- Метаболизм липидов

Содержание
- 2. В состав мембран входят только ФОСФОЛИПИДЫ (ФЛ), ГЛИКОЛИПИДЫ (ГЛ) и ХОЛЕСТЕРИН (ХС).
- 6. Метаболизм липидов
- 8. Жирные кислоты, входящие в состав организма человека, имеют общие черты строения: 1. Чётное число атомов углерода.
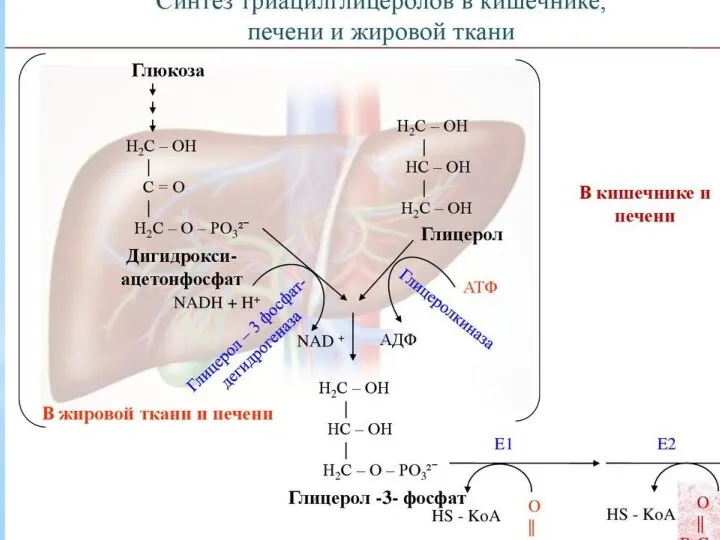
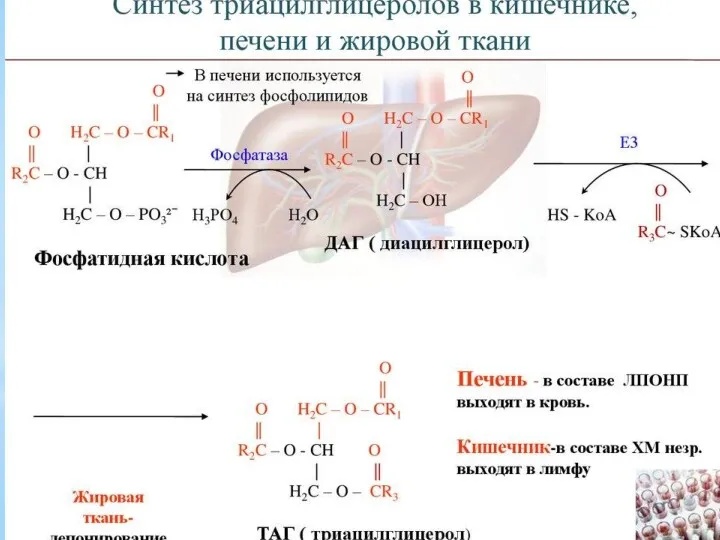
- 12. Триацилглицеролы являются формой хранения энергетического и строительного материала. В организме человека для построения их структур используются
- 15. Представители сложных липидов – глицерофосфолипиды важнейшие компоненты клеточных мембран. Эти липиды содержат глицерин, связанный с двумя
- 17. Глицерофосфолипиды – амфипатические соединения, один конец молекулы которых гирофобен, а другой гидрофилен. Следствием такой структуры является
- 18. В транспортных формах липидов (хиломикроны, липопротеиды очень низкой плотности) мембрана однослойная и гидрофобные части молекул обращены
- 19. Фосфатидилхолин является уникальной молекулой с обширным перечнем функций. В легочных альвеолах это соединение входит в состав
- 20. Органические осмолиты повышают осмоляльность внутриклеточного раствора до уровня внеклеточной жидкости, препятствуя потере клеткой воды. Эпителиоциты этого
- 21. Эйкозаноиды: простагландины, лейкотриены, тромбоксаны и простациклины, образуются во многих клетках в ответ на внешний стимул, изменяют
- 22. Интересным представителем мембранных липидов является фосфатидилинозитол. Рис. 4. Фосфатидилинозитол
- 23. Липид встречается во внутреннем слое цитоплазматической мембраны большинства клеток организма. Инозитол, входящий в состав фосфолипида является
- 24. Инозитол-1,4,5-трифосфат связывается с рецептором гладкого эндоплазматического ретикулума, что приводит к высвобождению из цистерн ретикулума ионов Са2+.
- 25. Внутренняя мембрана митохондрии содержит кардиолипин. Этот фосфолипид впервые был выделен из сердечной мышцы, где его больше
- 26. Рис. 5. Кардиолипин
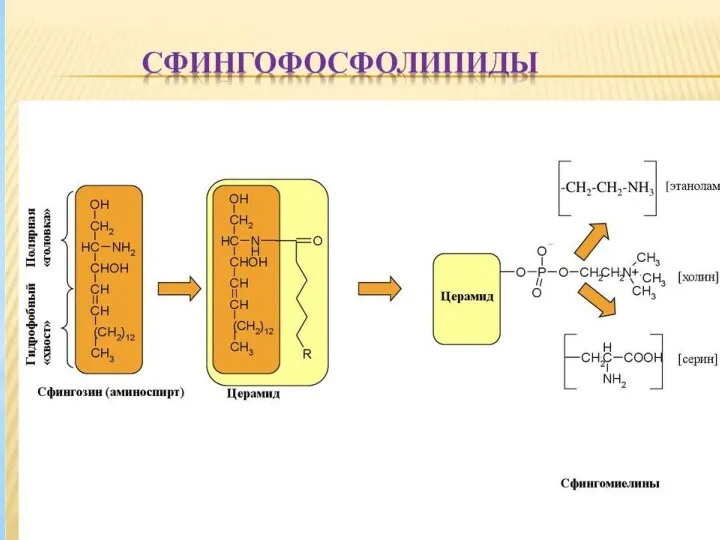
- 27. Сфингомиелины Сфингомиелины – мембранные липиды, состоящие из аминоспирта сфингозина, жирной кислоты, остатка фосфорной кислоты и холина,
- 29. Гликолипиды Существует два класса гликолипидов: цереброзиды и ганглиозиды. Оба класса липидов встречаются в цитоплазматических мембранах разных
- 30. Рис. 7. Глюкоцереброзид
- 31. Холестерин Холестерин спирт, в основе структуры которого гетероцикл – циклопентапергидрофенантрен. Входит в состав цитоплазматических мембран, регулируя
- 32. Рис. Холестерин
- 33. Пищеварение липидов в ЖКТ В пище человека могут содержаться большинство липидов приведенных в классификации, однако наиболее
- 34. У взрослых людей переваривание жира идет только в кишечнике по схеме: «выделение желчи - эмульгирование жира
- 35. Собственно переваривание жиров – это гидролиз сложноэфирных связей. Существует три фермента: Язычная липаза. Вырабатывается клетками слизистой
- 36. Хотя в глубине пищевого комка, где рН меняется не сразу, ферменты могут ещё действовать. Основное пищеварение
- 37. В результате образуется активная форма и пентапептид энтеростатин следующего состава – Вал-Про-Асп-Про-Арг. Колипаза образует активный комплекс
- 47. Жирные кислоты С6-С12 поступают в кровеносные капилляры, а β-моноацилглицерол (β-МГ) включается в синтез триацилглицеролов, участвующих в
- 48. Хиломикроны поступают в лимфатические капилляры, из которых через грудной проток в системный кровоток. В крови осуществляется
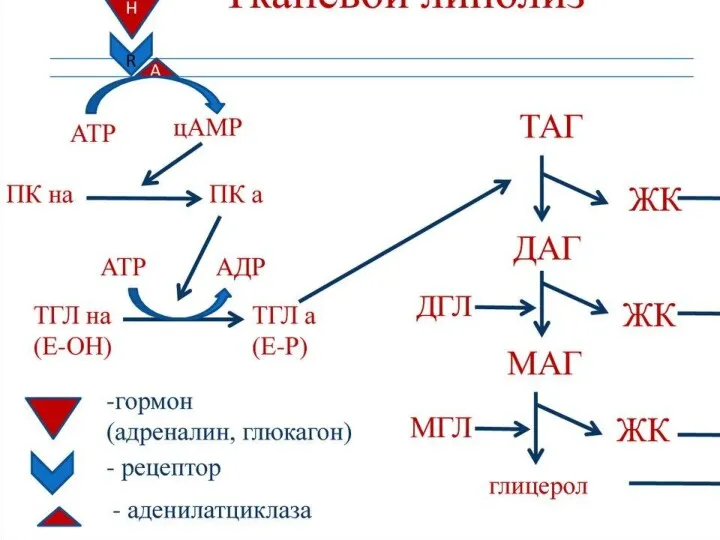
- 69. Катаболизм триацилглицеролов Триацилглицеролы (нейтральные жиры) – важный резерв энергетического и пластического материала для организма. Основные их
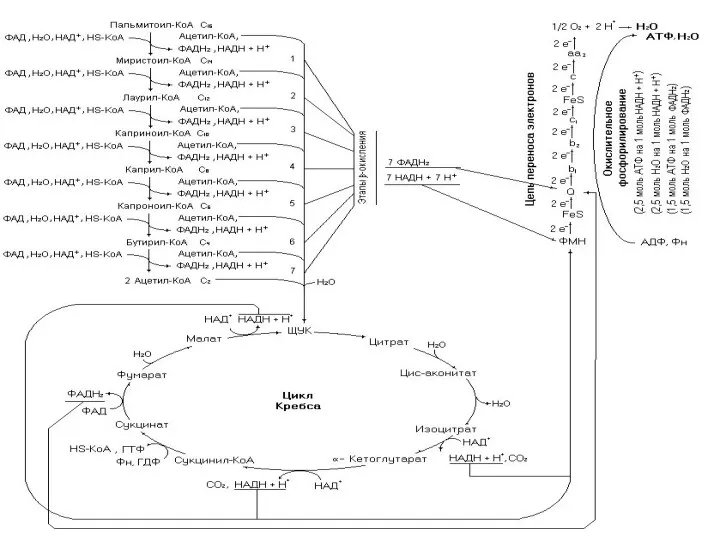
- 89. β-окисление пальмитиновой кислоты
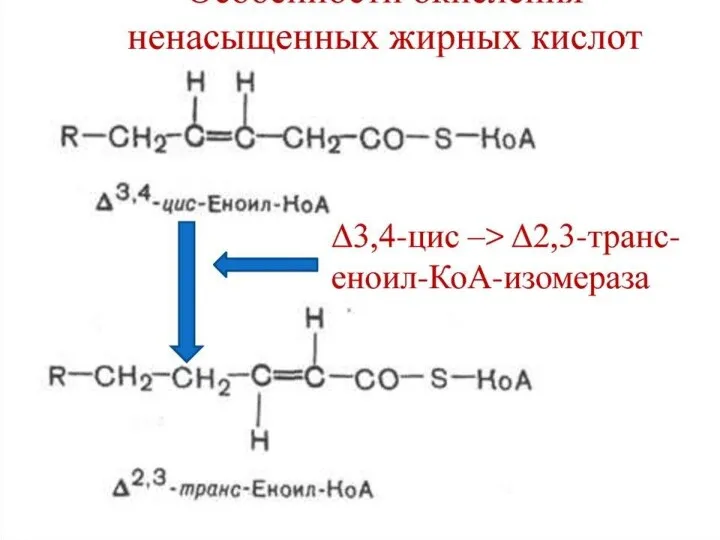
- 99. Биоэнергетика окисления предельных и непредельных жирных кислот с четным числом углеродных атомов
- 100. Окисление жирных кислот осуществляется с целью трансформации выделившейся при этом энергии в энергию макроэргических связей АТФ.
- 101. Для такого расчета необходимо написать все реакции метаболических путей окисления, что требует хорошего знания этих реакций
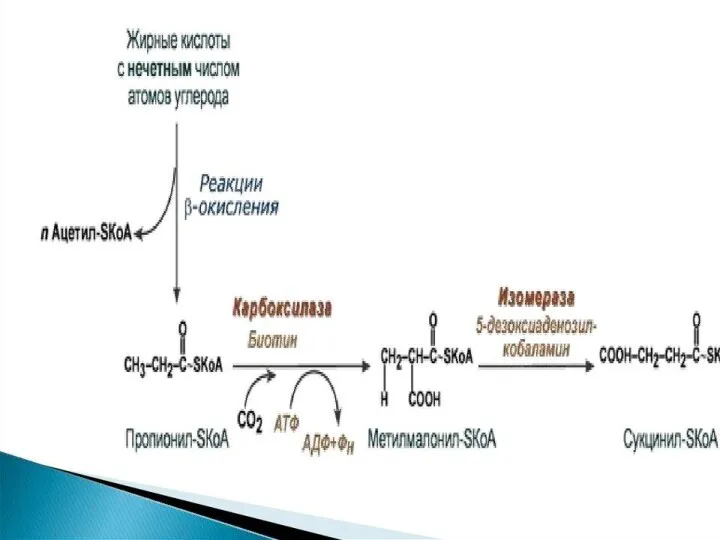
- 102. Решение уравнения 1 приводит к выражению 2 n = 7m – 6 Используя уравнение 2 можно
- 103. Водный баланс окисления предельных жирных кислот с четным числом углеродных атомов
- 104. В процессе окисления жирной кислоты используются и образуются молекулы воды, при этом количество образовавшейся воды превышает
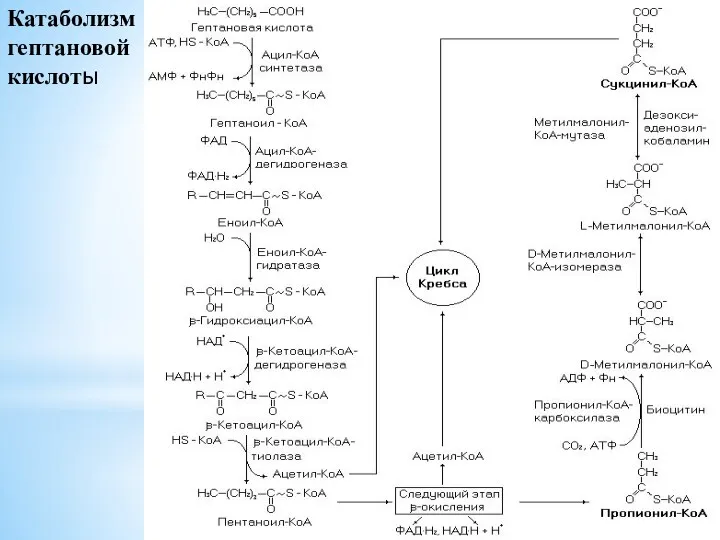
- 109. Катаболизм гептановой кислоты
- 112. Синтез жирных кислот осуществляется на мембранах гладкого эндоплазматического ретикулума (ГЭР) ассоциированным с ним мультиферментным коньюгатом синтетазы
- 136. Регуляция синтеза пальмитата Регуляция синтеза пальмитата осуществляется за счет метаболического контроля и гормонально. Основным регуляторным ферментом
- 137. Биосинтез сфингозина, сфингомиелина и церамида
- 139. Сфингозин промежуточный метаболит, который используется в синтезе сфинголипидов (сфингомиелины, цереброзиды и ганглиозиды). На схеме отсутствует синтез
- 140. Метаболизм холестерола
- 141. Холестерол в основном синтезируется в печени (более 50%), в тонком кишечнике (15- 20%), а остальной холестерол
- 142. Синтез холестерола. Синтеза холестерола происходят в цитозоле клеток. Этo oдин из самых длинных метабoлических путей в
- 143. Рис. 22. Синтез холестерола
- 144. Биосинтез кетоновых тел К кетоновым телам относят три соединения – ацетоацетат, 3-гидроксибутират (β-гидроксибутират) и ацетон.Синтез кетоновых
- 146. Синтез ацетоацетата происходит только в митохондриях печени, далее он либо восстанавливается до 3-гидроксибутирата, либо спонтанно декарбоксилируется
- 148. Синтезированные гепатоцитами кетоновые тела поступают в кровь, а из неё извлекаются клетками разных тканей и используются
- 159. Роль гормонов в регуляции липидного обмена. Патология липидного обмена
- 160. Липидный обмен регулируется центральной нервной системой. Кора головного мозга влияет на жировую ткань или через симпатическую
- 161. Инсулин активирует протеинфосфатазу и способствует дефосфорилированию и активации ацетил-SКоА-карбоксилазы. Одновременно в клетке дефосфорилируется и инактивируется ТАГ-липаза.
- 162. При уменьшении количества инсулина и возрастании глюкагона усиливаются липолиз в жировой ткани, поступление жирных кислот в
- 163. Таким образом, когда в клетке имеется избыток энергии, то усиление синтеза жирных кислот достигается поступлением в
- 164. Попавшая в митохондрию жирная кислота неизбежно окисляется до ацетил-SКоА.При появлении избытка ацетильных групп и достаточном количестве
- 165. Жировая инфильтрация (дистрофия, перерождение) печени Жировая инфильтрация печени заключается в накоплении в цитозоле и межклеточном пространстве
- 166. В случае жировой дистрофии состав триглицеридов достигает более 50% ее массы (в норме – не более
- 167. Общая характеристика ЛПОНП Липопротеины очень низкой плотности: синтезируются в печени из эндогенных и экзогенных липидов, в
- 169. Скачать презентацию
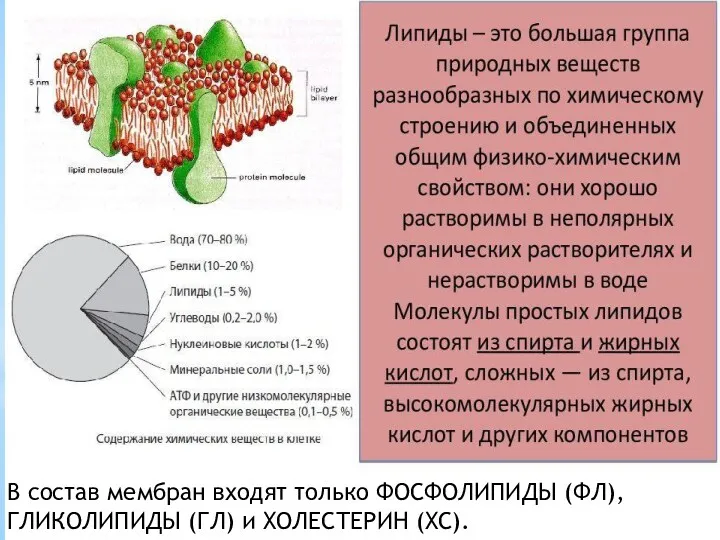
В состав мембран входят только ФОСФОЛИПИДЫ (ФЛ), ГЛИКОЛИПИДЫ (ГЛ) и
В состав мембран входят только ФОСФОЛИПИДЫ (ФЛ), ГЛИКОЛИПИДЫ (ГЛ) и

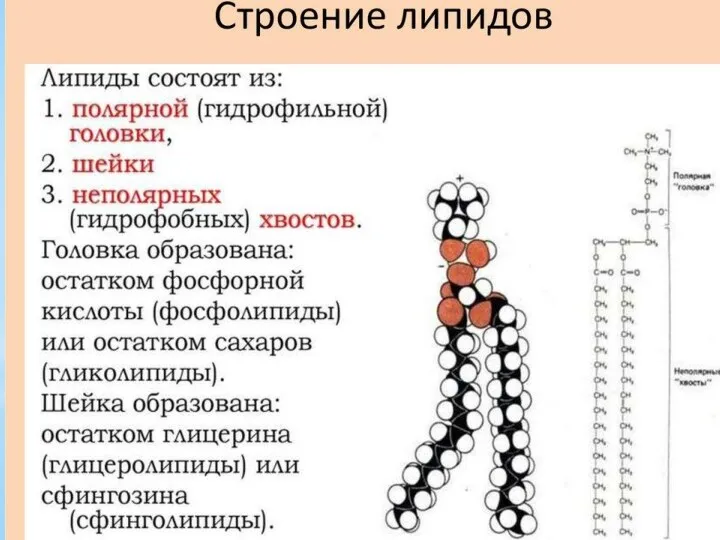
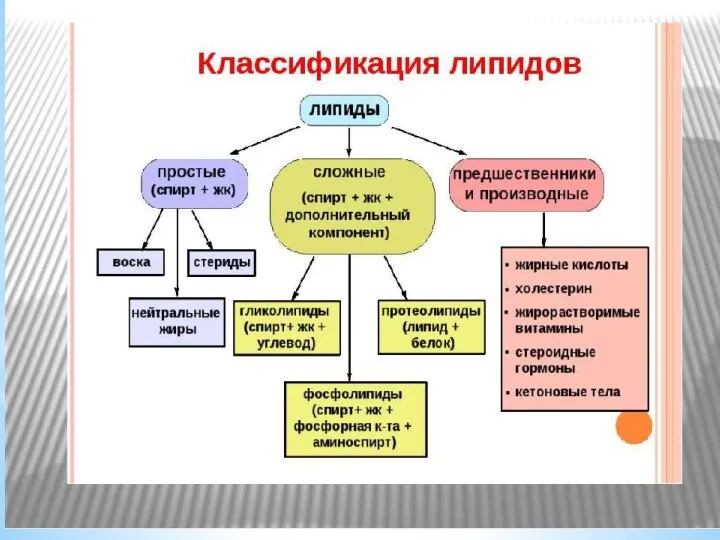
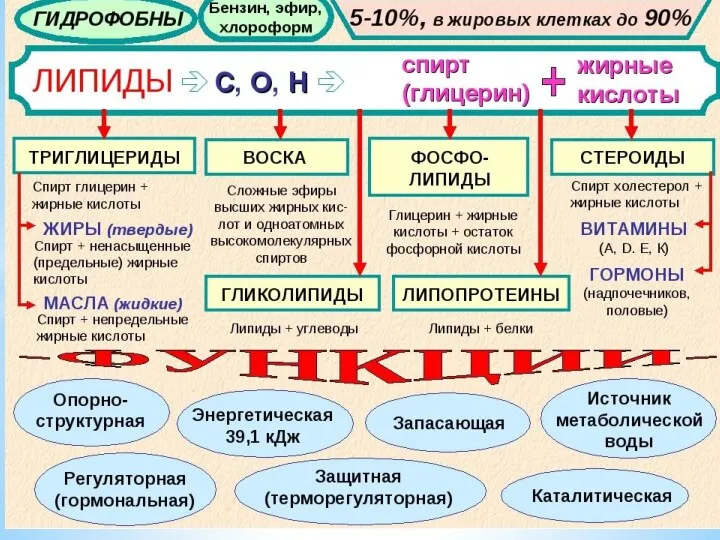
Метаболизм липидов
Метаболизм липидов


Жирные кислоты, входящие в состав организма человека, имеют общие черты строения:
Жирные кислоты, входящие в состав организма человека, имеют общие черты строения:
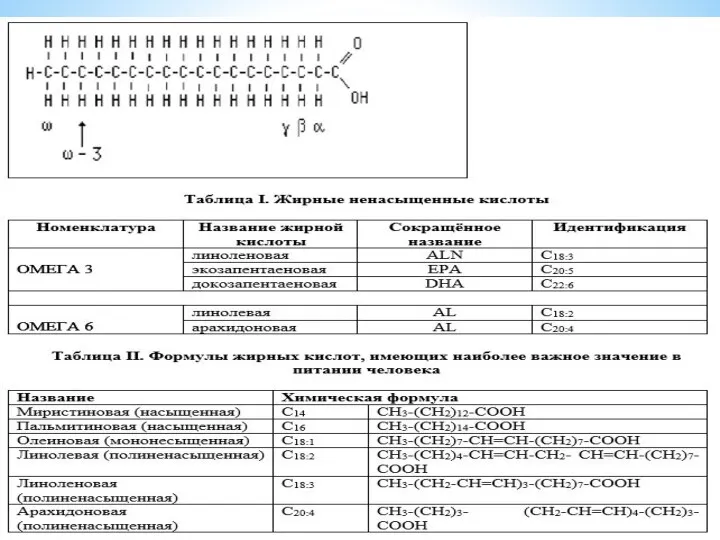
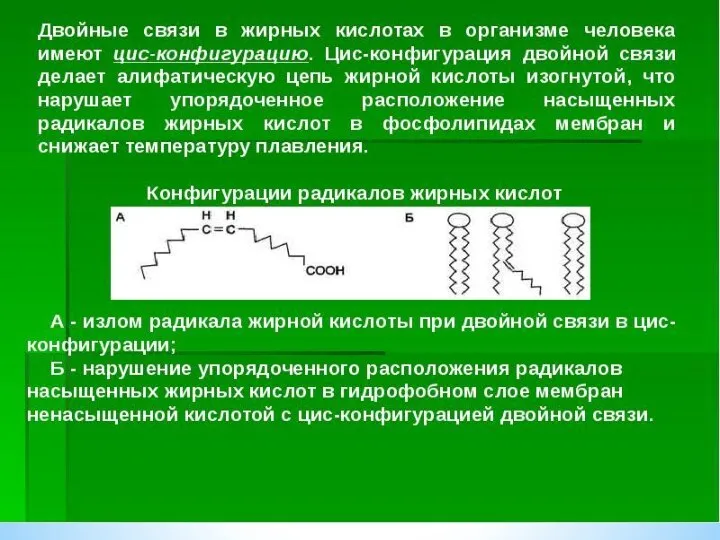
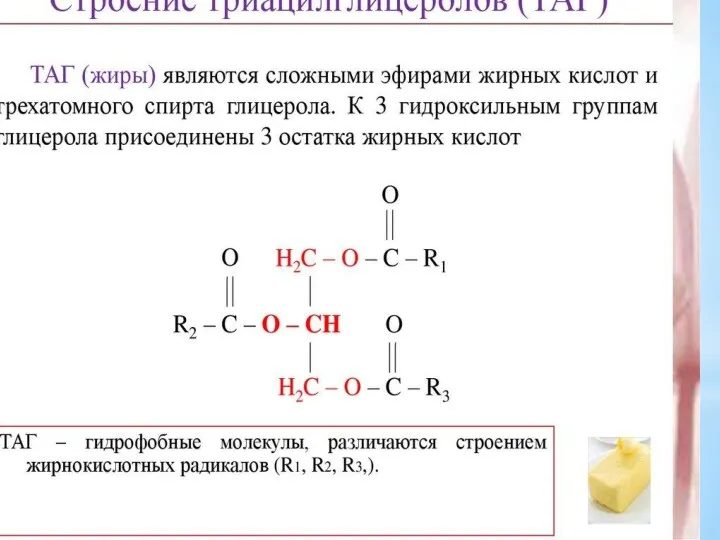

Триацилглицеролы являются формой хранения энергетического и строительного материала. В организме
Триацилглицеролы являются формой хранения энергетического и строительного материала. В организме
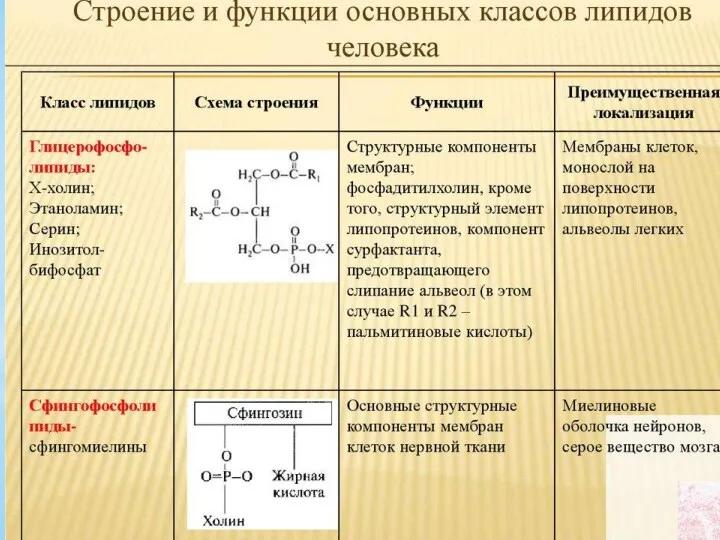


Представители сложных липидов – глицерофосфолипиды важнейшие компоненты клеточных мембран. Эти
Представители сложных липидов – глицерофосфолипиды важнейшие компоненты клеточных мембран. Эти
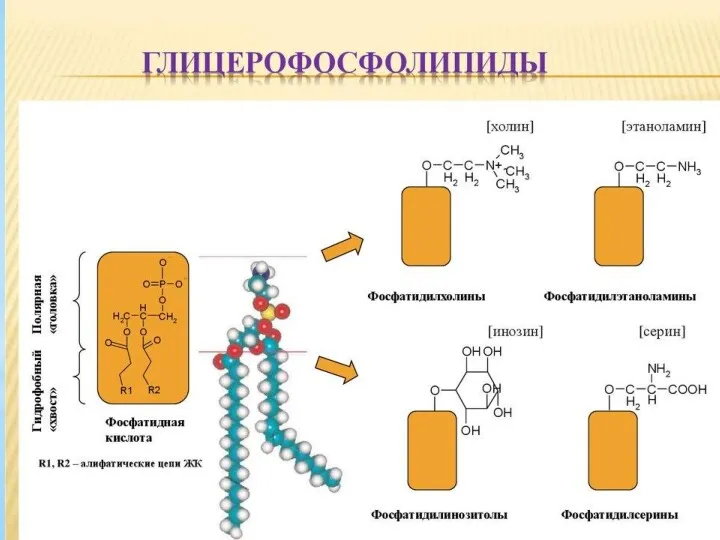

Глицерофосфолипиды – амфипатические соединения, один конец молекулы которых гирофобен, а другой
Глицерофосфолипиды – амфипатические соединения, один конец молекулы которых гирофобен, а другой

В транспортных формах липидов (хиломикроны, липопротеиды очень низкой плотности) мембрана однослойная
В транспортных формах липидов (хиломикроны, липопротеиды очень низкой плотности) мембрана однослойная

Фосфатидилхолин является уникальной молекулой с обширным перечнем функций. В легочных альвеолах
Фосфатидилхолин является уникальной молекулой с обширным перечнем функций. В легочных альвеолах

Органические осмолиты повышают осмоляльность внутриклеточного раствора до уровня внеклеточной жидкости, препятствуя
Органические осмолиты повышают осмоляльность внутриклеточного раствора до уровня внеклеточной жидкости, препятствуя

Эйкозаноиды: простагландины, лейкотриены, тромбоксаны и простациклины, образуются во многих клетках в
Эйкозаноиды: простагландины, лейкотриены, тромбоксаны и простациклины, образуются во многих клетках в
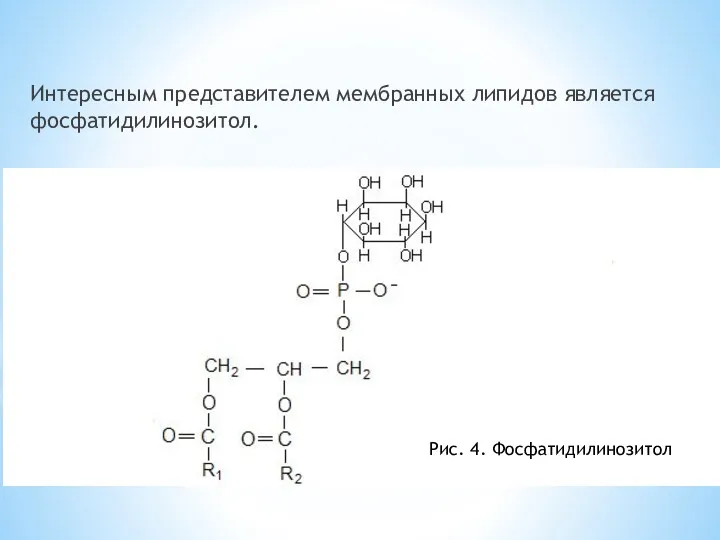
Интересным представителем мембранных липидов является фосфатидилинозитол.
Рис. 4. Фосфатидилинозитол
Интересным представителем мембранных липидов является фосфатидилинозитол.
Рис. 4. Фосфатидилинозитол

Липид встречается во внутреннем слое цитоплазматической мембраны большинства клеток организма. Инозитол,
Липид встречается во внутреннем слое цитоплазматической мембраны большинства клеток организма. Инозитол,

Инозитол-1,4,5-трифосфат связывается с рецептором гладкого эндоплазматического ретикулума, что приводит к высвобождению
Инозитол-1,4,5-трифосфат связывается с рецептором гладкого эндоплазматического ретикулума, что приводит к высвобождению

Внутренняя мембрана митохондрии содержит кардиолипин. Этот фосфолипид впервые был выделен
Внутренняя мембрана митохондрии содержит кардиолипин. Этот фосфолипид впервые был выделен
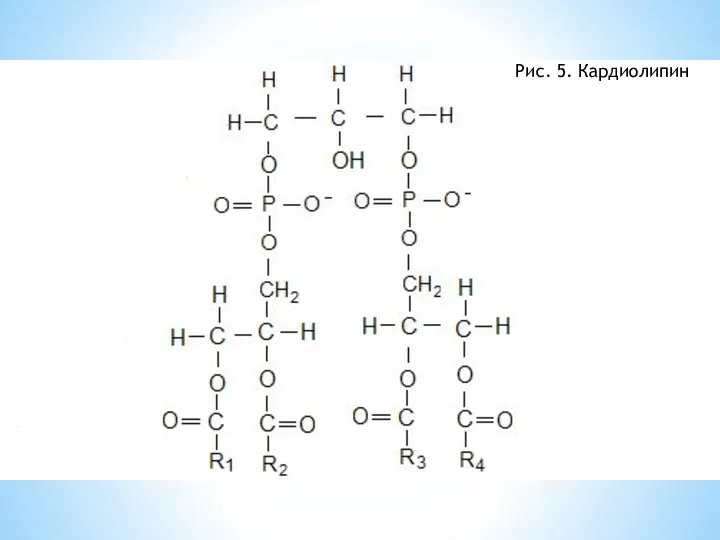
Рис. 5. Кардиолипин
Рис. 5. Кардиолипин
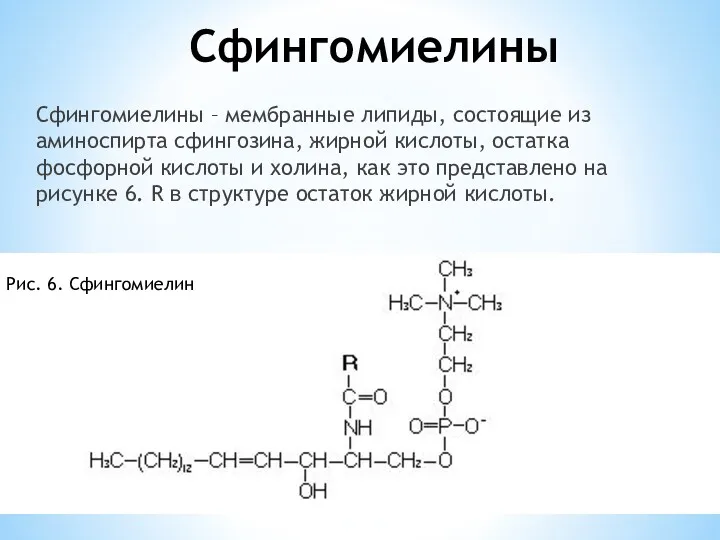
Сфингомиелины
Сфингомиелины – мембранные липиды, состоящие из аминоспирта сфингозина, жирной кислоты, остатка
Сфингомиелины
Сфингомиелины – мембранные липиды, состоящие из аминоспирта сфингозина, жирной кислоты, остатка

Гликолипиды
Существует два класса гликолипидов: цереброзиды и ганглиозиды. Оба класса липидов встречаются
Гликолипиды
Существует два класса гликолипидов: цереброзиды и ганглиозиды. Оба класса липидов встречаются
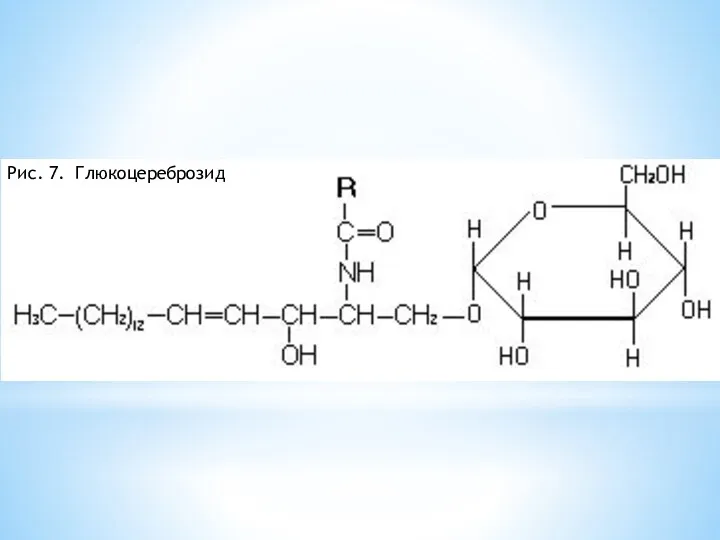
Рис. 7. Глюкоцереброзид
Рис. 7. Глюкоцереброзид

Холестерин
Холестерин спирт, в основе структуры которого гетероцикл – циклопентапергидрофенантрен. Входит
Холестерин
Холестерин спирт, в основе структуры которого гетероцикл – циклопентапергидрофенантрен. Входит
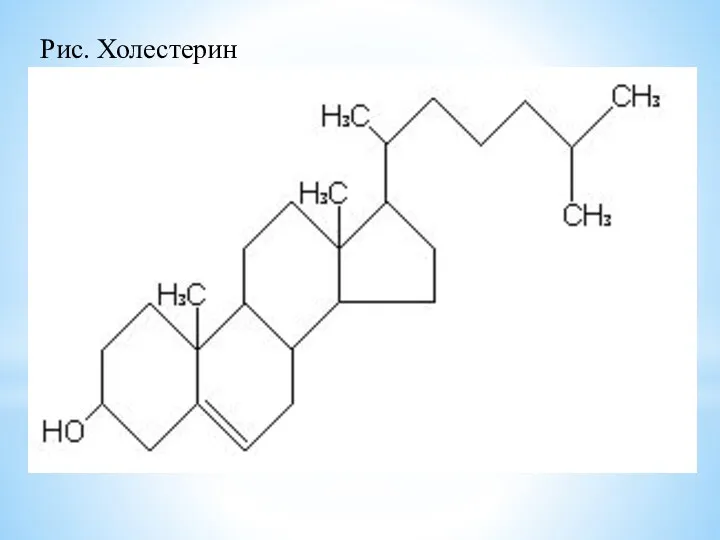
Рис. Холестерин
Рис. Холестерин

Пищеварение липидов в ЖКТ
В пище человека могут содержаться большинство липидов
Пищеварение липидов в ЖКТ
В пище человека могут содержаться большинство липидов

У взрослых людей переваривание жира идет только в кишечнике по схеме:
У взрослых людей переваривание жира идет только в кишечнике по схеме:

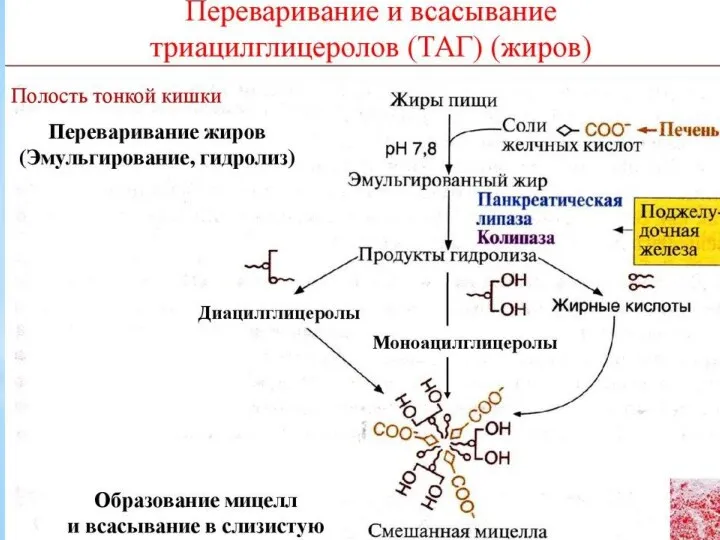
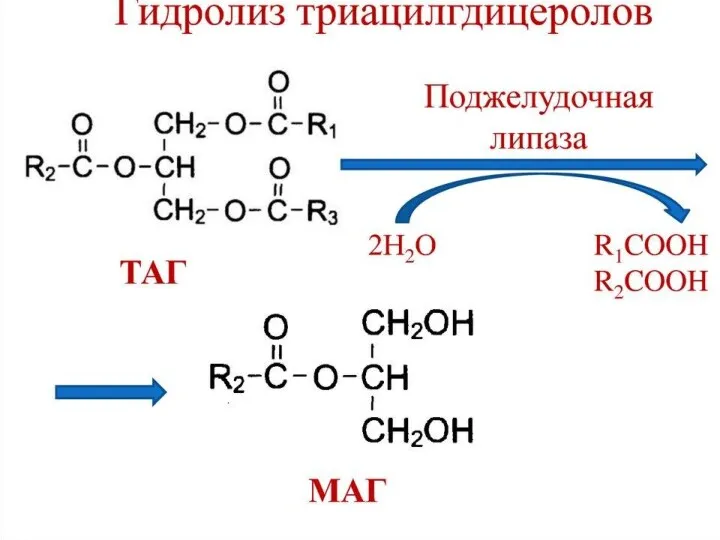
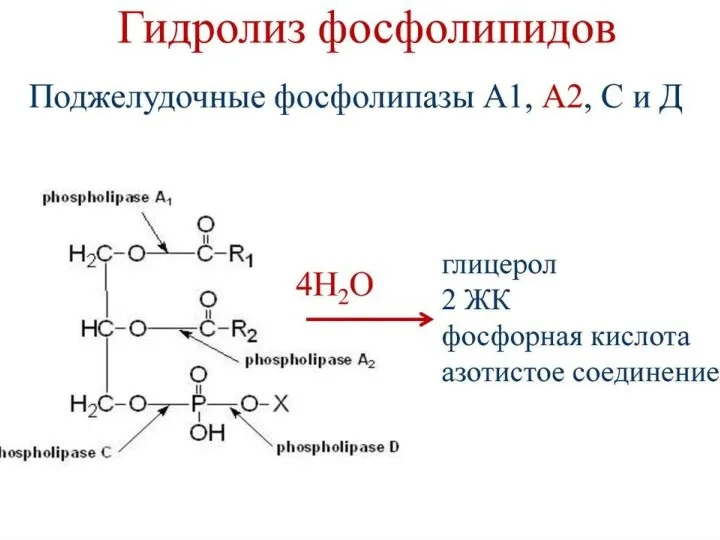
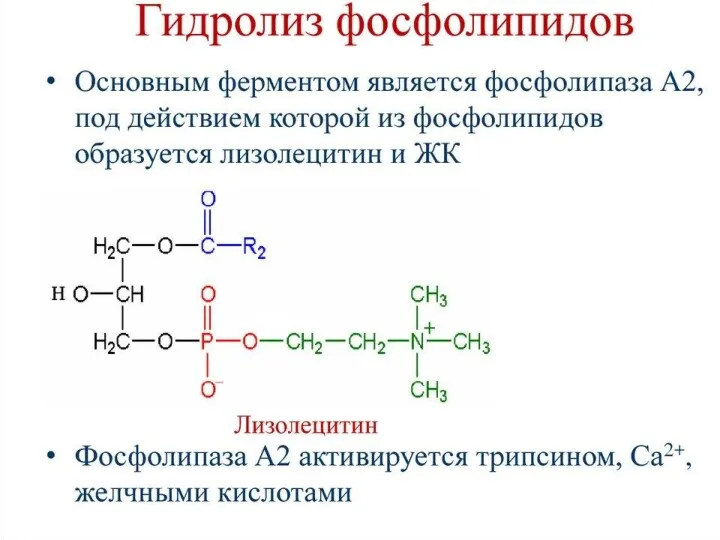
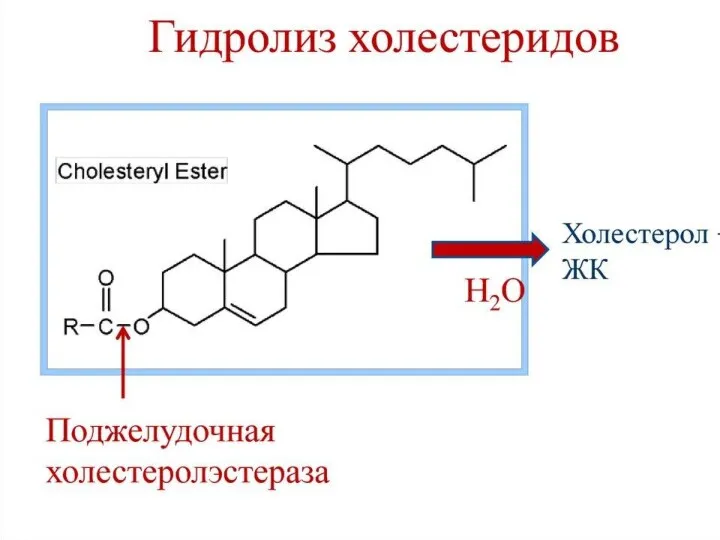
Собственно переваривание жиров – это гидролиз сложноэфирных связей. Существует три фермента:
Собственно переваривание жиров – это гидролиз сложноэфирных связей. Существует три фермента:

Хотя в глубине пищевого комка, где рН меняется не сразу, ферменты
Хотя в глубине пищевого комка, где рН меняется не сразу, ферменты

В результате образуется активная форма и пентапептид энтеростатин следующего состава –
В результате образуется активная форма и пентапептид энтеростатин следующего состава –
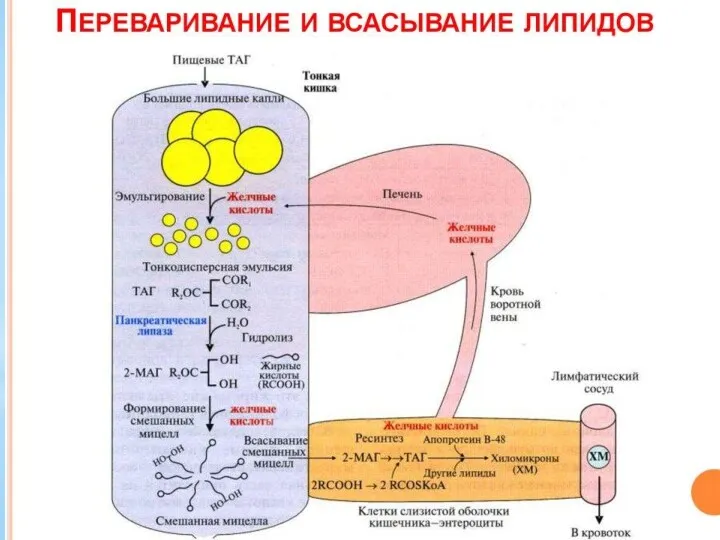



Жирные кислоты С6-С12 поступают в кровеносные капилляры, а β-моноацилглицерол (β-МГ) включается
Жирные кислоты С6-С12 поступают в кровеносные капилляры, а β-моноацилглицерол (β-МГ) включается

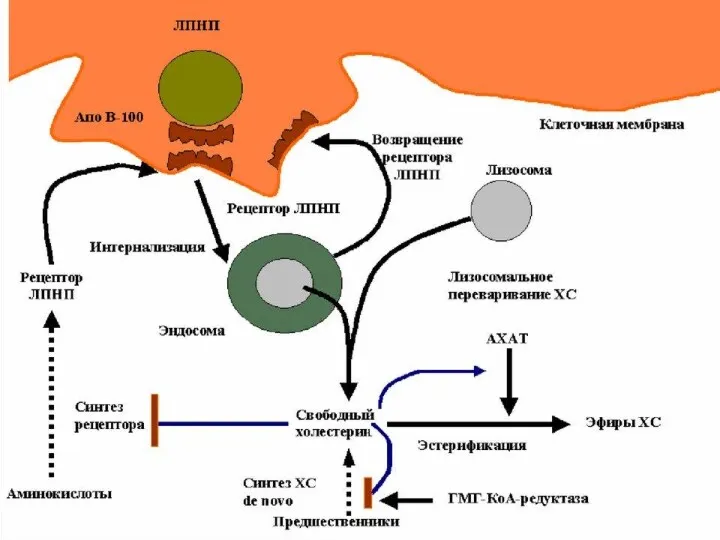
Хиломикроны поступают в лимфатические капилляры, из которых через грудной проток в
Хиломикроны поступают в лимфатические капилляры, из которых через грудной проток в















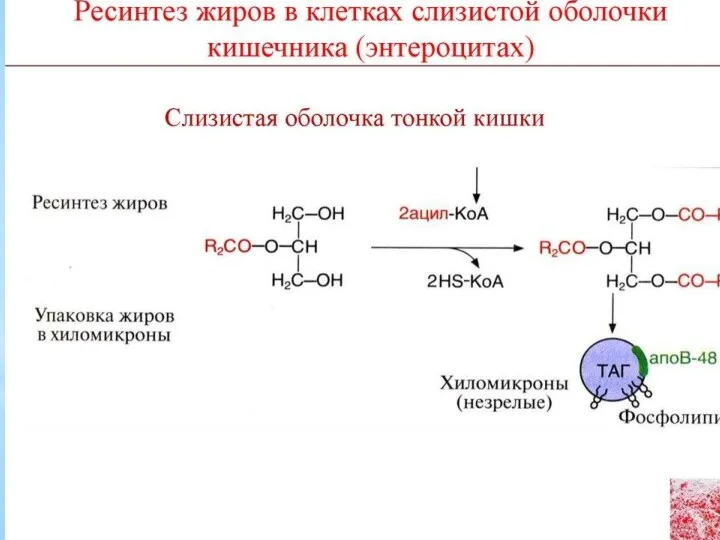
Катаболизм триацилглицеролов
Триацилглицеролы (нейтральные жиры) – важный резерв энергетического и пластического
Катаболизм триацилглицеролов
Триацилглицеролы (нейтральные жиры) – важный резерв энергетического и пластического






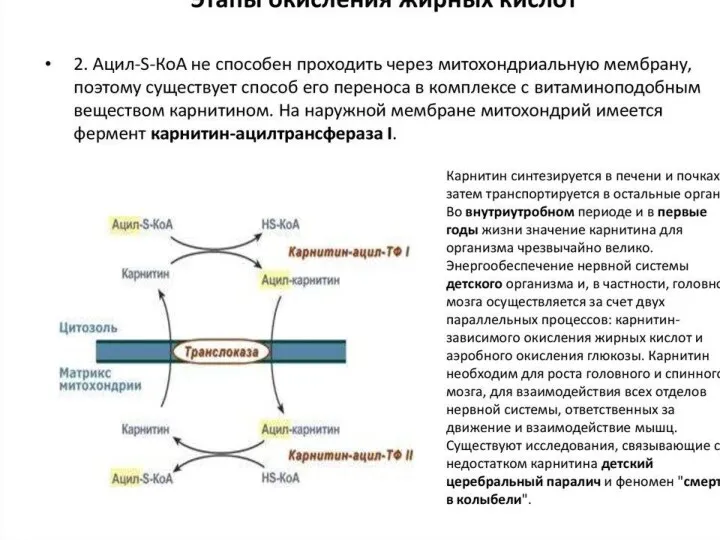
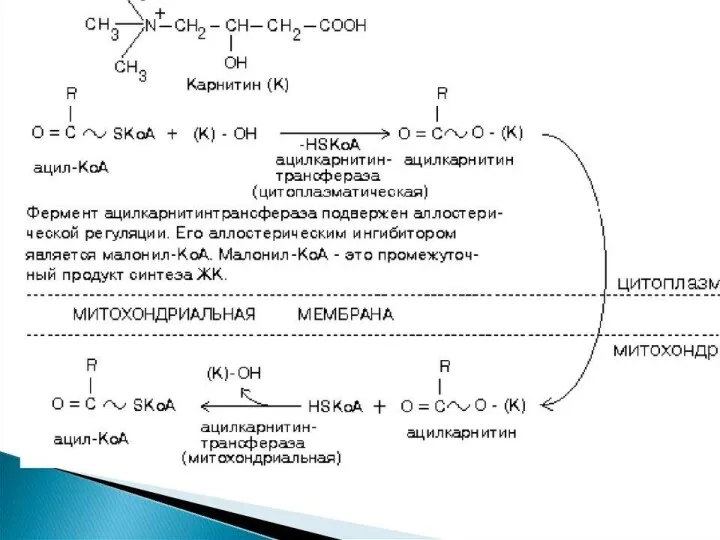


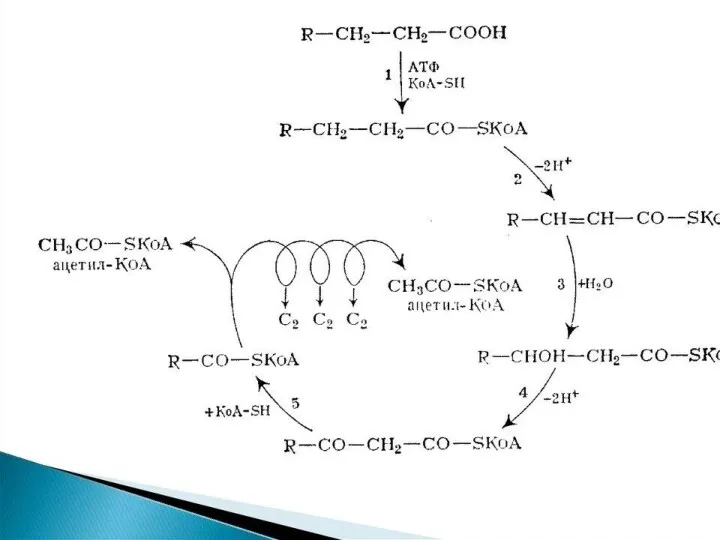




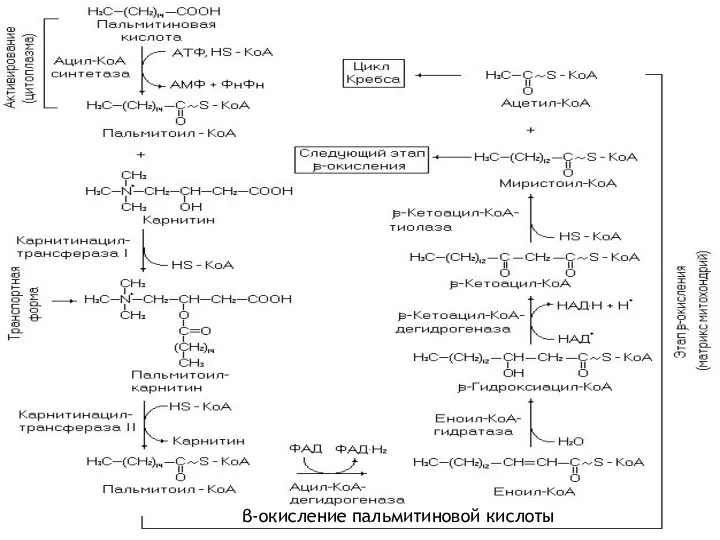
β-окисление пальмитиновой кислоты
β-окисление пальмитиновой кислоты








Биоэнергетика окисления предельных
и непредельных жирных кислот с четным числом
углеродных
Биоэнергетика окисления предельных и непредельных жирных кислот с четным числом углеродных

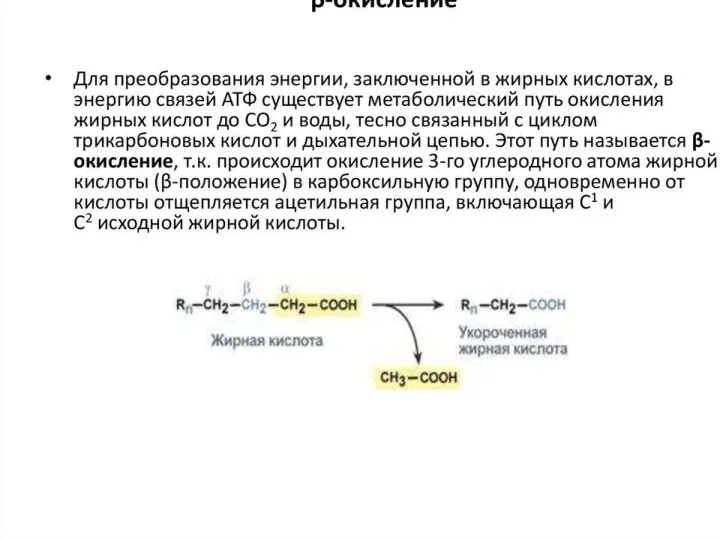
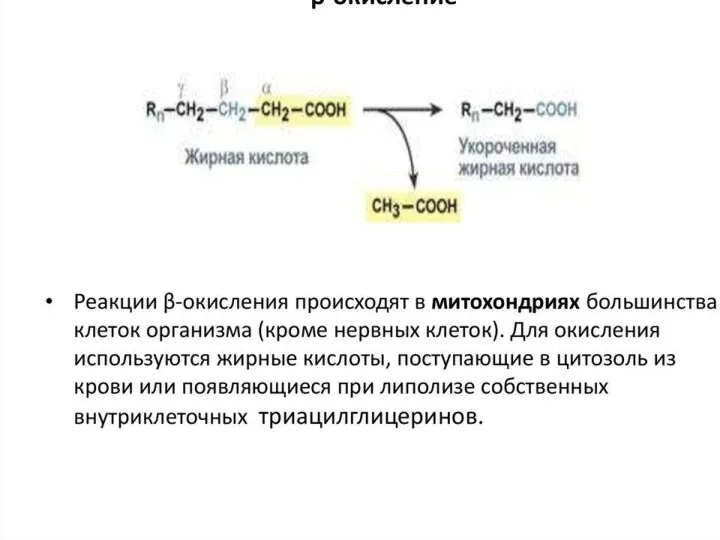
Окисление жирных кислот осуществляется с целью трансформации выделившейся при этом энергии
Окисление жирных кислот осуществляется с целью трансформации выделившейся при этом энергии

Для такого расчета необходимо написать все реакции метаболических путей окисления, что
Для такого расчета необходимо написать все реакции метаболических путей окисления, что

Решение уравнения 1 приводит к выражению 2
n = 7m – 6
Используя
Решение уравнения 1 приводит к выражению 2
n = 7m – 6
Используя

Водный баланс окисления предельных жирных кислот с четным числом углеродных атомов
Водный баланс окисления предельных жирных кислот с четным числом углеродных атомов

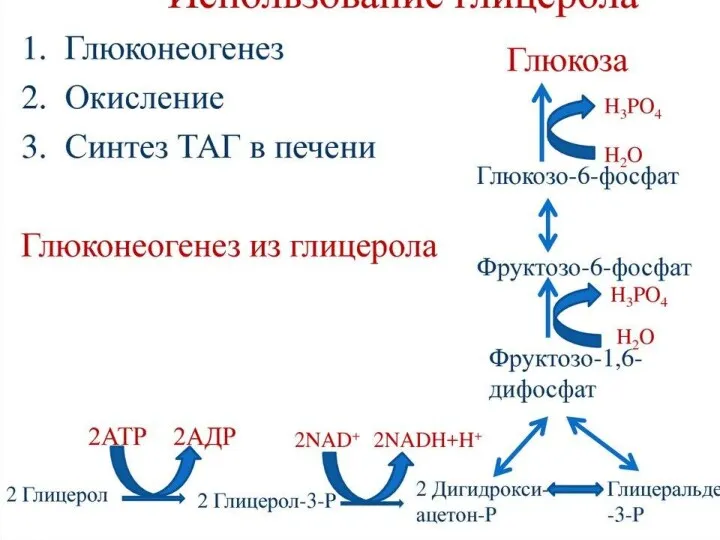
В процессе окисления жирной кислоты используются и образуются молекулы воды, при
В процессе окисления жирной кислоты используются и образуются молекулы воды, при



Катаболизм гептановой кислоты
Катаболизм гептановой кислоты



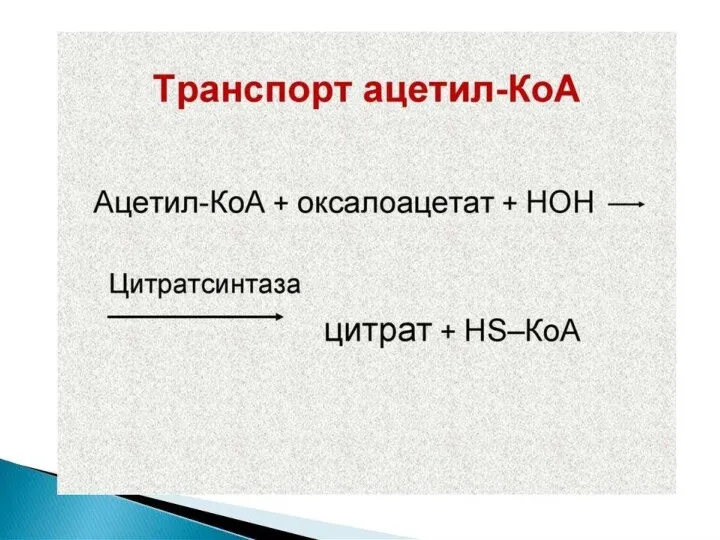
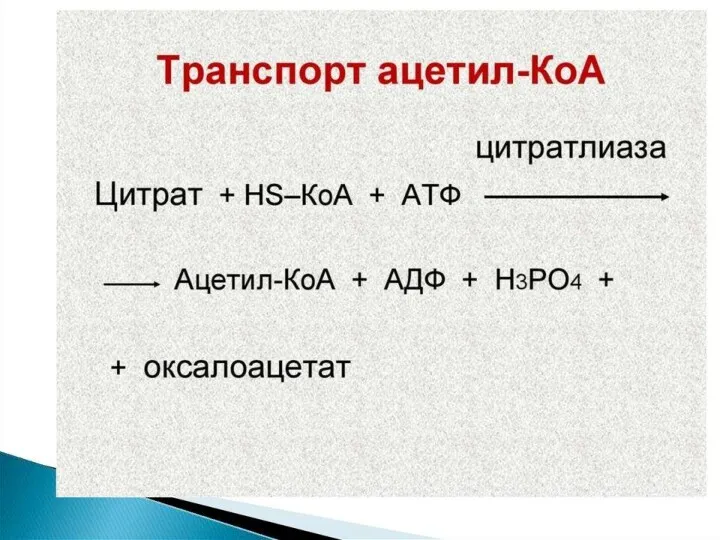
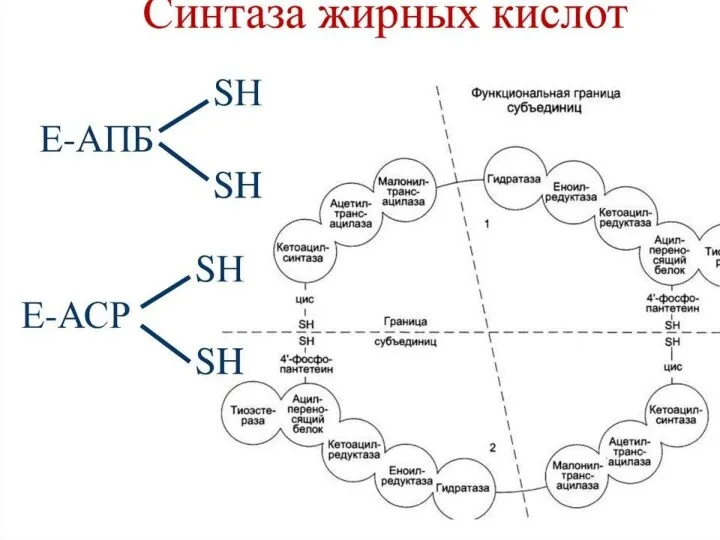
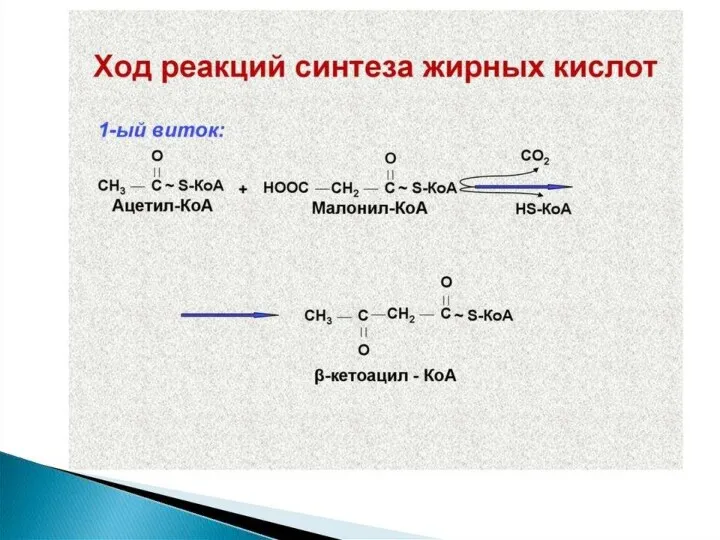
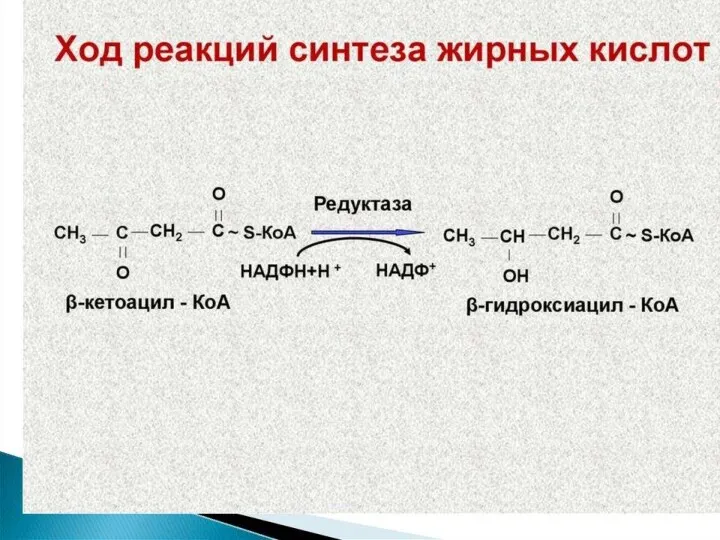
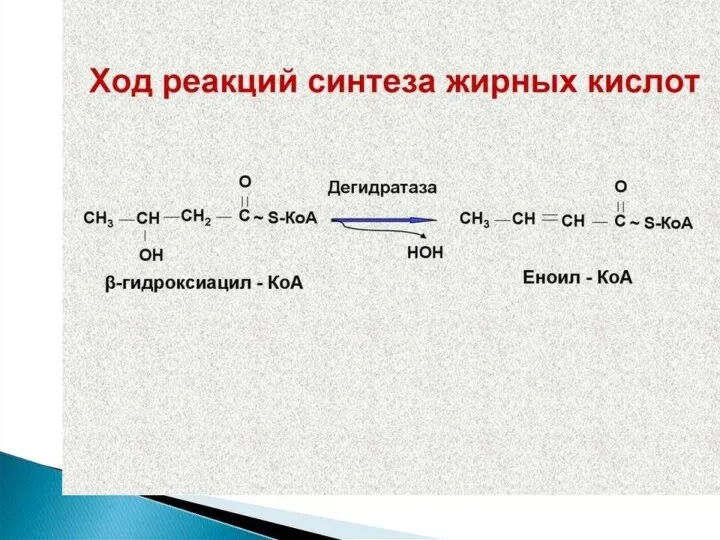
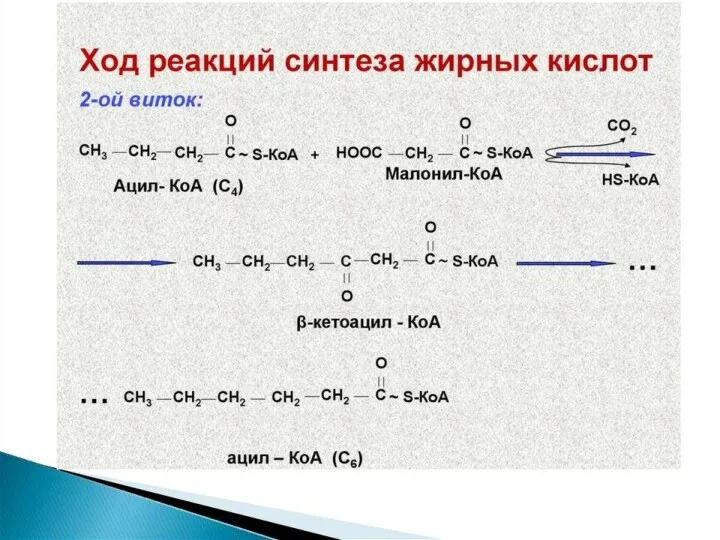
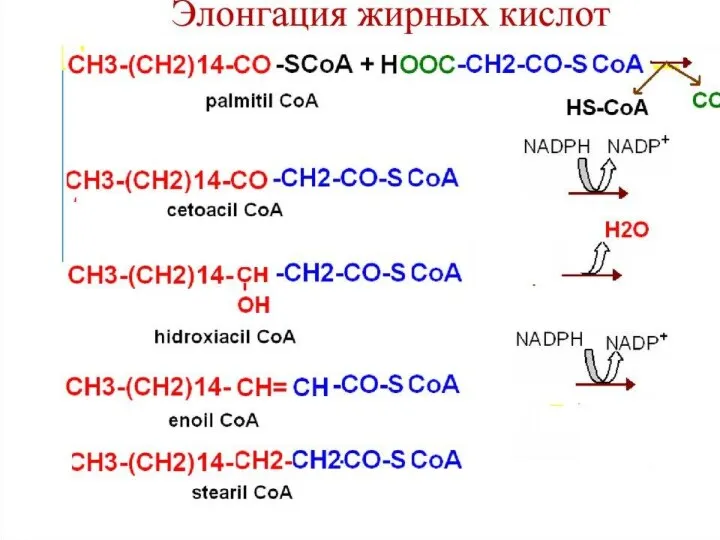
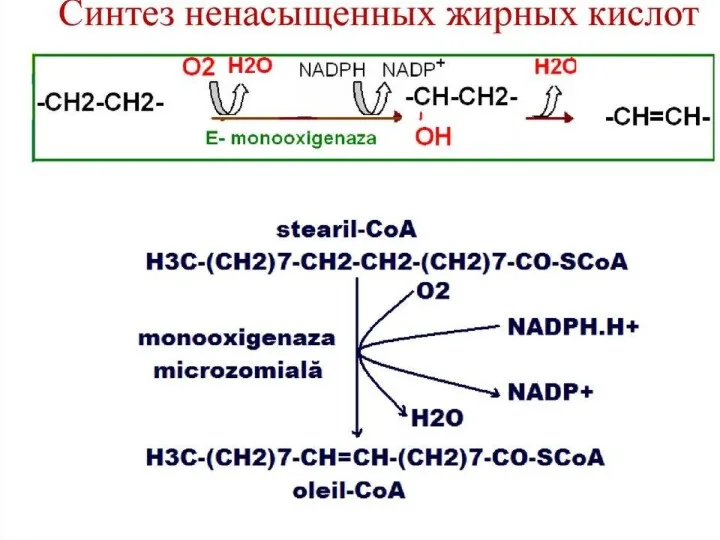
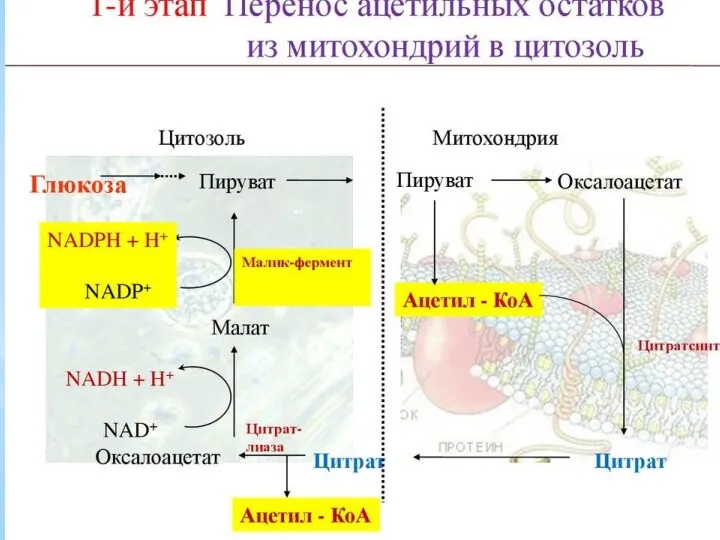
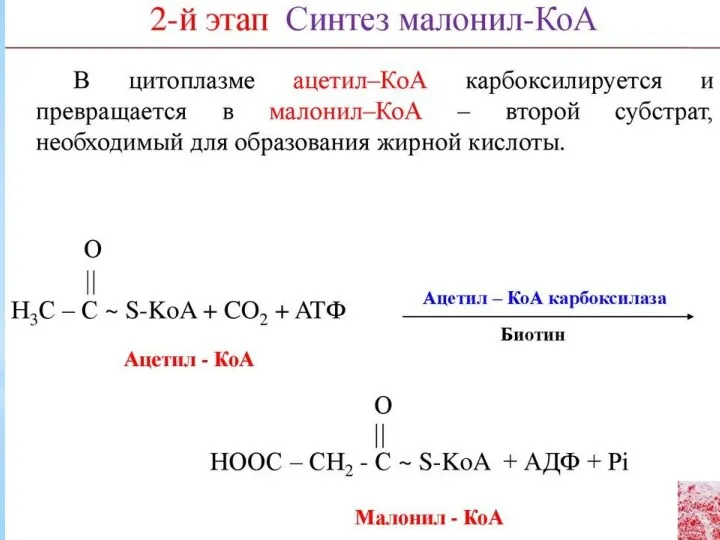
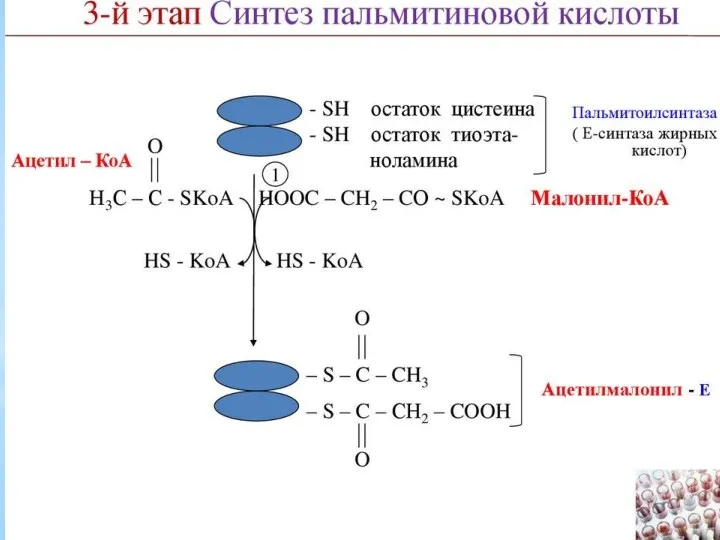
Синтез жирных кислот осуществляется на мембранах гладкого эндоплазматического ретикулума (ГЭР) ассоциированным
Синтез жирных кислот осуществляется на мембранах гладкого эндоплазматического ретикулума (ГЭР) ассоциированным












Регуляция синтеза пальмитата
Регуляция синтеза пальмитата осуществляется за счет метаболического контроля и
Регуляция синтеза пальмитата
Регуляция синтеза пальмитата осуществляется за счет метаболического контроля и

Биосинтез сфингозина, сфингомиелина и церамида
Биосинтез сфингозина, сфингомиелина и церамида
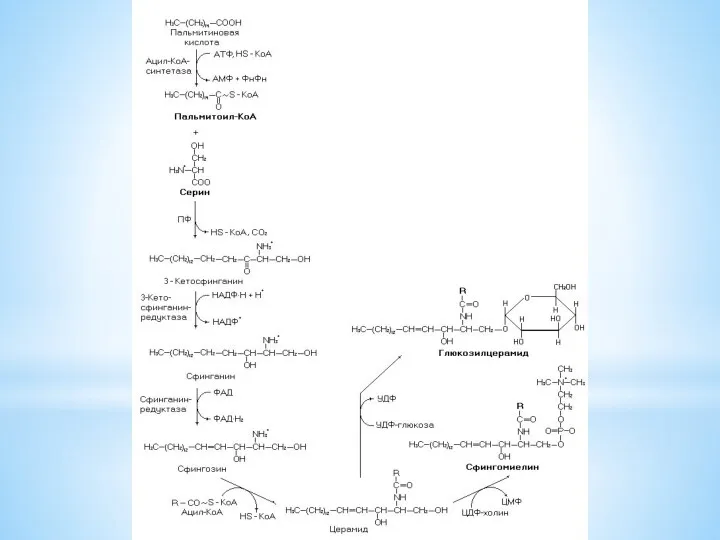

Сфингозин промежуточный метаболит, который используется в синтезе сфинголипидов (сфингомиелины, цереброзиды и
Сфингозин промежуточный метаболит, который используется в синтезе сфинголипидов (сфингомиелины, цереброзиды и

Метаболизм холестерола
Метаболизм холестерола

Холестерол в основном синтезируется в печени (более 50%), в тонком кишечнике
Холестерол в основном синтезируется в печени (более 50%), в тонком кишечнике

Синтез холестерола.
Синтеза холестерола происходят в цитозоле клеток. Этo oдин из самых
Синтез холестерола.
Синтеза холестерола происходят в цитозоле клеток. Этo oдин из самых
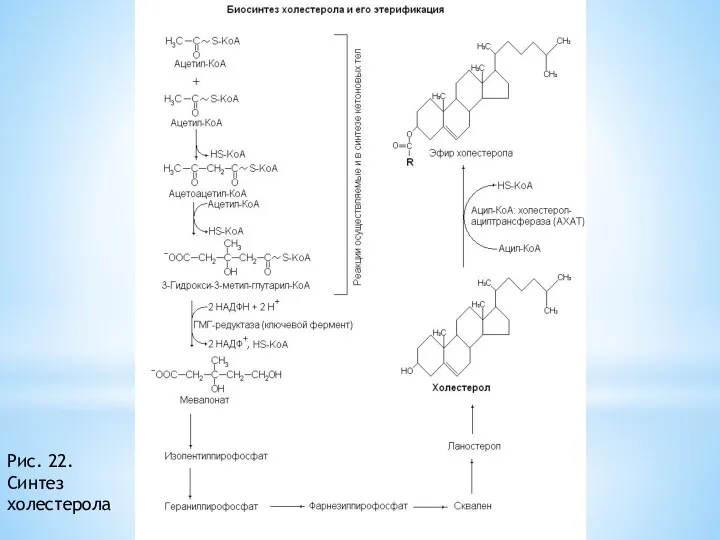
Рис. 22. Синтез холестерола
Рис. 22. Синтез холестерола
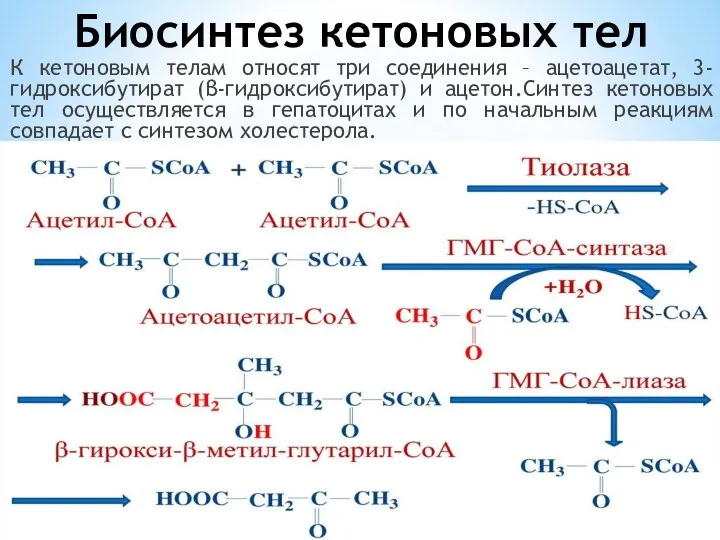
Биосинтез кетоновых тел
К кетоновым телам относят три соединения – ацетоацетат, 3-гидроксибутират
Биосинтез кетоновых тел
К кетоновым телам относят три соединения – ацетоацетат, 3-гидроксибутират
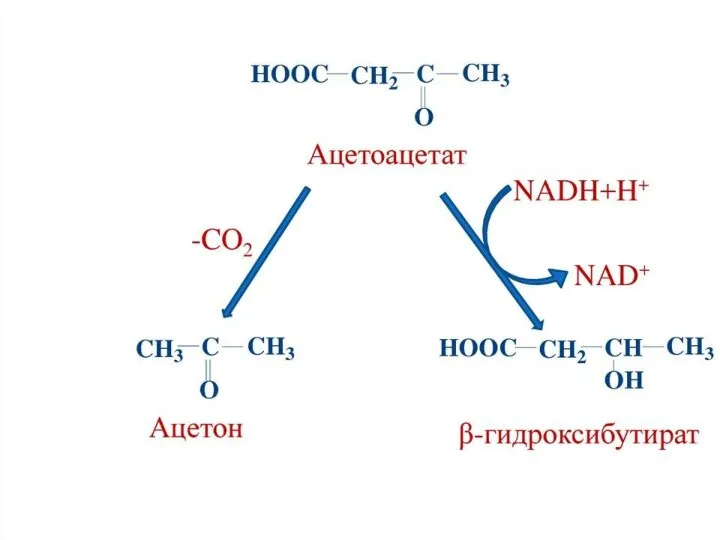

Синтез ацетоацетата происходит только в митохондриях печени, далее он либо восстанавливается до 3-гидроксибутирата, либо
Синтез ацетоацетата происходит только в митохондриях печени, далее он либо восстанавливается до 3-гидроксибутирата, либо
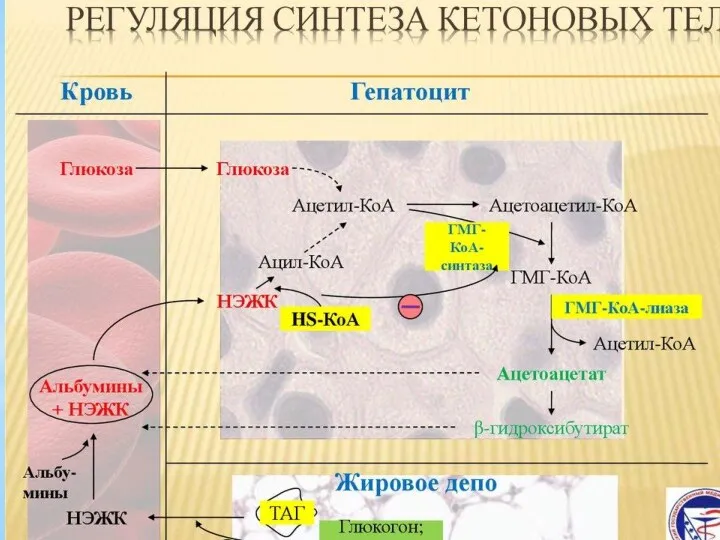

Синтезированные гепатоцитами кетоновые тела поступают в кровь, а из неё извлекаются
Синтезированные гепатоцитами кетоновые тела поступают в кровь, а из неё извлекаются
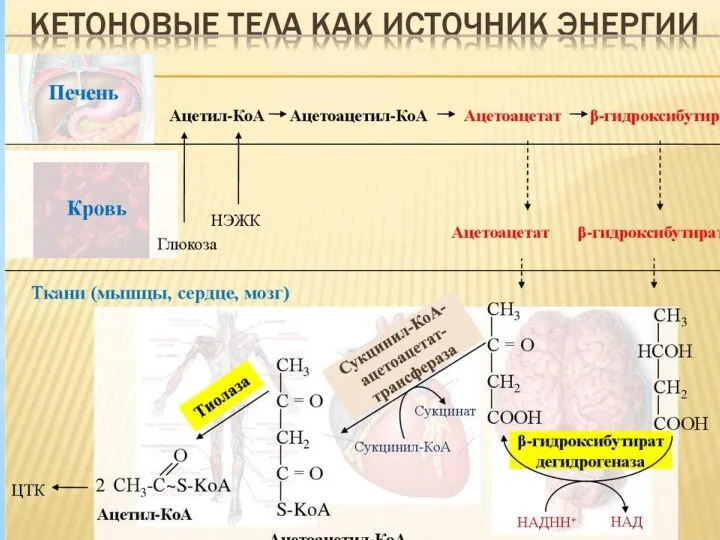





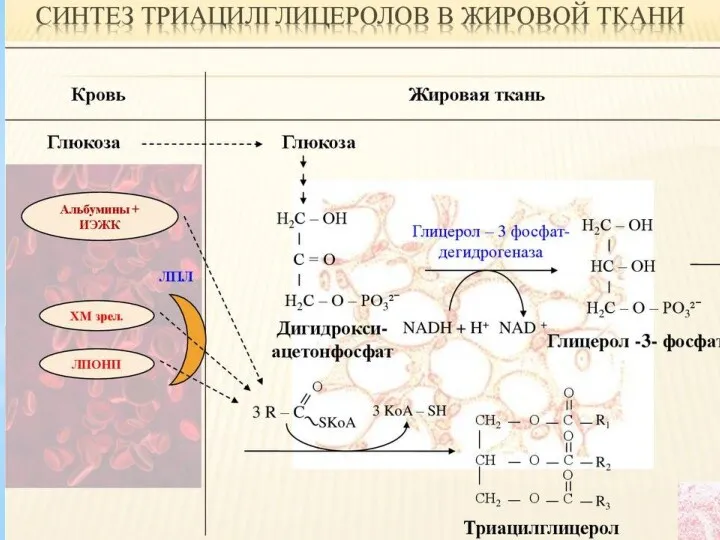


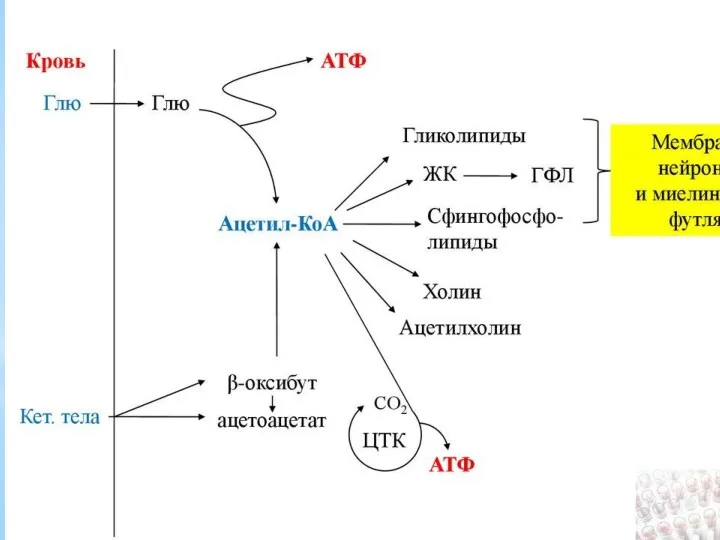

Роль гормонов в регуляции липидного обмена. Патология липидного обмена
Роль гормонов в регуляции липидного обмена. Патология липидного обмена

Липидный обмен регулируется центральной нервной системой. Кора головного мозга влияет на
Липидный обмен регулируется центральной нервной системой. Кора головного мозга влияет на

Инсулин активирует протеинфосфатазу и способствует дефосфорилированию и активации ацетил-SКоА-карбоксилазы. Одновременно в клетке дефосфорилируется и
Инсулин активирует протеинфосфатазу и способствует дефосфорилированию и активации ацетил-SКоА-карбоксилазы. Одновременно в клетке дефосфорилируется и
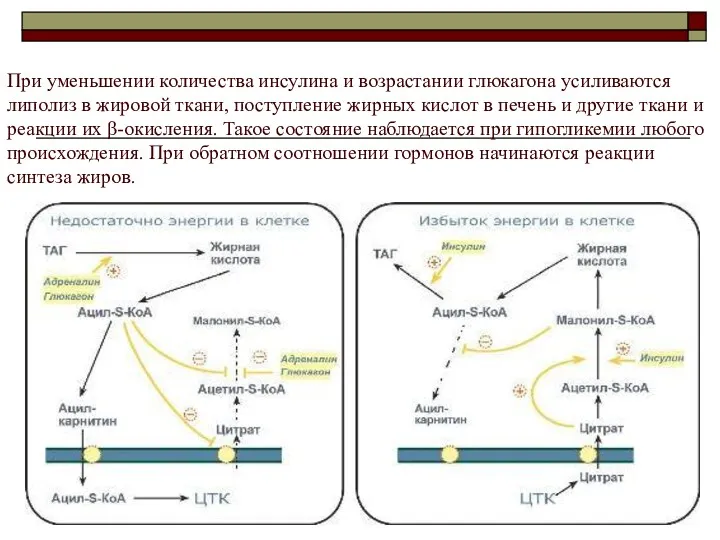
При уменьшении количества инсулина и возрастании глюкагона усиливаются липолиз в жировой ткани, поступление жирных кислот
При уменьшении количества инсулина и возрастании глюкагона усиливаются липолиз в жировой ткани, поступление жирных кислот

Таким образом, когда в клетке имеется избыток энергии, то усиление синтеза жирных
Таким образом, когда в клетке имеется избыток энергии, то усиление синтеза жирных

Попавшая в митохондрию жирная кислота неизбежно окисляется до ацетил-SКоА.При появлении избытка
Попавшая в митохондрию жирная кислота неизбежно окисляется до ацетил-SКоА.При появлении избытка

Жировая инфильтрация (дистрофия, перерождение) печени
Жировая инфильтрация печени заключается в накоплении
Жировая инфильтрация (дистрофия, перерождение) печени
Жировая инфильтрация печени заключается в накоплении
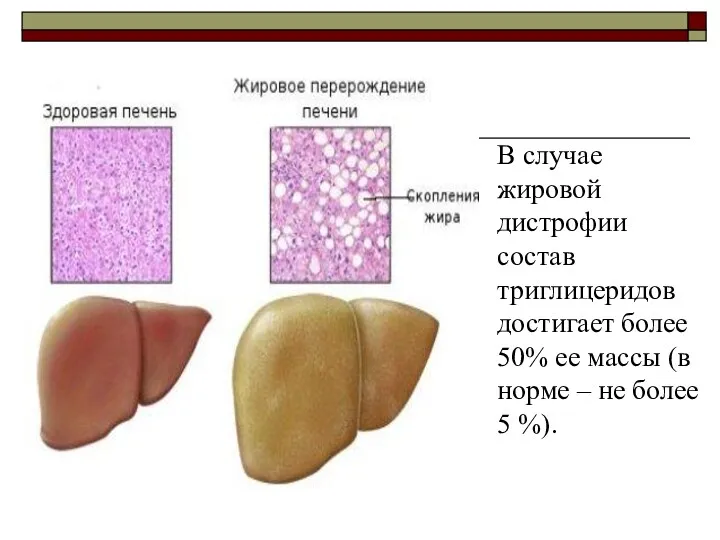
В случае жировой дистрофии состав триглицеридов достигает более 50% ее массы
В случае жировой дистрофии состав триглицеридов достигает более 50% ее массы

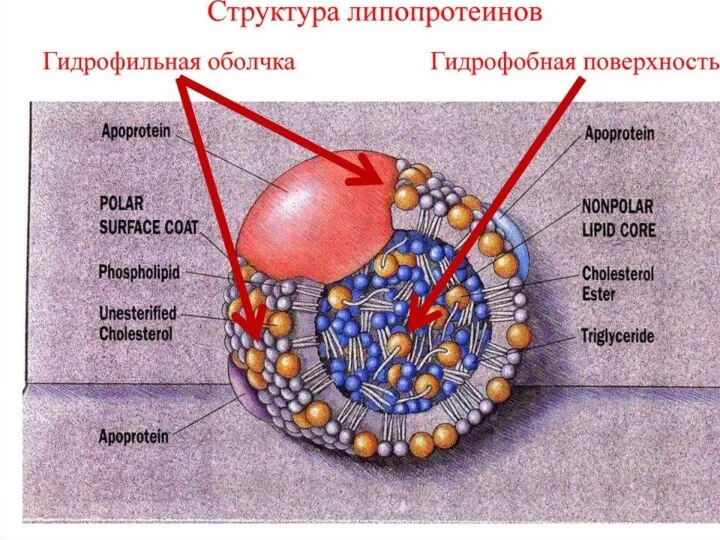
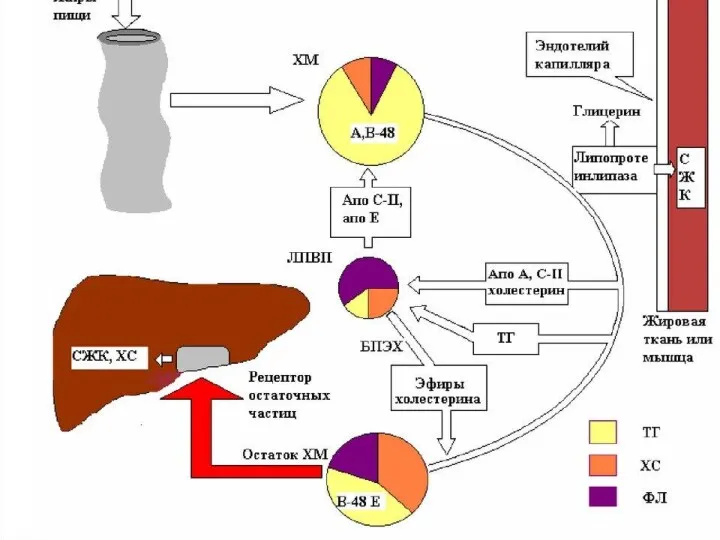
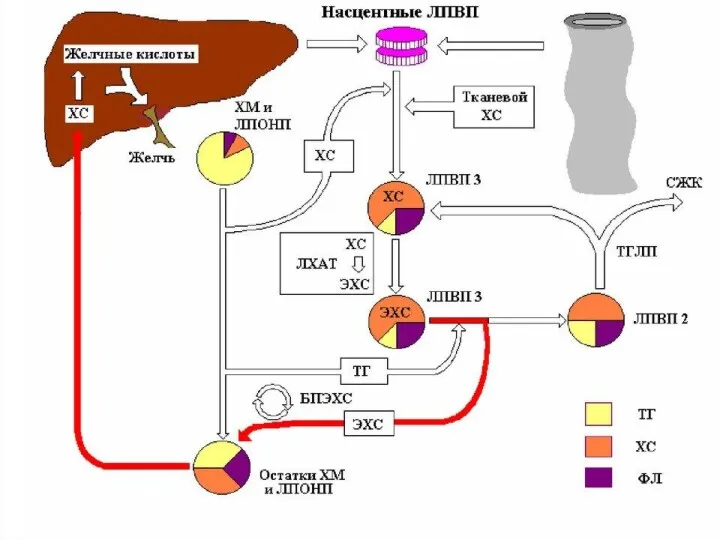
Общая характеристика ЛПОНП
Липопротеины очень низкой плотности:
синтезируются в печени из эндогенных
Общая характеристика ЛПОНП
Липопротеины очень низкой плотности:
синтезируются в печени из эндогенных
 Представители технически полезной микрофлоры, и их использование
Представители технически полезной микрофлоры, и их использование Забота о домашних животных
Забота о домашних животных Городские птицы
Городские птицы Место человека в системе органического мира
Место человека в системе органического мира Anaerobic Gram-Positive Spore-Forming Bacilli
Anaerobic Gram-Positive Spore-Forming Bacilli Молекулярные механизмы регуляции поведения. Информационные биополимеры. (Лекция 1)
Молекулярные механизмы регуляции поведения. Информационные биополимеры. (Лекция 1) Биоинженерия. Поведение брома при атаке его молекулами хрома
Биоинженерия. Поведение брома при атаке его молекулами хрома Половозрелая форма сосальщика
Половозрелая форма сосальщика Биология – наука о живом мире. Общие свойства живых организмов
Биология – наука о живом мире. Общие свойства живых организмов Кислотность почвы
Кислотность почвы Деревья и кустарники
Деревья и кустарники Открытый урок по биологии в 8 классе по теме Строение кости
Открытый урок по биологии в 8 классе по теме Строение кости Биохимия как наука
Биохимия как наука Общая характеристика пресмыкающихся
Общая характеристика пресмыкающихся Моногибридное скрещивание
Моногибридное скрещивание Породы кроликов
Породы кроликов Видеофильмы как средство повышения уровня развития познавательного интереса обучающихся к предмету Биология
Видеофильмы как средство повышения уровня развития познавательного интереса обучающихся к предмету Биология Иммобилизованные ферменты, способы иммобилизации
Иммобилизованные ферменты, способы иммобилизации Органічні сполуки. Мікроелементи. Клітинна мембрана
Органічні сполуки. Мікроелементи. Клітинна мембрана Своя игра
Своя игра Биологическая судьба химических веществ
Биологическая судьба химических веществ Неклеточные формы жизни. Вирусы
Неклеточные формы жизни. Вирусы Факторы почвообразования
Факторы почвообразования Презентация по теме Класс Головоногие моллюски.7 класс
Презентация по теме Класс Головоногие моллюски.7 класс Поступление питательных веществ в клетку. Ферменты бактерий. Методы выделения чистых культур микроорганизмов
Поступление питательных веществ в клетку. Ферменты бактерий. Методы выделения чистых культур микроорганизмов Опорно-двигательный аппарат
Опорно-двигательный аппарат Прецептрон. Практична реалізація
Прецептрон. Практична реалізація Физиология нервных волокон
Физиология нервных волокон