- Метаболизм РНК

Содержание
- 2. Транскрипция Экспрессия заложенной в гене информации происходит через образование молекулы РНК, транскрибируемой с матрицы ДНК.
- 3. РНК выполняет свои функции в виде одноцепочечных молекул, которые складываются различными способами. гораздо более разнообразны (много
- 4. В1959 году Нобелевская премия присуждена С.Очоа (Severo Ochoa) и А.Корнбергу (A.Kornberg) за открытие механизма биологического синтеза
- 5. Транскрипция Транскрипция у эукариот происходит в ядре. Транскрипция не связана с фазами клеточного цикла. Ускоряется и
- 6. Итальянский нейробиолог Рита Леви-Монтальчини и американский биохимик Стенли Коэн за открытие факторов роста получили Нобелевскую премию
- 7. Транскрибируются отдельные гены или группы генов –транскриптон. Клетка ограничивает экспрессию генетической информации. Вся совокупность молекул РНК,
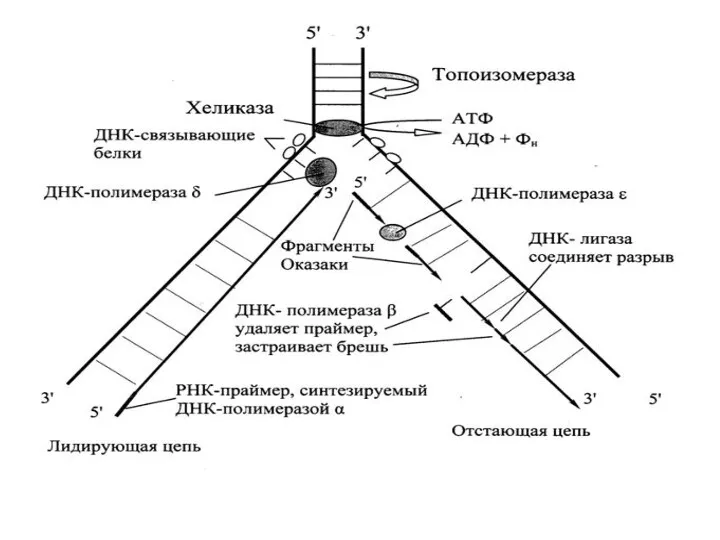
- 8. ДНК-зависимый синтез РНК. Транскрипция похожа на репликацию своим механизмом, направлением синтеза (от 5'→ З' концу), наличием
- 9. Транскрипция Транскрипция у E.coli осуществляется специальным ферментом ДНК-зависимой РНК–полимеразой. Рибонуклеозид-5'- трифосфаты -ЦТФ, ГТФ, АТФ, УТФ -
- 10. РНК-полимераза удлиняет цепь РНК, добавляя нуклеотиды к 3-гидроксильному концу (нуклеофил, атакует α-фосфат поступающего нуклеотида) (НМФ)n +
- 11. ДНК-зависимая РНК–полимераза E.coli - олигомерный фермент, состоящий из 5-ти субъединиц – α2ββ'ω –кор-фермент (Mr =390 000)
- 12. Все 6 субъединиц (α2ββ'ω + σ- субъединица) РНК-полимеразы называются холоферментом.
- 13. С помощью различных σ- субъединиц клетка может координировать экспрессию под физиологические потребности. Какой именно набор генов
- 15. РНК-полимеразы не имеют независимой З'→ 5' -экзонуклеазной активности для исправления ошибок, поэтому частота ошибок выше, 1
- 16. Транскрипция В процессе транскрипции различают стадии: инициации элонгации терминации
- 17. Процесс инициации транскрипции состоит из двух основных этапов: связывания РНК-полимеразы с ДНК. собственно инициация.
- 18. Первый этап: σ- субъединица временно связывается с кор-ферментом и в составе холофермента направляет фермент к специфическому
- 19. В составе бактериальных промоторов выделяют консервативные последовательности. Эти последовательности называют консенсусными последовательностями (ТАТААТ, ТТGАСА). Расположены на
- 20. Эффективность связывания РНК-полимеразы с промотором определяется консенсусными последовательностями, расстоянием между ними и их удаленностью от точки
- 21. В состав некоторых промоторов входит богатый АТ основаниями UP-элемент (upstream promoter). У E.coli α- субъединица РНК-полимеразы
- 22. Промотор не несет информации и служит для присоединения и ориентации РНК-полимеразы. Определяется рамка считывания информации с
- 23. Второй этап инициации: в результате холофермент РНК-полимераза связывается с ДНК (интактная) и образуется закрытый промоторный комплекс,
- 24. Закрытый промоторный комплекс: В результате раскручивания цепей ДНК разрываются водородные связи между парами нуклеотидов ДНК и
- 25. Эти короткие фрагменты РНК провоцируют разрыв контактов РНК-полимеразы с промотором. σ- субъединица отсоединяется и замещается белком
- 26. На стадии элонгации кор-фермент РНК-полимераза осуществляет поэтапное присоединение нуклеотидов, комплементарных матричной цепи ДНК. Комплекс РНК-полимераза с
- 27. Механизм присоединения нуклеотидов: связывание входящего нуклеотида. если нуклеотид комплементарен нуклеотиду матричной цепи ДНК, то активный центр
- 28. Транслокация: перемещение РНК-полимеразы на 1 нуклеотид вперед по матрице ДНК. Присоединение следующего нуклеотида. Элонгация цепей РНК
- 29. Рифампицин связывается с β-субъединицей бактериальной РНК-полимеразы, препятствуя высвобождению промотора при транскрипции.
- 30. Терминация E.coli имеет два типа сигналов терминации: зависимый от присутствия ρ-белкового фактора ρ-независимый
- 31. ρ-независимые терминаторы имеют два свойства: первое: содержат последовательность, транскрипт которой имеет комплементарные участки, образующие структуры в
- 32. Во-вторых: их матричная цепь содержит консервативную последовательность из трех остатков А, которые при транскрипции превращаются в
- 33. Взаимодействие новосинтезированного олиго U-участка РНК с РНК-полимеразой приводит к возникновению транскрипционных пауз. За это время успевает
- 34. Синтез ~80% транскриптов E.coli терминируется по ρ-независимому пути. ρ-зависимый путь: необходим ρ-фактор – гомогексамерный белок, обладающий
- 35. ρ-фактор связывается с РНК в специальных пиримидин-богатых сайтах RUT (Rho utilization) и движется в направлении З'
- 36. ρ-фактор настигает транскрипционный комплекс на сайте терминации, пирофосфат перестает выделяться, ρ-фактор гидролизует АТФ, после чего происходит
- 37. Транскрипция
- 38. В клетках эукариот обнаружено 3 вида РНК-полимераз (I, II, III). состав комплекса каждая полимераза выполняет особые
- 39. POL I – синтез прерибосомной РНК (пре-рРНК) POL II – синтез мРНК промоторы POL II обладают
- 40. POL III образует молекулы тРНК, 5S рРНК и другие небольшие специализированные молекулы РНК.
- 41. POL II играет ведущую роль в экспрессии эукариотических генов. состоит из 12 субъединиц (Mr =10 000-220
- 42. RBP1 – гомология с β' - субъединицой прокариотической РНК-полимеразы. RBP2 – гомология с β - субъединицой
- 43. Для формирования активного транскрипционного комплекса POL II нуждается в ряде белков – факторы транскрипции (TF II
- 44. инициация ТАТА-связывающий белок (ТВР) взамодействует с ТАТА-боксом. ТВР связывается с фактором транскрипции TFIIB, который связывается с
- 45. комплекс TFIIB- ТВР связывается с другим комплексом TFIIF- POL II. TFIIF помогает точной стыковке POL II
- 46. TFIIH обладает хеликазной активностью и начинает раскручивание ДНК вблизи точки начала транскрипции РНК, гидролизуя АТФ. Образуется
- 47. Фосфорилирование POL II приводит к конформационным изменениям и инциирует транскрипцию. По мере синтеза первых 60-70 нуклеотидов
- 48. На стадии элонгации TFIIF- POL II остаются в комплексе. Усиливается факторами элонгации, которые препятствуют остановке транскрипции.
- 49. В фазе терминации POL II освобождается, дефосфорилируется, и может участвовать в новом синтезе.
- 50. Бледная поганка синтезирует α-аманитин, который прерывает образование мРНК в клетках, блокируя POL II, а в высоких
- 51. Процессинг РНК Многие молекулы РНК бактерий и практически все молекулы РНК эукариот после синтеза подвергаются процессингу.
- 52. процессинг матричной РНК Модификация 5'-конца: Модификации пре-мРНК начинаются на стадии элонгации. Когда длина первичного транскрипта достигает
- 53. процессинг матричной РНК Модифицированный 5'-конец удлиняет время жизни мРНК, защищая её от действия 5'-экзонуклеаз в цитоплазме.
- 54. процессинг матричной РНК Модификация 3'-конца 3'-конец большинства транскриптов, также подвергается модификации, при которой специальным ферментом полиА-полимеразой
- 55. процессинг матричной РНК полиА- «хвост» эукариот защищает мРНК от ферментативного расщепления. Многие молекулы мРНК прокариот имеют
- 56. Первичные транскрипты эукариот строго комплементарны матрице (ДНК), содержат как экзоны, так и интроны. обычно содержат один
- 57. первичные транскрипты прокариот также могут содержать интроны (бактерии, археи).
- 58. в процессе сплайсинга («сплайсинг» от англ, to splice - сращивать) интроны удаляются из первичного транскрипта, а
- 59. процессинг матричной РНК сплайсинг происходит в ядре, в цитоплазму поступает уже "зрелая" мРНК некоторые реакции процессинга
- 60. аутосплайсинг Процесс "вырезания" некоторых интронов протекает при участии самих РНК. впервые это было обнаружено Томасом Чеком
- 61. Сплайсинг
- 62. Большинство интронов являются сплайсосомными интронами. Удаление происходит и катализируется внутри крупного белкового комплекса сплайсосомы.
- 63. Сплайсосома состоит из специализированных РНК-белковых комплексов – малых ядерных рибонуклеопротеинов (мяРНП). Каждый мяРНП содержит одну молекулу
- 64. В реакциях сплайсинга участвуют 5 мяРНК (ядро эукариот) и примерно 50 белков. Белки и РНК в
- 65. на 5' конце сплайсосомные интроны имеют GU-последовательность. на 3' конце – AG-последовательность. Эти последовательности указывают место
- 66. Сложность организмов не коррелирует с количеством генов, кодирующих белки. Из некоторых транскриптов эукариотических мРНК образуется только
- 67. Альтернативный сплайсинг В результате процессинга могут образовываться различные мРНК из первичного транскрипта. Первичный транскрипт содержит молекулярные
- 68. Альтернативные варианты сплайсинга РНК могут использоваться для одновременного синтеза двух разных гормонов: кальцитонин (обмен кальция) в
- 69. Многие гены у млекопитающих подвергаются альтернативному сплайсингу, что значительно увеличивает количество кодируемых генами белков.
- 71. Число идентифицированных классов специализированных РНК быстро растет: малая ядерная РНК, малая ядрышковая РНК, микро-РНК, малые интерферирующие
- 72. Регулируют функции мРНК путем ее расщепления или подавления ее трансляции. Обнаружены у многоклеточных эукариот, в том
- 73. Сплайсинг Нобелевская премия 1993 года была присуждена Р.Дж.Робертсу и Ф.А.Шарпу (Richard J.Roberts, Philip A.Sharp) за открытие
- 74. Альтернативный сплайсинг
- 75. Регуляция транскрипции Потребности в продуктах любых генов изменяются в зависимости от состояния клетки и стадии развития.
- 76. Транскрипция –первая стадия в сложном и энергозатратном процессе синтеза белка, поэтому регуляция концентрации белков у бактерий
- 77. регуляция может происходить на любой стадии транскрипции, но чаще на стадии связывания РНК-полимеразы и инициации транскрипции.
- 78. регулируемой скоростью синтеза и деградации РНК. постсинтетическими модификациями, переводящими σ- субъединицу из активной формы в неактивную
- 79. специализированным классом анти σ- белков, связывающихся с определенным типом σ- субъединиц и делающих их недоступными для
- 80. Решающим фактором, управляющим экспрессией генов является концентрация в клетке соответствующей мРНК, что определяется скоростью ее синтеза
- 81. Скорость расщепления мРНК разных эукариотических генов различаются- от нескольких секунд до нескольких суток. Среднее время полужизни
- 82. мРНК разрушается рибонуклеазами, присутствующими во всех клетках. Структура бактериальной мРНК с ρ-независимым терминатором обеспечивает защиту от
- 83. Спасибо за внимание
- 85. Транскрипция Транскрипция ДНК происходит отдельными участками, в которые входит один или несколько генов - транскриптон Каждый
- 86. Транскрипция
- 87. Транскрипция Транскриптон бактерий называется опероном
- 89. Скачать презентацию

Транскрипция
Экспрессия заложенной в гене информации происходит через образование молекулы РНК, транскрибируемой
Транскрипция
Экспрессия заложенной в гене информации происходит через образование молекулы РНК, транскрибируемой

РНК выполняет свои функции в виде одноцепочечных молекул, которые складываются различными
РНК выполняет свои функции в виде одноцепочечных молекул, которые складываются различными

В1959 году Нобелевская премия присуждена С.Очоа (Severo Ochoa) и А.Корнбергу (A.Kornberg)
В1959 году Нобелевская премия присуждена С.Очоа (Severo Ochoa) и А.Корнбергу (A.Kornberg)

Транскрипция
Транскрипция у эукариот происходит в ядре.
Транскрипция не связана с фазами
Транскрипция
Транскрипция у эукариот происходит в ядре.
Транскрипция не связана с фазами

Итальянский нейробиолог Рита Леви-Монтальчини и американский биохимик Стенли Коэн за открытие
Итальянский нейробиолог Рита Леви-Монтальчини и американский биохимик Стенли Коэн за открытие

Транскрибируются отдельные гены или группы генов –транскриптон.
Клетка ограничивает экспрессию генетической информации.
Вся
Транскрибируются отдельные гены или группы генов –транскриптон.
Клетка ограничивает экспрессию генетической информации.
Вся

ДНК-зависимый синтез РНК.
Транскрипция похожа на репликацию своим механизмом, направлением синтеза (от
ДНК-зависимый синтез РНК.
Транскрипция похожа на репликацию своим механизмом, направлением синтеза (от

Транскрипция
Транскрипция у E.coli осуществляется специальным ферментом ДНК-зависимой РНК–полимеразой.
Рибонуклеозид-5'- трифосфаты -ЦТФ, ГТФ,
Транскрипция
Транскрипция у E.coli осуществляется специальным ферментом ДНК-зависимой РНК–полимеразой.
Рибонуклеозид-5'- трифосфаты -ЦТФ, ГТФ,

РНК-полимераза удлиняет цепь РНК, добавляя нуклеотиды к 3-гидроксильному концу (нуклеофил, атакует
РНК-полимераза удлиняет цепь РНК, добавляя нуклеотиды к 3-гидроксильному концу (нуклеофил, атакует

ДНК-зависимая РНК–полимераза E.coli - олигомерный фермент, состоящий из 5-ти субъединиц –
ДНК-зависимая РНК–полимераза E.coli - олигомерный фермент, состоящий из 5-ти субъединиц –

Все 6 субъединиц (α2ββ'ω + σ- субъединица) РНК-полимеразы называются холоферментом.
Все 6 субъединиц (α2ββ'ω + σ- субъединица) РНК-полимеразы называются холоферментом.

С помощью различных σ- субъединиц клетка может координировать экспрессию под физиологические
С помощью различных σ- субъединиц клетка может координировать экспрессию под физиологические
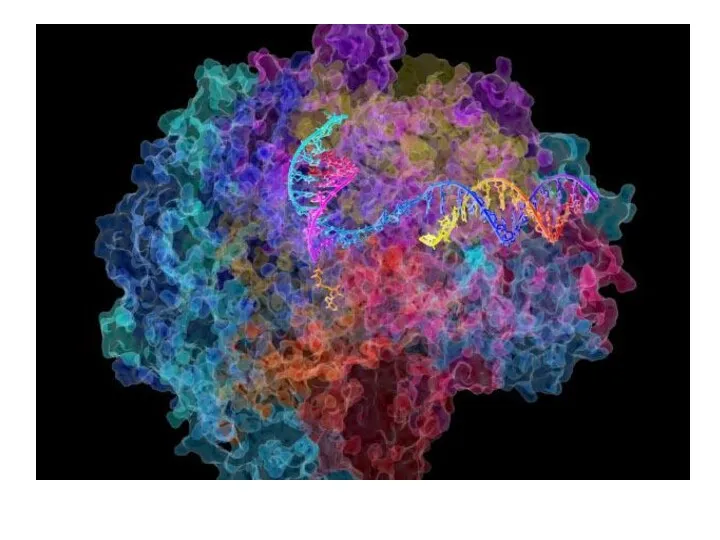

РНК-полимеразы не имеют независимой З'→ 5' -экзонуклеазной активности для исправления ошибок,
РНК-полимеразы не имеют независимой З'→ 5' -экзонуклеазной активности для исправления ошибок,

Транскрипция
В процессе транскрипции различают стадии:
инициации
элонгации
терминации
Транскрипция
В процессе транскрипции различают стадии:
инициации
элонгации
терминации

Процесс инициации транскрипции состоит из двух основных этапов:
связывания РНК-полимеразы с ДНК.
собственно
Процесс инициации транскрипции состоит из двух основных этапов:
связывания РНК-полимеразы с ДНК.
собственно

Первый этап:
σ- субъединица временно связывается с кор-ферментом и в составе холофермента
Первый этап:
σ- субъединица временно связывается с кор-ферментом и в составе холофермента

В составе бактериальных промоторов выделяют консервативные последовательности.
Эти последовательности называют консенсусными последовательностями
В составе бактериальных промоторов выделяют консервативные последовательности.
Эти последовательности называют консенсусными последовательностями

Эффективность связывания РНК-полимеразы с промотором определяется консенсусными последовательностями, расстоянием между ними
Эффективность связывания РНК-полимеразы с промотором определяется консенсусными последовательностями, расстоянием между ними

В состав некоторых промоторов входит богатый АТ основаниями UP-элемент (upstream promoter).
У
В состав некоторых промоторов входит богатый АТ основаниями UP-элемент (upstream promoter).
У

Промотор не несет информации и служит для присоединения и ориентации РНК-полимеразы.
Определяется
Промотор не несет информации и служит для присоединения и ориентации РНК-полимеразы.
Определяется

Второй этап инициации:
в результате холофермент РНК-полимераза связывается с ДНК (интактная) и
Второй этап инициации:
в результате холофермент РНК-полимераза связывается с ДНК (интактная) и

Закрытый промоторный комплекс:
В результате раскручивания цепей ДНК разрываются водородные связи между
Закрытый промоторный комплекс:
В результате раскручивания цепей ДНК разрываются водородные связи между

Эти короткие фрагменты РНК провоцируют разрыв контактов РНК-полимеразы с промотором.
σ- субъединица
Эти короткие фрагменты РНК провоцируют разрыв контактов РНК-полимеразы с промотором.
σ- субъединица

На стадии элонгации кор-фермент РНК-полимераза осуществляет поэтапное присоединение нуклеотидов, комплементарных матричной
На стадии элонгации кор-фермент РНК-полимераза осуществляет поэтапное присоединение нуклеотидов, комплементарных матричной

Механизм присоединения нуклеотидов:
связывание входящего нуклеотида.
если нуклеотид комплементарен нуклеотиду матричной цепи ДНК,
Механизм присоединения нуклеотидов:
связывание входящего нуклеотида.
если нуклеотид комплементарен нуклеотиду матричной цепи ДНК,

Транслокация: перемещение РНК-полимеразы на 1 нуклеотид вперед по матрице ДНК.
Присоединение следующего
Транслокация: перемещение РНК-полимеразы на 1 нуклеотид вперед по матрице ДНК.
Присоединение следующего

Рифампицин связывается с β-субъединицей бактериальной РНК-полимеразы, препятствуя высвобождению промотора при транскрипции.
Рифампицин связывается с β-субъединицей бактериальной РНК-полимеразы, препятствуя высвобождению промотора при транскрипции.

Терминация
E.coli имеет два типа сигналов терминации:
зависимый от присутствия ρ-белкового фактора
ρ-независимый
Терминация
E.coli имеет два типа сигналов терминации:
зависимый от присутствия ρ-белкового фактора
ρ-независимый

ρ-независимые терминаторы имеют два свойства:
первое: содержат последовательность, транскрипт которой имеет комплементарные
ρ-независимые терминаторы имеют два свойства:
первое: содержат последовательность, транскрипт которой имеет комплементарные

Во-вторых: их матричная цепь содержит консервативную последовательность из трех остатков А,
Во-вторых: их матричная цепь содержит консервативную последовательность из трех остатков А,

Взаимодействие новосинтезированного олиго U-участка РНК с РНК-полимеразой приводит к возникновению транскрипционных
Взаимодействие новосинтезированного олиго U-участка РНК с РНК-полимеразой приводит к возникновению транскрипционных

Синтез ~80% транскриптов E.coli терминируется по ρ-независимому пути.
ρ-зависимый путь: необходим
Синтез ~80% транскриптов E.coli терминируется по ρ-независимому пути.
ρ-зависимый путь: необходим

ρ-фактор связывается с РНК в специальных пиримидин-богатых сайтах RUT (Rho utilization)
ρ-фактор связывается с РНК в специальных пиримидин-богатых сайтах RUT (Rho utilization)

ρ-фактор настигает транскрипционный комплекс на сайте терминации, пирофосфат перестает выделяться, ρ-фактор
ρ-фактор настигает транскрипционный комплекс на сайте терминации, пирофосфат перестает выделяться, ρ-фактор
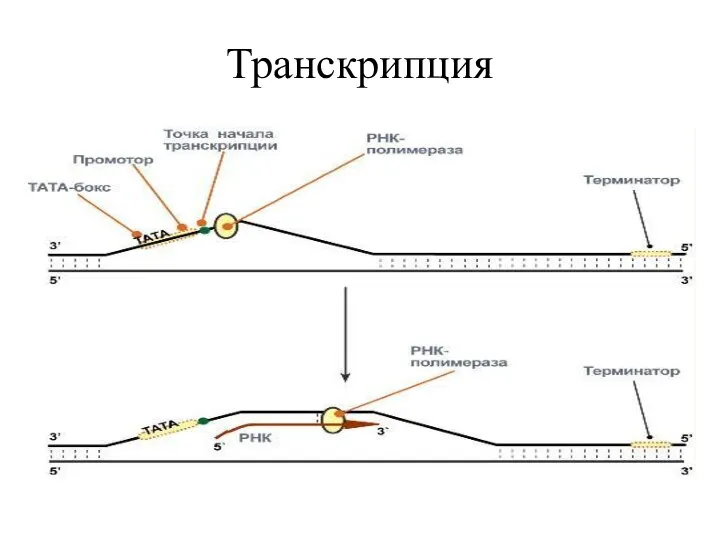
Транскрипция
Транскрипция

В клетках эукариот обнаружено 3 вида РНК-полимераз (I, II, III).
состав комплекса
каждая
В клетках эукариот обнаружено 3 вида РНК-полимераз (I, II, III).
состав комплекса
каждая

POL I – синтез прерибосомной РНК (пре-рРНК)
POL II – синтез
POL I – синтез прерибосомной РНК (пре-рРНК)
POL II – синтез

POL III образует молекулы тРНК, 5S рРНК и другие небольшие специализированные
POL III образует молекулы тРНК, 5S рРНК и другие небольшие специализированные

POL II играет ведущую роль в экспрессии эукариотических генов.
состоит из 12
POL II играет ведущую роль в экспрессии эукариотических генов.
состоит из 12

RBP1 – гомология с β' - субъединицой прокариотической РНК-полимеразы.
RBP2 – гомология
RBP1 – гомология с β' - субъединицой прокариотической РНК-полимеразы.
RBP2 – гомология

Для формирования активного транскрипционного комплекса POL II нуждается в ряде белков
Для формирования активного транскрипционного комплекса POL II нуждается в ряде белков

инициация
ТАТА-связывающий белок (ТВР) взамодействует с ТАТА-боксом.
ТВР связывается с фактором транскрипции TFIIB,
инициация
ТАТА-связывающий белок (ТВР) взамодействует с ТАТА-боксом.
ТВР связывается с фактором транскрипции TFIIB,

комплекс TFIIB- ТВР связывается с другим комплексом TFIIF- POL II.
TFIIF помогает
комплекс TFIIB- ТВР связывается с другим комплексом TFIIF- POL II.
TFIIF помогает

TFIIH обладает хеликазной активностью и начинает раскручивание ДНК вблизи точки начала
TFIIH обладает хеликазной активностью и начинает раскручивание ДНК вблизи точки начала

Фосфорилирование POL II приводит к конформационным изменениям и инциирует транскрипцию.
По мере
Фосфорилирование POL II приводит к конформационным изменениям и инциирует транскрипцию.
По мере

На стадии элонгации TFIIF- POL II остаются в комплексе.
Усиливается факторами элонгации,
На стадии элонгации TFIIF- POL II остаются в комплексе.
Усиливается факторами элонгации,

В фазе терминации POL II освобождается, дефосфорилируется, и может участвовать в
В фазе терминации POL II освобождается, дефосфорилируется, и может участвовать в

Бледная поганка синтезирует α-аманитин, который прерывает образование мРНК в клетках, блокируя
Бледная поганка синтезирует α-аманитин, который прерывает образование мРНК в клетках, блокируя

Процессинг РНК
Многие молекулы РНК бактерий и практически все молекулы РНК эукариот
Процессинг РНК
Многие молекулы РНК бактерий и практически все молекулы РНК эукариот

процессинг матричной РНК
Модификация 5'-конца:
Модификации пре-мРНК начинаются на стадии элонгации.
процессинг матричной РНК
Модификация 5'-конца:
Модификации пре-мРНК начинаются на стадии элонгации.

процессинг матричной РНК
Модифицированный 5'-конец удлиняет время жизни мРНК, защищая её от
процессинг матричной РНК
Модифицированный 5'-конец удлиняет время жизни мРНК, защищая её от

процессинг матричной РНК
Модификация 3'-конца
3'-конец большинства транскриптов, также подвергается модификации, при
процессинг матричной РНК
Модификация 3'-конца
3'-конец большинства транскриптов, также подвергается модификации, при

процессинг матричной РНК
полиА- «хвост» эукариот защищает мРНК от ферментативного расщепления.
Многие молекулы
процессинг матричной РНК
полиА- «хвост» эукариот защищает мРНК от ферментативного расщепления.
Многие молекулы

Первичные транскрипты эукариот строго комплементарны матрице (ДНК), содержат как экзоны, так
Первичные транскрипты эукариот строго комплементарны матрице (ДНК), содержат как экзоны, так

первичные транскрипты прокариот также могут содержать интроны (бактерии, археи).
первичные транскрипты прокариот также могут содержать интроны (бактерии, археи).

в процессе сплайсинга («сплайсинг» от англ, to splice - сращивать) интроны
в процессе сплайсинга («сплайсинг» от англ, to splice - сращивать) интроны

процессинг матричной РНК
сплайсинг происходит в ядре, в цитоплазму поступает уже "зрелая"
процессинг матричной РНК
сплайсинг происходит в ядре, в цитоплазму поступает уже "зрелая"

аутосплайсинг
Процесс "вырезания" некоторых интронов протекает при участии самих РНК.
впервые это было
аутосплайсинг
Процесс "вырезания" некоторых интронов протекает при участии самих РНК.
впервые это было
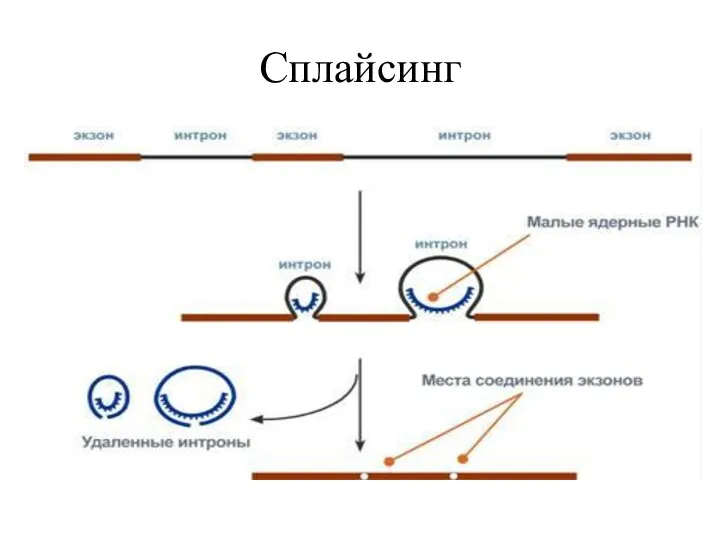
Сплайсинг
Сплайсинг

Большинство интронов являются сплайсосомными интронами.
Удаление происходит и катализируется внутри крупного белкового
Большинство интронов являются сплайсосомными интронами.
Удаление происходит и катализируется внутри крупного белкового

Сплайсосома состоит из специализированных РНК-белковых комплексов – малых ядерных рибонуклеопротеинов (мяРНП).
Каждый
Сплайсосома состоит из специализированных РНК-белковых комплексов – малых ядерных рибонуклеопротеинов (мяРНП).
Каждый

В реакциях сплайсинга участвуют 5 мяРНК (ядро эукариот) и примерно 50
В реакциях сплайсинга участвуют 5 мяРНК (ядро эукариот) и примерно 50

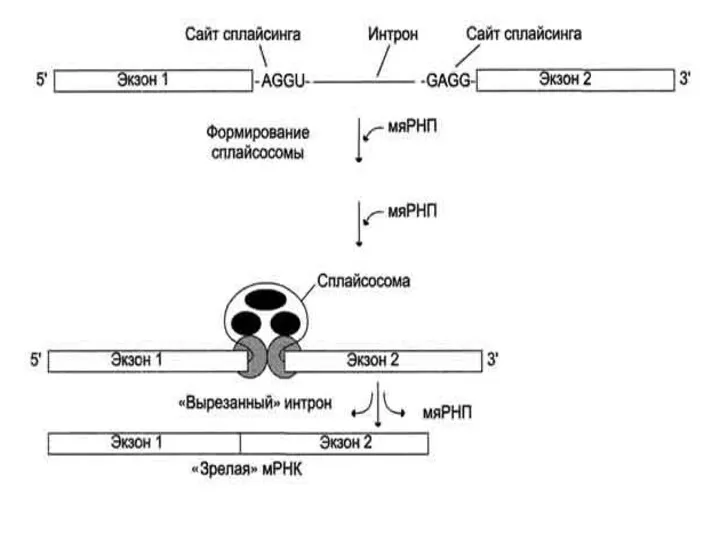
на 5' конце сплайсосомные интроны имеют GU-последовательность.
на 3' конце – AG-последовательность.
Эти
на 5' конце сплайсосомные интроны имеют GU-последовательность.
на 3' конце – AG-последовательность.
Эти

Сложность организмов не коррелирует с количеством генов, кодирующих белки.
Из некоторых транскриптов
Сложность организмов не коррелирует с количеством генов, кодирующих белки.
Из некоторых транскриптов

Альтернативный сплайсинг
В результате процессинга могут образовываться различные мРНК из первичного транскрипта.
Первичный
Альтернативный сплайсинг
В результате процессинга могут образовываться различные мРНК из первичного транскрипта.
Первичный

Альтернативные варианты сплайсинга РНК могут использоваться для одновременного синтеза двух разных
Альтернативные варианты сплайсинга РНК могут использоваться для одновременного синтеза двух разных

Многие гены у млекопитающих подвергаются альтернативному сплайсингу, что значительно увеличивает количество
Многие гены у млекопитающих подвергаются альтернативному сплайсингу, что значительно увеличивает количество

Число идентифицированных классов специализированных РНК быстро растет: малая ядерная РНК, малая
Число идентифицированных классов специализированных РНК быстро растет: малая ядерная РНК, малая

Регулируют функции мРНК путем ее расщепления или подавления ее трансляции.
Обнаружены у
Регулируют функции мРНК путем ее расщепления или подавления ее трансляции.
Обнаружены у

Сплайсинг
Нобелевская премия 1993 года была присуждена Р.Дж.Робертсу и Ф.А.Шарпу (Richard J.Roberts,
Сплайсинг
Нобелевская премия 1993 года была присуждена Р.Дж.Робертсу и Ф.А.Шарпу (Richard J.Roberts,
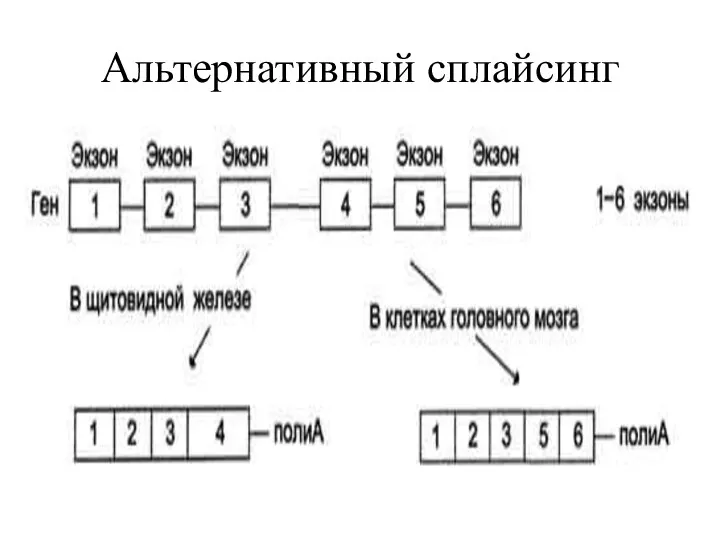
Альтернативный сплайсинг
Альтернативный сплайсинг

Регуляция транскрипции
Потребности в продуктах любых генов изменяются в зависимости от состояния
Регуляция транскрипции
Потребности в продуктах любых генов изменяются в зависимости от состояния

Транскрипция –первая стадия в сложном и энергозатратном процессе синтеза белка, поэтому
Транскрипция –первая стадия в сложном и энергозатратном процессе синтеза белка, поэтому

регуляция может происходить на любой стадии транскрипции, но чаще на стадии
регуляция может происходить на любой стадии транскрипции, но чаще на стадии

регулируемой скоростью синтеза и деградации РНК.
постсинтетическими модификациями, переводящими σ- субъединицу из
регулируемой скоростью синтеза и деградации РНК.
постсинтетическими модификациями, переводящими σ- субъединицу из

специализированным классом анти σ- белков, связывающихся с определенным типом σ- субъединиц
специализированным классом анти σ- белков, связывающихся с определенным типом σ- субъединиц

Решающим фактором, управляющим экспрессией генов является концентрация в клетке соответствующей мРНК,
Решающим фактором, управляющим экспрессией генов является концентрация в клетке соответствующей мРНК,

Скорость расщепления мРНК разных эукариотических генов различаются- от нескольких секунд до
Скорость расщепления мРНК разных эукариотических генов различаются- от нескольких секунд до

мРНК разрушается рибонуклеазами, присутствующими во всех клетках.
Структура бактериальной мРНК с ρ-независимым
мРНК разрушается рибонуклеазами, присутствующими во всех клетках.
Структура бактериальной мРНК с ρ-независимым

Спасибо за внимание
Спасибо за внимание

Транскрипция
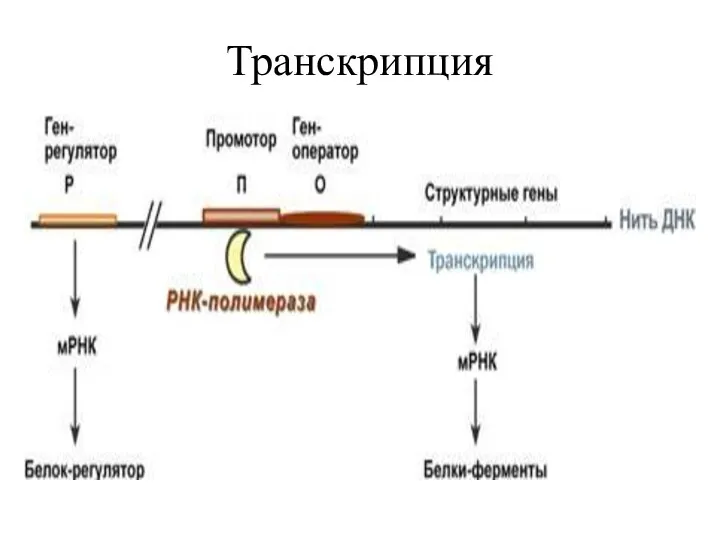
Транскрипция ДНК происходит отдельными участками, в которые входит один или несколько
Транскрипция
Транскрипция ДНК происходит отдельными участками, в которые входит один или несколько
Транскрипция
Транскрипция

Транскрипция
Транскриптон бактерий называется опероном
Транскрипция
Транскриптон бактерий называется опероном
 Клеточные структуры и их функции
Клеточные структуры и их функции Взаимодействие неаллельных генов. Цитоплазматическая наследственность
Взаимодействие неаллельных генов. Цитоплазматическая наследственность Репродукція клітин
Репродукція клітин ОГЭ по биологии
ОГЭ по биологии Акселерация как социальная и гигиеническая проблема
Акселерация как социальная и гигиеническая проблема Место животных в системе живого мира
Место животных в системе живого мира Нәруыз. Пептидті байланыспен қосылған аминқышқылдардан тұратын құрылым
Нәруыз. Пептидті байланыспен қосылған аминқышқылдардан тұратын құрылым Физиология растений
Физиология растений Теорія Мальтуса та сучасність
Теорія Мальтуса та сучасність Бобовые. Особенности строения вегетативных органов
Бобовые. Особенности строения вегетативных органов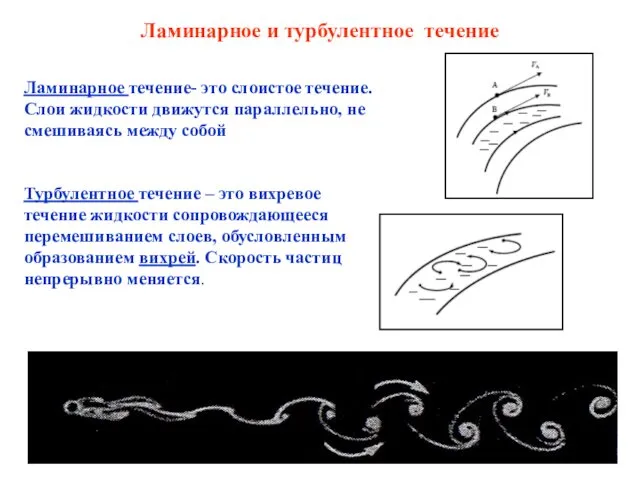 Ламинарное и турбулентное течение. Скорость течения крови в разных отделах кровеносной системы. (Лекции 4 и 6)
Ламинарное и турбулентное течение. Скорость течения крови в разных отделах кровеносной системы. (Лекции 4 и 6) Жизнь организмов в морях и океанах
Жизнь организмов в морях и океанах Основные понятия генетики
Основные понятия генетики Молекулярная биология
Молекулярная биология Тип Членистоногие
Тип Членистоногие Лабораторне заняття № 4. М’язи тулуба. Діафрагма. Утворення стінок черевної порожнини
Лабораторне заняття № 4. М’язи тулуба. Діафрагма. Утворення стінок черевної порожнини Сравнение клеток растений и животных. Отличия между клетками
Сравнение клеток растений и животных. Отличия между клетками Хлор и его роль в организме человека
Хлор и его роль в организме человека Тренажёр Вода
Тренажёр Вода Цепи питания
Цепи питания Необычное животное хамелеон
Необычное животное хамелеон Классификация и свойства бактерий
Классификация и свойства бактерий Типы питания. Автотрофы и гетеротрофы
Типы питания. Автотрофы и гетеротрофы Происхождение митохондрий
Происхождение митохондрий Фитопатогенді бактериялардың таксономикалық бөлімдері
Фитопатогенді бактериялардың таксономикалық бөлімдері Тип: моллюски. Класс: двустворчатые. Устрица
Тип: моллюски. Класс: двустворчатые. Устрица Ферменты - 2. Эффекторы
Ферменты - 2. Эффекторы Земноводні. Види земноводних
Земноводні. Види земноводних