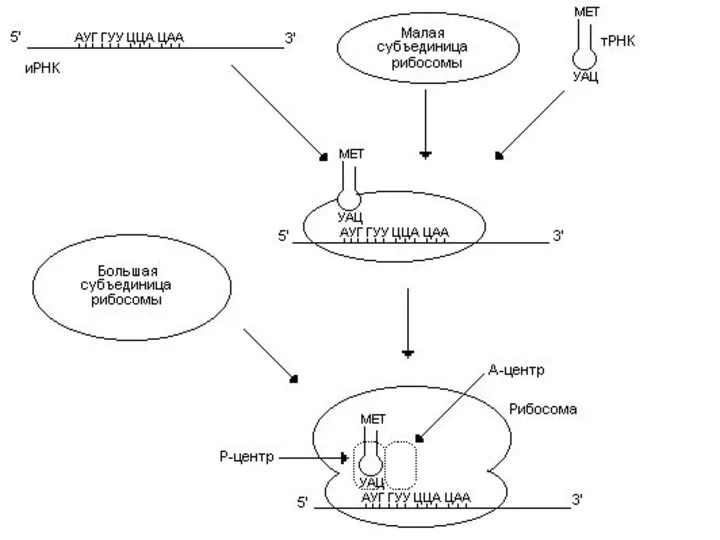

Для осуществления инициации трансляции необходима энергия. Ее поставляет ГТФ. Энергия высвобождается
Для осуществления инициации трансляции необходима энергия. Ее поставляет ГТФ. Энергия высвобождается

Элонгация белкового синтеза
В процессе элонгации происходит наращивание полипептидной цепи.
В А-центре
Элонгация белкового синтеза
В процессе элонгации происходит наращивание полипептидной цепи.
В А-центре

Далее рибосома перемещается на один кодон в направлении от 5’- к
Далее рибосома перемещается на один кодон в направлении от 5’- к

Теперь рибосома готова к новому циклу элонгации.
Дипептид с дипептидил-тРНК переносится
Дипептид с дипептидил-тРНК переносится
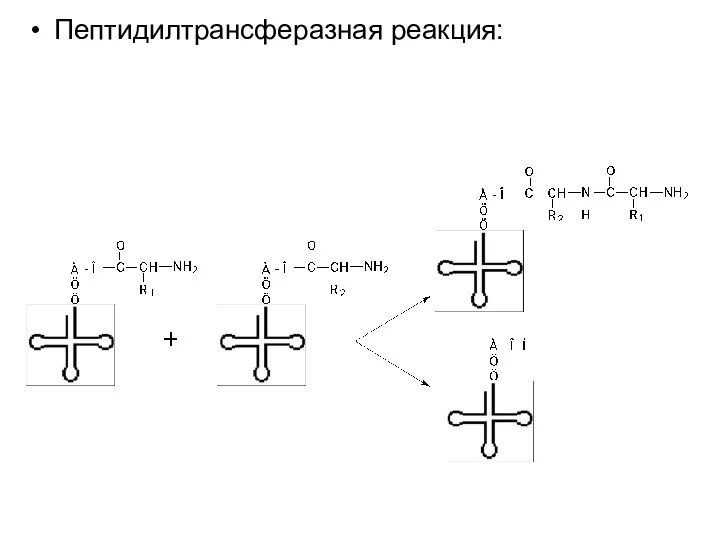
Пептидилтрансферазная реакция:
Пептидилтрансферазная реакция:

Терминация
Терминация белового синтеза наступает, как только в А-центре окажется один из
Терминация
Терминация белового синтеза наступает, как только в А-центре окажется один из

ПОСТТРАНСЛЯЦИОННЫЕ МОДИФИКАЦИИ БЕЛКОВ
В процессе трансляции участвуют 20 протеиногенных аминокислот.
К Несмотря
В процессе трансляции участвуют 20 протеиногенных аминокислот.
К Несмотря

В настоящее время известно более двухсот различных вариантов ПТМ белков. Среди
В настоящее время известно более двухсот различных вариантов ПТМ белков. Среди

химические перестройки основы аминокислотных остатков (формирование межцепочечных сшивок в молекулах коллагена
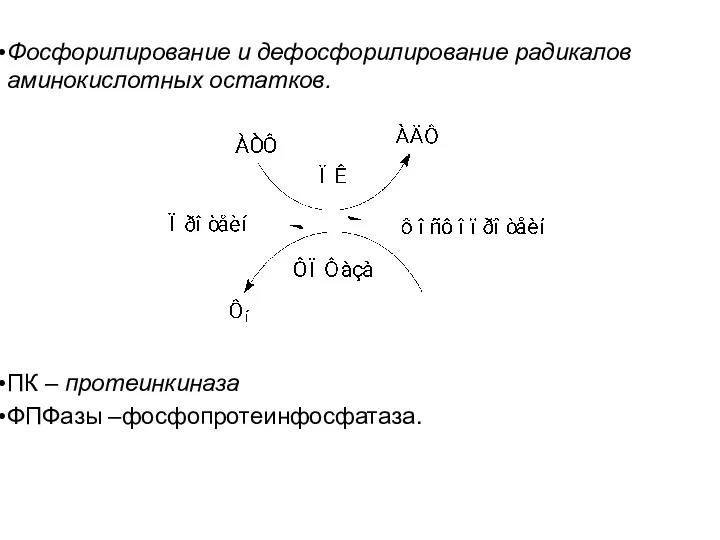
Фосфорилирование и дефосфорилирование радикалов аминокислотных остатков.
ПК – протеинкиназа
ФПФазы –фосфопротеинфосфатаза.
Фосфорилирование и дефосфорилирование радикалов аминокислотных остатков.
ПК – протеинкиназа
ФПФазы –фосфопротеинфосфатаза.

Реакциями фосфорилирования и дефосфорилирования
регулируются многочисленные ферменты, участвующие в метаболизме различных
Реакциями фосфорилирования и дефосфорилирования
регулируются многочисленные ферменты, участвующие в метаболизме различных

Гликозилирование белков.
В процессе гликозилирования белков к ним ковалентно присоединяется углеводный
Гликозилирование белков.
В процессе гликозилирования белков к ним ковалентно присоединяется углеводный

Протеолитические ферменты (протеазы, протеиназы)
Протеазы — ферменты класса гидролаз, расщепляющие пептидные связи между аминокислотами в
Протеолитические ферменты (протеазы, протеиназы)
Протеазы — ферменты класса гидролаз, расщепляющие пептидные связи между аминокислотами в

Протеазы разделяют на шесть групп, в зависимости от строения активного центра:
сериновые;
Протеазы разделяют на шесть групп, в зависимости от строения активного центра:
сериновые;

ОГРАНИЧЕННЫЙ ПРОТЕОЛИЗ БЕЛКОВ
Ограниченный протеолиз – расщепление одной или нескольких пептидных связей
ОГРАНИЧЕННЫЙ ПРОТЕОЛИЗ БЕЛКОВ
Ограниченный протеолиз – расщепление одной или нескольких пептидных связей
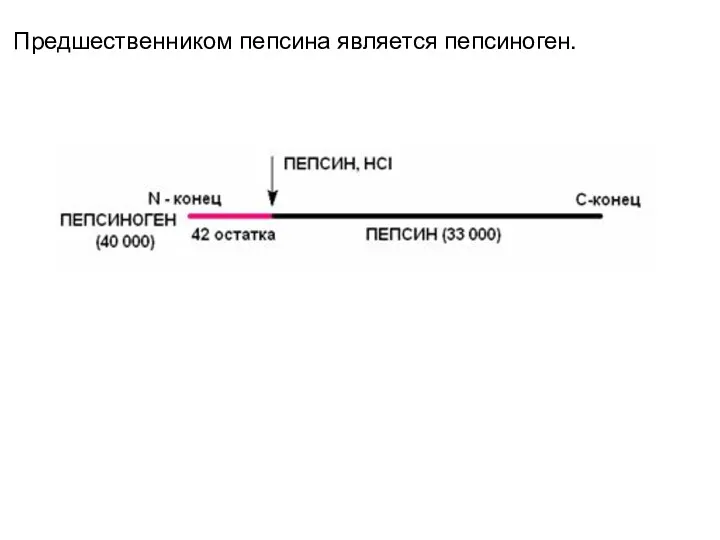
Предшественником пепсина является пепсиноген.
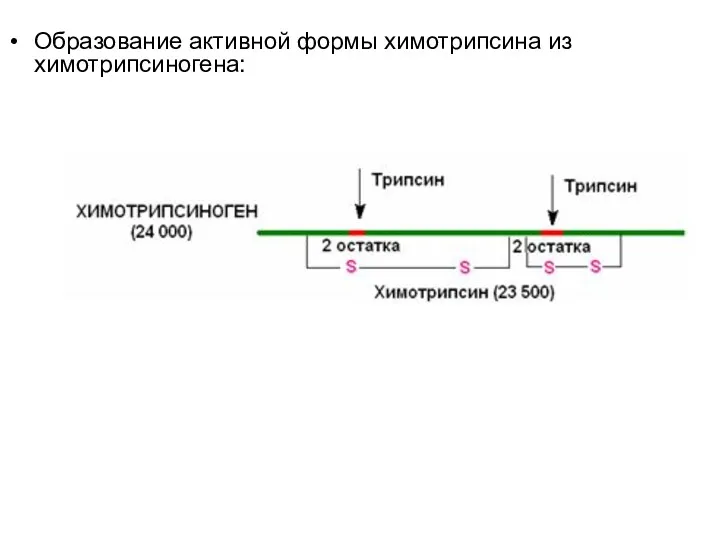
Образование активной формы химотрипсина из химотрипсиногена:
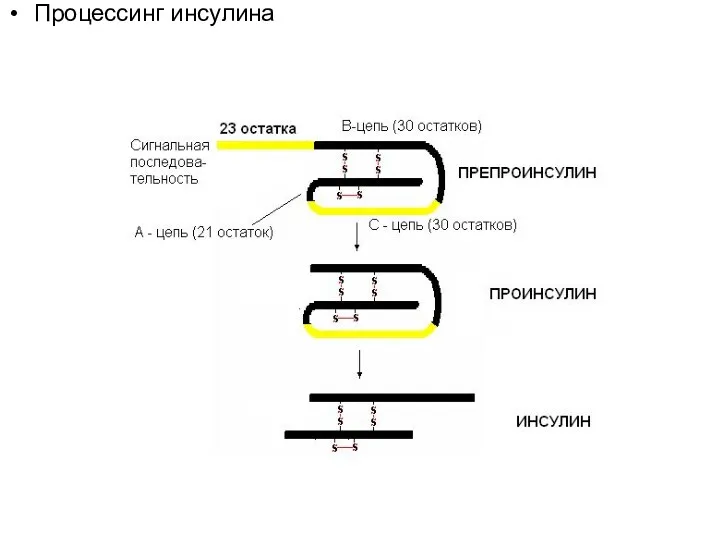
Процессинг инсулина
Процессинг инсулина

Диссимиляция белков
При окислении 1 г белка выделяется 17,2 кДж (4,1 ккал) энергии.
Белки редко использует в больших
Диссимиляция белков
При окислении 1 г белка выделяется 17,2 кДж (4,1 ккал) энергии.
Белки редко использует в больших
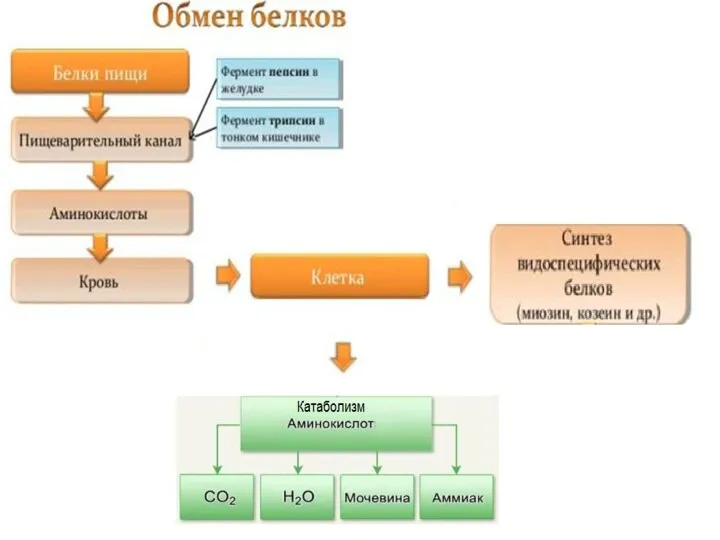

Бывают
- незаменимые аминокислоты (лейцин, изолейцин, валин, фенилаланин, триптофан, треонин, лизин, метионин),
-
Бывают
- незаменимые аминокислоты (лейцин, изолейцин, валин, фенилаланин, триптофан, треонин, лизин, метионин),
-

Катаболизм аминокислот
Катаболизм большинства аминокислот начинается с отщепления α-аминогруппы.
Аминокислота теряет аминогруппу
Катаболизм аминокислот
Катаболизм большинства аминокислот начинается с отщепления α-аминогруппы.
Аминокислота теряет аминогруппу
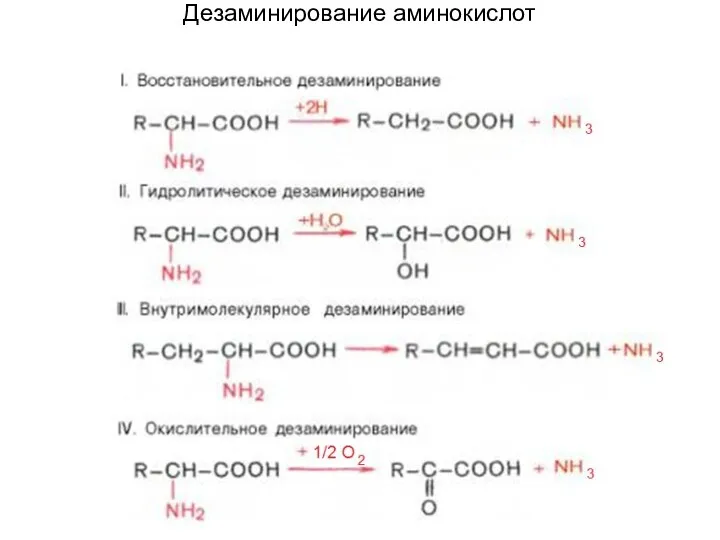
Дезаминирование аминокислот
Дезаминирование аминокислот
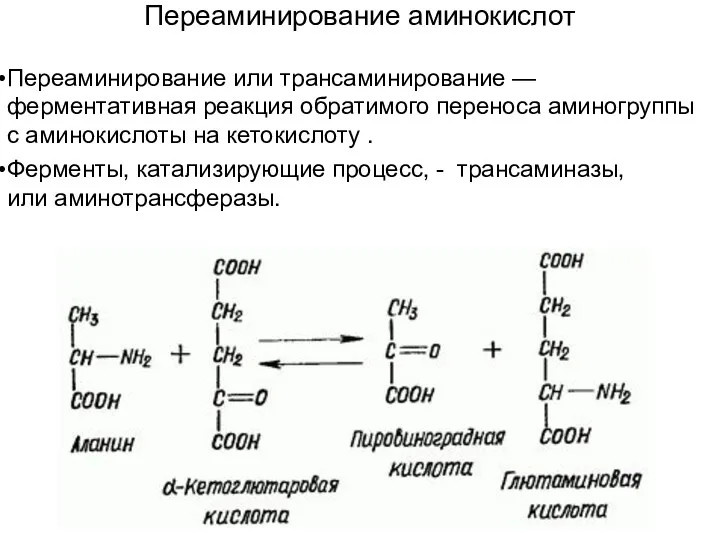
Переаминирование аминокислот
Переаминирование или трансаминирование —ферментативная реакция обратимого переноса аминогруппы с аминокислоты на кетокислоту .
Ферменты, катализирующие процесс,
Переаминирование аминокислот
Переаминирование или трансаминирование —ферментативная реакция обратимого переноса аминогруппы с аминокислоты на кетокислоту .
Ферменты, катализирующие процесс,

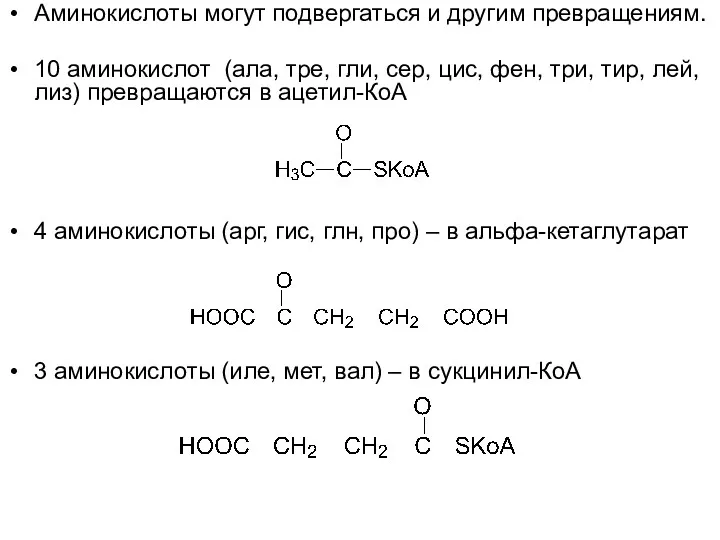
Аминокислоты могут подвергаться и другим превращениям.
10 аминокислот (ала, тре, гли, сер,
Аминокислоты могут подвергаться и другим превращениям.
10 аминокислот (ала, тре, гли, сер,
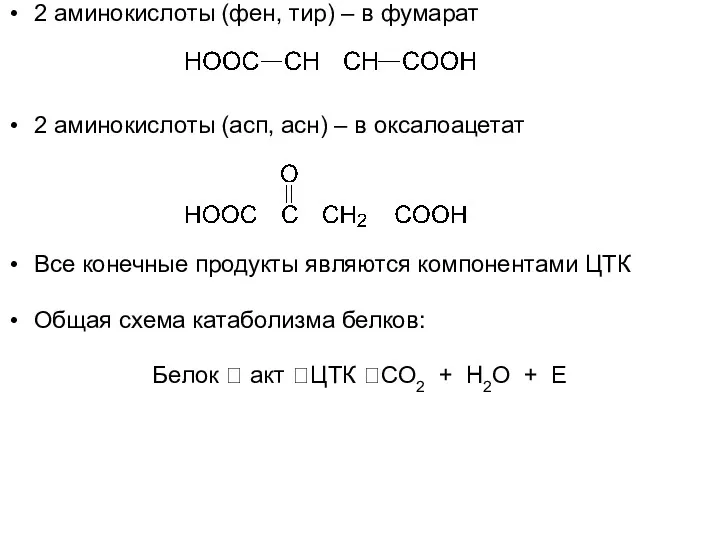
2 аминокислоты (фен, тир) – в фумарат
2 аминокислоты (асп, асн) –
2 аминокислоты (фен, тир) – в фумарат
2 аминокислоты (асп, асн) –

Биосинтез аминокислот в тканях.
В организме человека возможен синтез заменимых аминокислот.
Основными
Биосинтез аминокислот в тканях.
В организме человека возможен синтез заменимых аминокислот.
Основными
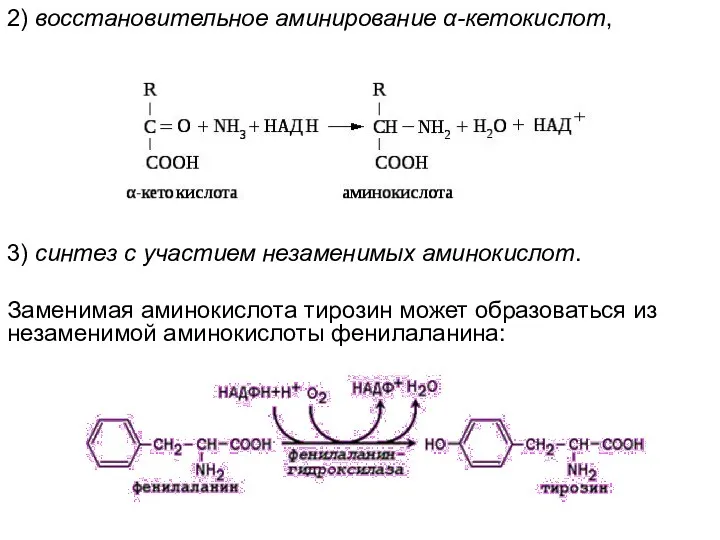
2) восстановительное аминирование α-кетокислот,
3) синтез с участием незаменимых аминокислот.
Заменимая аминокислота тирозин может образоваться
2) восстановительное аминирование α-кетокислот,
3) синтез с участием незаменимых аминокислот.
Заменимая аминокислота тирозин может образоваться

Цикл мочевины
Деградация аминокислот происходит преимущественно в печени. При этом освобождается аммиак.
Цикл мочевины
Деградация аминокислот происходит преимущественно в печени. При этом освобождается аммиак.

У наземных позвоночных аммиак превращается в мочевину (уреотелические организмы)
Живущие в воде животные
У наземных позвоночных аммиак превращается в мочевину (уреотелические организмы)
Живущие в воде животные
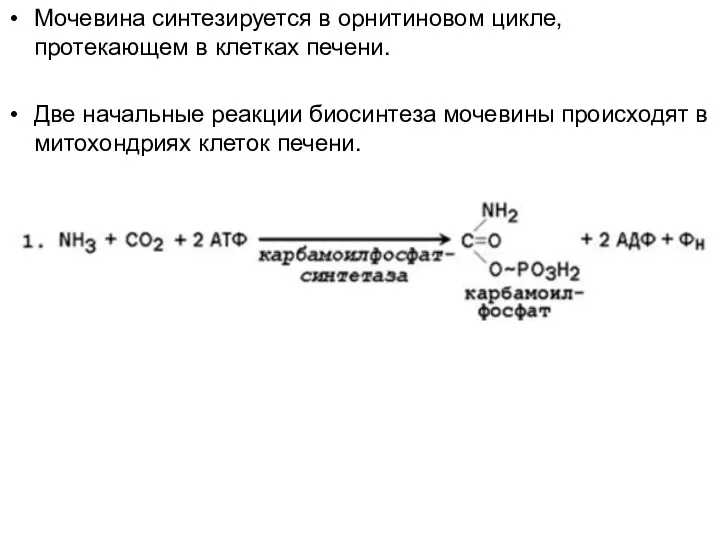
Мочевина синтезируется в орнитиновом цикле, протекающем в клетках печени.
Две начальные
Мочевина синтезируется в орнитиновом цикле, протекающем в клетках печени.
Две начальные
 Удивительный мир живой природы
Удивительный мир живой природы Эволюционный процесс
Эволюционный процесс Энергетический метаболизм микробов. Роль генома в метаболической активности микроорганизмов
Энергетический метаболизм микробов. Роль генома в метаболической активности микроорганизмов Урок на тему :Совместная жизнь видов в биогеоценозе.
Урок на тему :Совместная жизнь видов в биогеоценозе. Основы микробиологии и биотехнологии
Основы микробиологии и биотехнологии Красноухая черепаха
Красноухая черепаха Нәсілдердің пайда болуы
Нәсілдердің пайда болуы Жизнь и творчество Виктора Драгунского
Жизнь и творчество Виктора Драгунского A Tour of the Cell
A Tour of the Cell Разнообразие клеток
Разнообразие клеток Невидимые нити. Окружающий мир 2 класс
Невидимые нити. Окружающий мир 2 класс Гербарии растений, содержащих флавоноиды I
Гербарии растений, содержащих флавоноиды I Собака – верный друг
Собака – верный друг Приспособленность организмов к определенной среде обитания
Приспособленность организмов к определенной среде обитания Удивительные животные
Удивительные животные Лишайники. Внешнее и внутреннее строение
Лишайники. Внешнее и внутреннее строение Лишайники. Симбиоз
Лишайники. Симбиоз изменчивость
изменчивость В краю родном. Викторина о природе Ростовской области
В краю родном. Викторина о природе Ростовской области Строение спинного мозга человека
Строение спинного мозга человека Рентгеноанатомия черепа. Обозначьте кости мозгового черепа
Рентгеноанатомия черепа. Обозначьте кости мозгового черепа Первобытный человек
Первобытный человек Где и когда жили динозавры?
Где и когда жили динозавры? Вирусы. (11 класс)
Вирусы. (11 класс) Ткани, органы и системы органов
Ткани, органы и системы органов Значення мікробіології в практичній діяльності лікаря-стоматолога
Значення мікробіології в практичній діяльності лікаря-стоматолога Физиология возбудимых клеток. Медиаторы (лекция № 5)
Физиология возбудимых клеток. Медиаторы (лекция № 5) Сердце. Анатомия сердца
Сердце. Анатомия сердца