- Метилирование мтДНК. ЛЕКЦИЯ 4

Содержание
- 2. Метилирование ДНК
- 3. Метилирование ядерной ДНК Происходит преимущественно в CpG-участках ДНК Служит для регуляции транскрипции: метилирование ДНК в СрG-участках
- 4. Перед транскрипционно активными генами обычно существуют неметилированные CрG островки. CрG в неактивных генах обычно метилированы для
- 5. В ядерной ДНК при эмбриогенезе метилирование осуществляют метилазы DNMT 3a и 3b Поддерживает паттерны метилирования в
- 6. Изначальные паттерны метилирования создаются ДНК метилтрансферазами DNMT3a и 3b, активно работающими в эмбриогенезе. DNMT3L, видимо, функционирует
- 7. Под воздействием ферментов семейства ТЕТ (ten eleven translocation) происходит окисление 5mC с образованием 5-гидроксиметилцитозина (5hmC)
- 8. Структура и механизм работы ферментов семейства ТЕТ (ten eleven translocation) PMID: 24153300
- 9. PMID: 24153300
- 10. PMID: 24153300
- 11. Стохастическая модель метилирования метилирование в каждом сайте - результат двух противоположных процессов – метилирования и деметилирования,
- 12. Бисульфитное секвенирование
- 14. Метилирование мтДНК Открыто в 1974 году До 2019 года были неизвестны точные места метилирования Неизвестна функциональная
- 15. Митохондриальная форма DNMT1 PMID:21321201
- 16. Лидерные пептиды DNMT1 слитые с GFP локализованы в митохондриях PMID:21321201
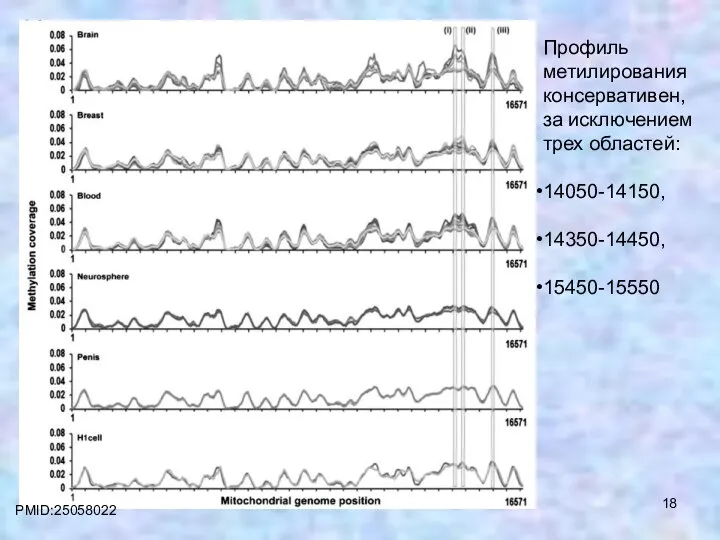
- 17. PMID:25058022 Распределение 5mC по митохондриальному геному человека в разных клеточных линиях и тканях
- 18. PMID:25058022 Профиль метилирования консервативен, за исключением трех областей: 14050-14150, 14350-14450, 15450-15550
- 19. Метилирование мтДНК происходит в основном не по CpG-сайтам PMID: 31665742
- 20. Метилирование мтДНК происходит по-разному в нормальных и раковых клетках А – гепатоциты В – раковые клетки
- 21. Уровень метилирования мтДНК повышается в ходе процессов развития PMID: 31417147 Синий – зигота Красный – бластоциста
- 22. Метилирование мтДНК в области D-loop преимущественно происходит не в СpG последовательностях. Метилирование в D-loop в основном
- 23. Одновременная инактивация DNMT3a, 3b и DNMT1 уменьшает CpG метилирование в D-loop и слабо влияет на метилирование
- 24. Как и в ядерной ДНК в мтДНК мало динуклеотидов CpG – 435 на 16659 4747 остатков
- 25. CpG в области TERM защищены от метилирования (in vitro) предположительно из-за связывания с белком MTERF –
- 26. Возможная регуляция репликации с помощью метилирования PMID:25524586
- 27. Когда происходят изменения в метилировании мтДНК? В ответ на изменения во внешней среде (загрязнение воздуха, окислительный
- 28. Гиперметилирование генов 12S рРНК, Phe-тРНК и области D-петли наблюдалось у рабочих, профессиональная деятельность которых связана с
- 29. p53 снижает экспрессию mtDNMT1 в клетках без р53 количество мтDNMT1 увеличивается в 6 раз PMID: 21321201
- 30. ND6 (L-цепь) ↑ ND1 (H-цепь) ↓ В р53-/- клетках изменена экспрессия некоторых генов мтДНК PMID:21321201
- 31. Возможная связь метилирования мт ДНК с онкологическими заболеваниями Метилирование мтДНК Изменение (качественное или количественное) экспрессии мт-генов
- 32. Метилирование цитозина в тканях мозга человека уменьшается в соответствии со стадией развития в областях перед генами
- 33. Дыхательная цепь пожилых людей работает менее эффективно, чем в молодости, что приводит к формированию «старческого» фенотипа
- 34. В образовании глицина в митохондриях участвует также продукт гена SHMT2 – сериновая гидроксиметилтрансфераза. Сравнили количества мРНК
- 35. С возрастом происходит изменение экспрессии генов, продукты которых вовлечены в митохондриальный метаболизм, в частности, в образование
- 36. митохондрия Возрастные эпигенетические изменения в митохондриях и ядре →изменение экспрессии ядерных и/или митохондриальных генов →изменение метаболизма
- 37. Уровень метилированности мтДНК статистически значимо снижается с возрастом PMID: 31537639
- 39. Скачать презентацию
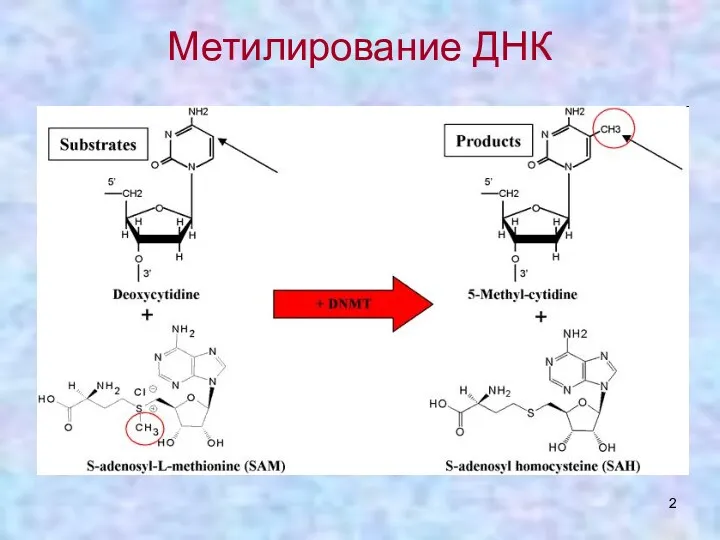
Метилирование ДНК
Метилирование ДНК
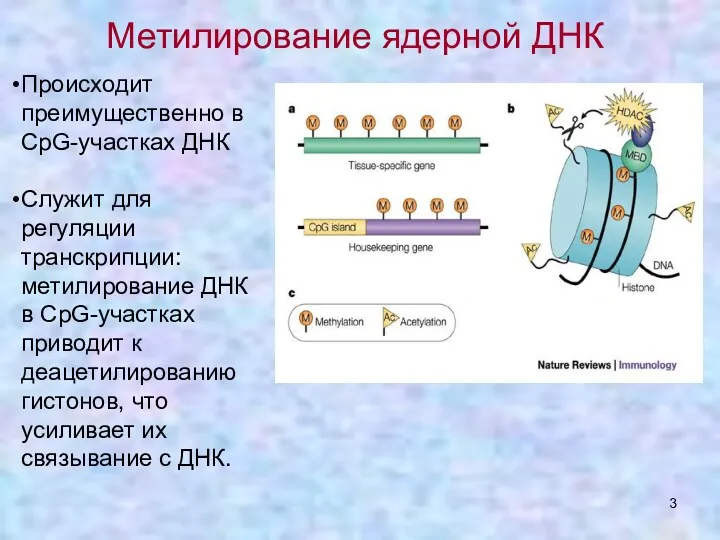
Метилирование ядерной ДНК
Происходит преимущественно в CpG-участках ДНК
Служит для регуляции транскрипции: метилирование
Метилирование ядерной ДНК
Происходит преимущественно в CpG-участках ДНК
Служит для регуляции транскрипции: метилирование
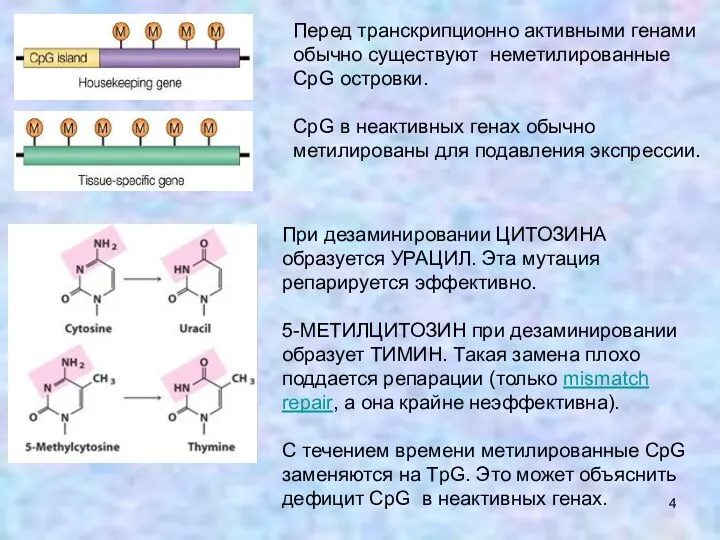
Перед транскрипционно активными генами обычно существуют неметилированные CрG островки.
CрG в неактивных
Перед транскрипционно активными генами обычно существуют неметилированные CрG островки.
CрG в неактивных
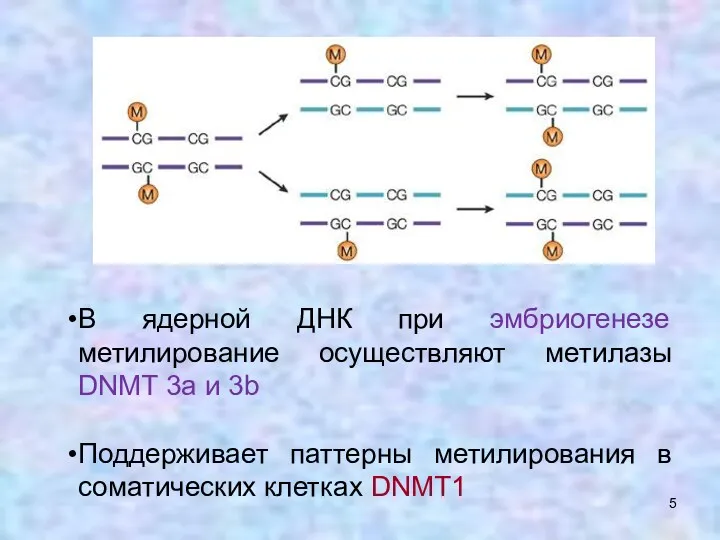
В ядерной ДНК при эмбриогенезе метилирование осуществляют метилазы DNMT 3a и
В ядерной ДНК при эмбриогенезе метилирование осуществляют метилазы DNMT 3a и

Изначальные паттерны метилирования создаются ДНК метилтрансферазами DNMT3a и 3b, активно работающими
Изначальные паттерны метилирования создаются ДНК метилтрансферазами DNMT3a и 3b, активно работающими
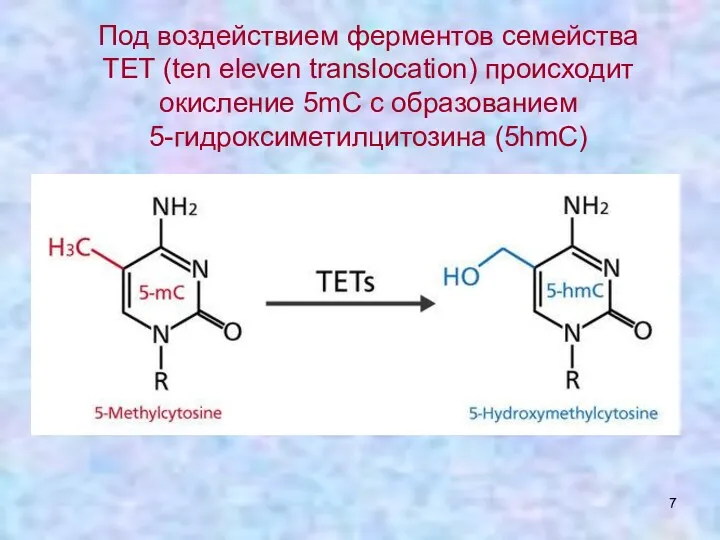
Под воздействием ферментов семейства
ТЕТ (ten eleven translocation) происходит окисление 5mC с
Под воздействием ферментов семейства
ТЕТ (ten eleven translocation) происходит окисление 5mC с
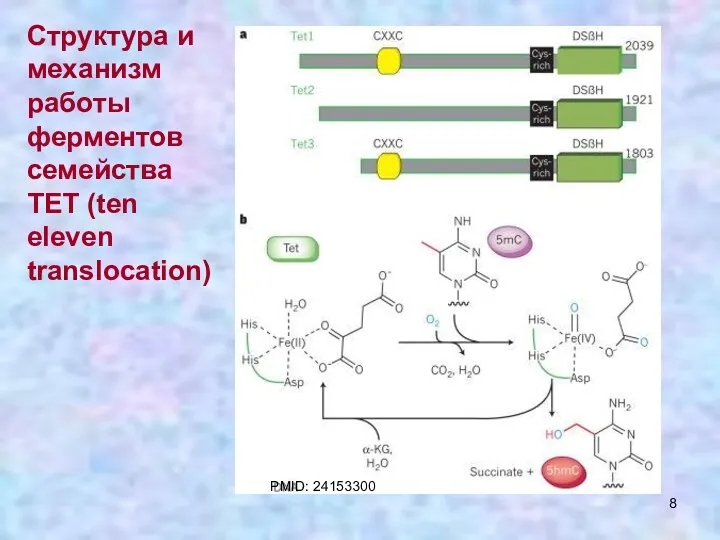
Структура и механизм работы ферментов семейства
ТЕТ (ten eleven translocation)
PMID: 24153300
Структура и механизм работы ферментов семейства
ТЕТ (ten eleven translocation)
PMID: 24153300
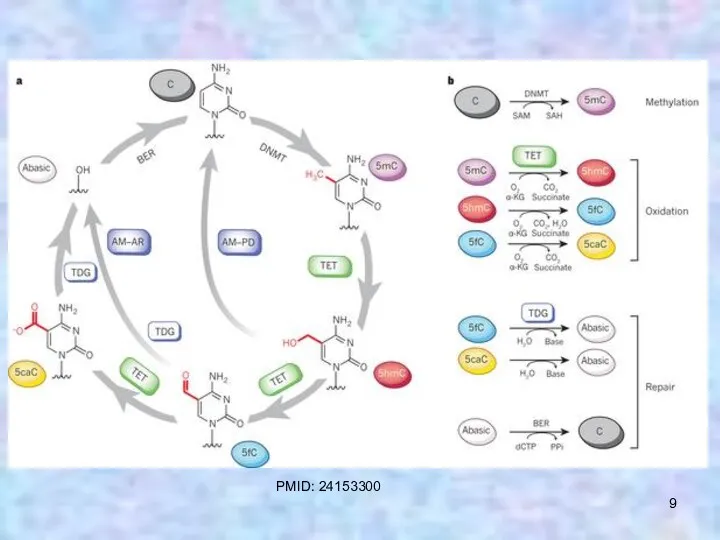
PMID: 24153300
PMID: 24153300
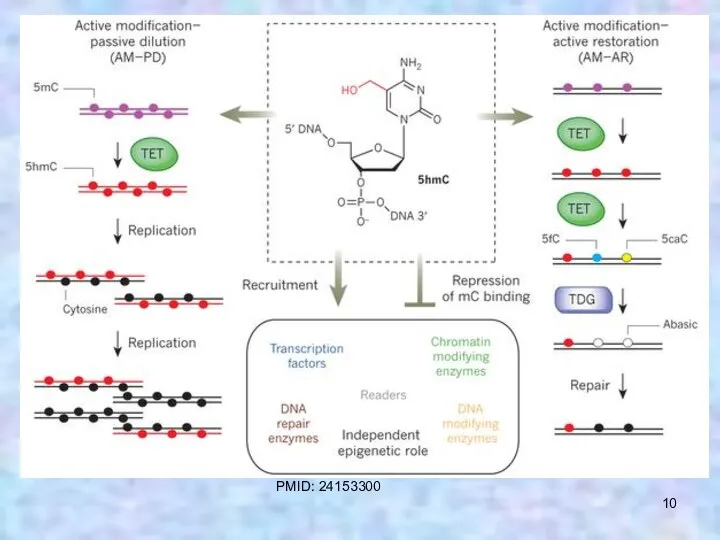
PMID: 24153300
PMID: 24153300

Стохастическая модель метилирования
метилирование в каждом сайте - результат двух противоположных процессов
Стохастическая модель метилирования
метилирование в каждом сайте - результат двух противоположных процессов
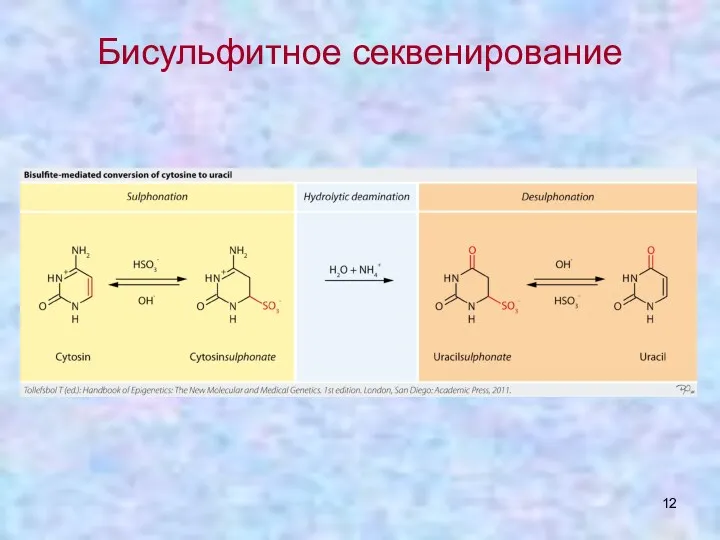
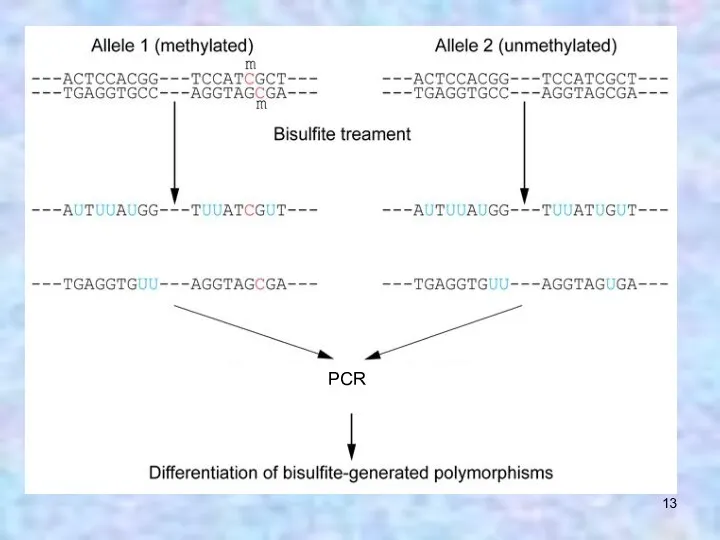
Бисульфитное секвенирование
Бисульфитное секвенирование
Метилирование мтДНК
Открыто в 1974 году
До 2019 года были неизвестны точные места
Метилирование мтДНК
Открыто в 1974 году
До 2019 года были неизвестны точные места

Митохондриальная форма DNMT1
PMID:21321201
Митохондриальная форма DNMT1
PMID:21321201
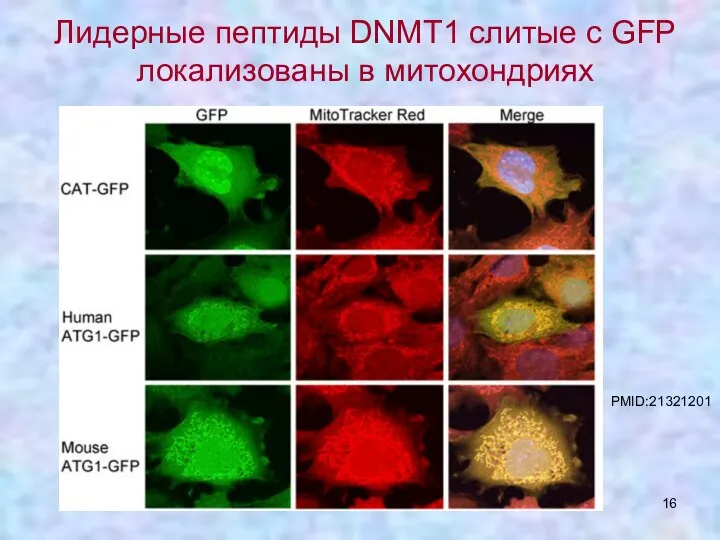
Лидерные пептиды DNMT1 слитые с GFP локализованы в митохондриях
PMID:21321201
Лидерные пептиды DNMT1 слитые с GFP локализованы в митохондриях
PMID:21321201

PMID:25058022
Распределение 5mC по митохондриальному геному человека в разных клеточных линиях и
Распределение 5mC по митохондриальному геному человека в разных клеточных линиях и
PMID:25058022
Профиль метилирования консервативен, за исключением трех областей:
14050-14150,
14350-14450,
15450-15550
Профиль метилирования консервативен, за исключением трех областей:
14050-14150,
14350-14450,
15450-15550
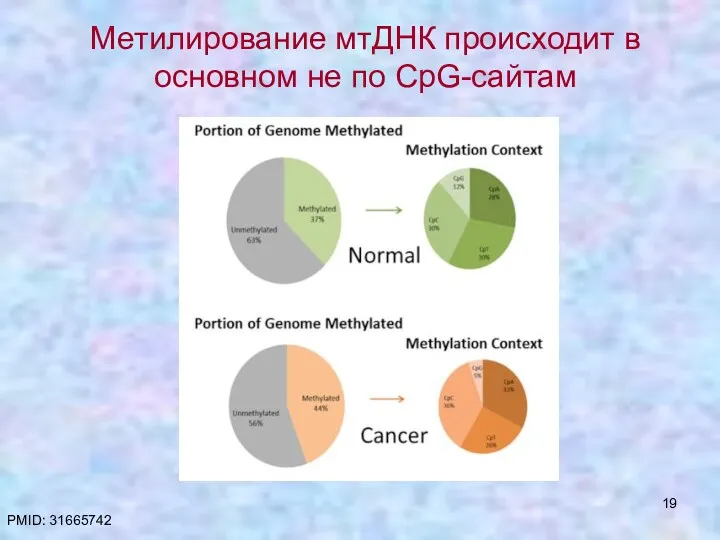
Метилирование мтДНК происходит в основном не по CpG-сайтам
PMID: 31665742
Метилирование мтДНК происходит в основном не по CpG-сайтам
PMID: 31665742

Метилирование мтДНК происходит по-разному в нормальных и раковых клетках
А – гепатоциты
В
Метилирование мтДНК происходит по-разному в нормальных и раковых клетках
А – гепатоциты
В
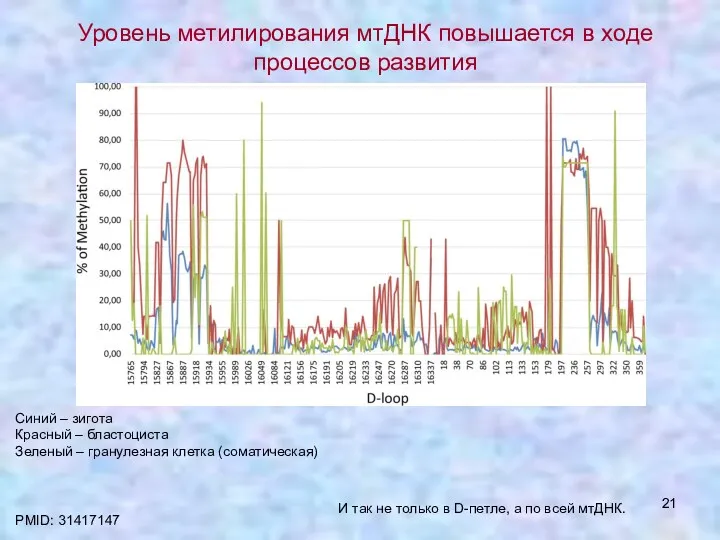
Уровень метилирования мтДНК повышается в ходе процессов развития
PMID: 31417147
Синий – зигота
Красный –
Уровень метилирования мтДНК повышается в ходе процессов развития
PMID: 31417147
Синий – зигота
Красный –
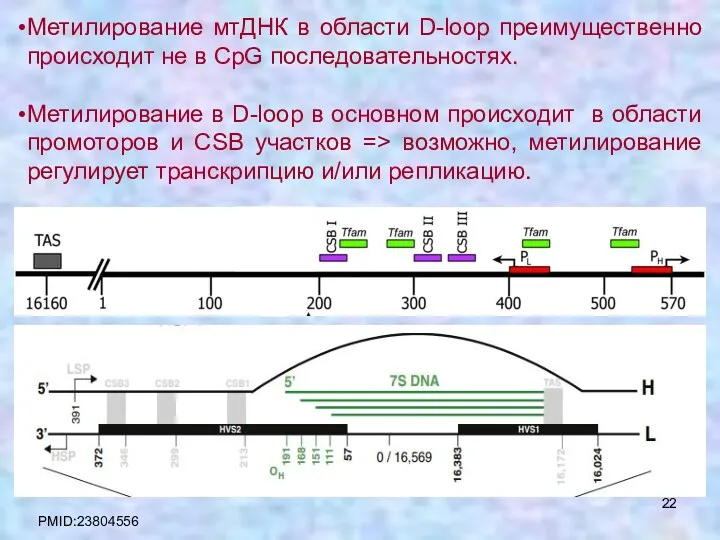
Метилирование мтДНК в области D-loop преимущественно происходит не в СpG последовательностях.
Метилирование
Метилирование мтДНК в области D-loop преимущественно происходит не в СpG последовательностях.
Метилирование

Одновременная инактивация DNMT3a, 3b и DNMT1 уменьшает CpG метилирование в D-loop
Одновременная инактивация DNMT3a, 3b и DNMT1 уменьшает CpG метилирование в D-loop
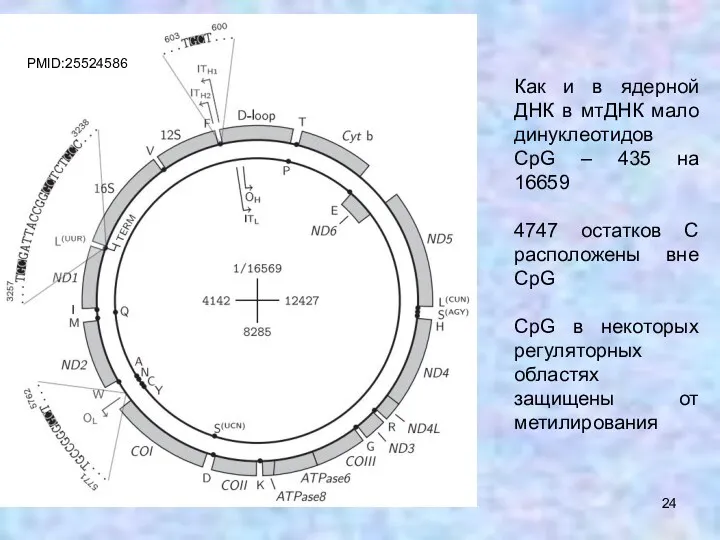
Как и в ядерной ДНК в мтДНК мало динуклеотидов CpG –
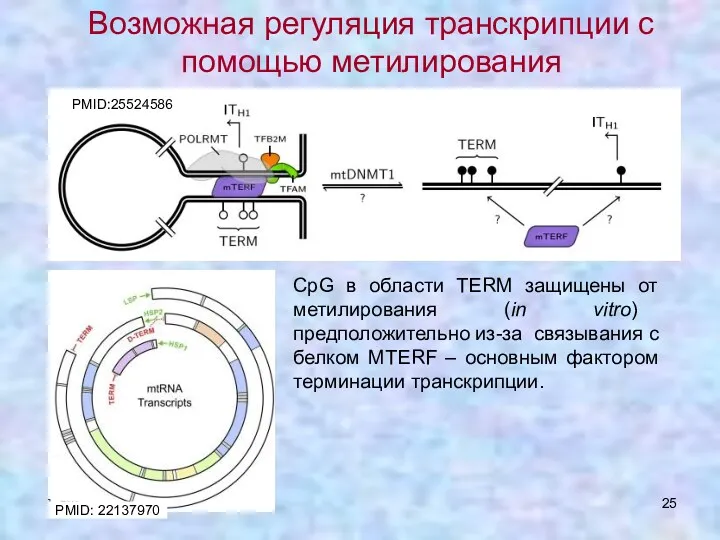
CpG в области TERM защищены от метилирования (in vitro) предположительно из-за
CpG в области TERM защищены от метилирования (in vitro) предположительно из-за
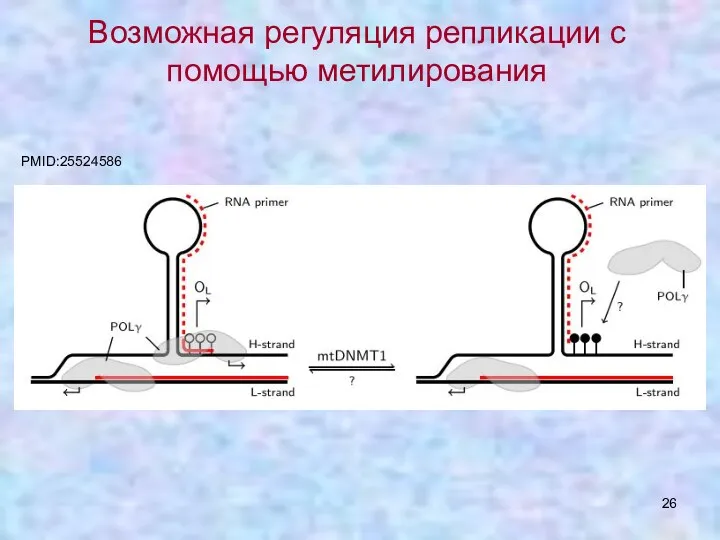
Возможная регуляция репликации с помощью метилирования
PMID:25524586
Возможная регуляция репликации с помощью метилирования
PMID:25524586

Когда происходят изменения в метилировании мтДНК?
В ответ на изменения во
Когда происходят изменения в метилировании мтДНК?
В ответ на изменения во

Гиперметилирование генов 12S рРНК, Phe-тРНК и области D-петли наблюдалось у рабочих,
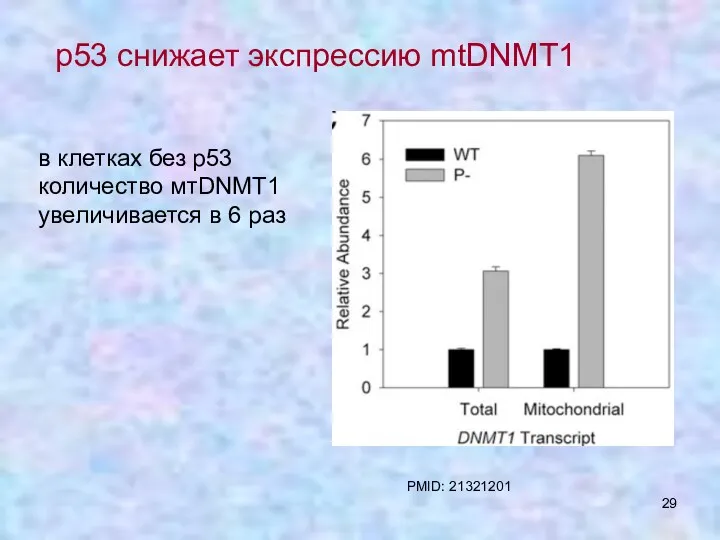
p53 снижает экспрессию mtDNMT1
в клетках без р53 количество мтDNMT1 увеличивается в
p53 снижает экспрессию mtDNMT1
в клетках без р53 количество мтDNMT1 увеличивается в
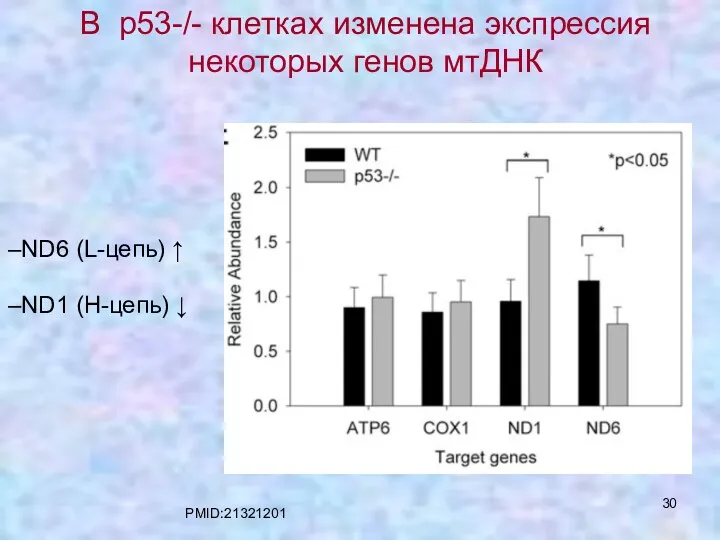
ND6 (L-цепь) ↑
ND1 (H-цепь) ↓
В р53-/- клетках изменена экспрессия некоторых генов
ND6 (L-цепь) ↑
ND1 (H-цепь) ↓
В р53-/- клетках изменена экспрессия некоторых генов
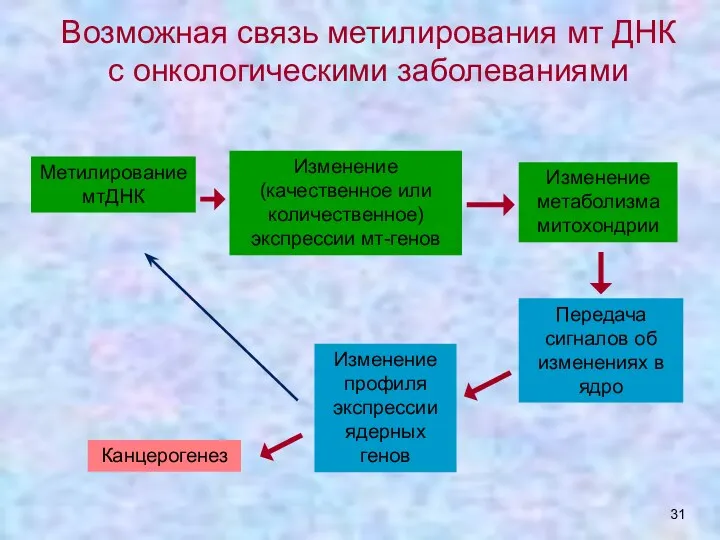
Возможная связь метилирования мт ДНК с онкологическими заболеваниями
Метилирование мтДНК
Изменение (качественное
Возможная связь метилирования мт ДНК с онкологическими заболеваниями
Метилирование мтДНК
Изменение (качественное

Метилирование цитозина в тканях мозга человека уменьшается в соответствии со стадией
Метилирование цитозина в тканях мозга человека уменьшается в соответствии со стадией

Дыхательная цепь пожилых людей работает менее эффективно, чем в молодости, что
Дыхательная цепь пожилых людей работает менее эффективно, чем в молодости, что

В образовании глицина в митохондриях участвует также продукт гена SHMT2 –
В образовании глицина в митохондриях участвует также продукт гена SHMT2 –

С возрастом происходит изменение экспрессии генов, продукты которых вовлечены в митохондриальный
С возрастом происходит изменение экспрессии генов, продукты которых вовлечены в митохондриальный
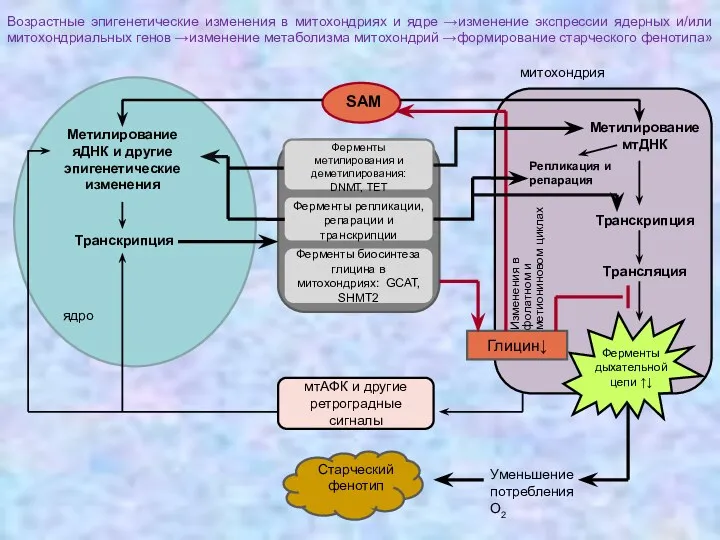
митохондрия
Возрастные эпигенетические изменения в митохондриях и ядре →изменение экспрессии ядерных и/или
митохондрия
Возрастные эпигенетические изменения в митохондриях и ядре →изменение экспрессии ядерных и/или
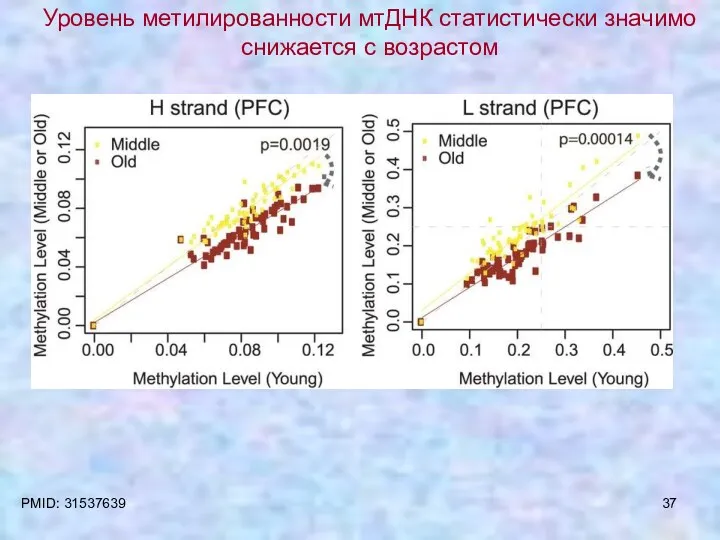
Уровень метилированности мтДНК статистически значимо снижается с возрастом
PMID: 31537639
Уровень метилированности мтДНК статистически значимо снижается с возрастом
PMID: 31537639
 Биоэлектрогенез. Строение и физические свойства биологических мембран
Биоэлектрогенез. Строение и физические свойства биологических мембран Пищеварительная система
Пищеварительная система Структурная организация микробной клетки
Структурная организация микробной клетки Биотехнология как источник био-риска
Биотехнология как источник био-риска Фізіологія. Механізми регуляції
Фізіологія. Механізми регуляції Систематика Покритонасінних рослин. Відділ Магноліофіти, Квіткові або Покритонасінні
Систематика Покритонасінних рослин. Відділ Магноліофіти, Квіткові або Покритонасінні Общая физиология сенсорных систем
Общая физиология сенсорных систем Отряд голенастые
Отряд голенастые Белгілерді ң тұқым қуалауының негізгі зандары, т ұқым қуалайтын белгілерді ңберілу ережелері
Белгілерді ң тұқым қуалауының негізгі зандары, т ұқым қуалайтын белгілерді ңберілу ережелері Правила техники безопасности в кабинете биологии
Правила техники безопасности в кабинете биологии Анатомия нервной системы
Анатомия нервной системы Тип членистоногие. Класс насекомые
Тип членистоногие. Класс насекомые Тип Членистоногие
Тип Членистоногие Листериоз. Род Listeria
Листериоз. Род Listeria Интегрированный урок по биологии,химии и физике. Викторина Колосок
Интегрированный урок по биологии,химии и физике. Викторина Колосок Дыхательная система. Органы дыхания
Дыхательная система. Органы дыхания Поведение собак
Поведение собак Основы селекции. Работы Н.И. Вавилова
Основы селекции. Работы Н.И. Вавилова Краткая история изучения клетки
Краткая история изучения клетки Обмен веществ. Возможные превращения веществ
Обмен веществ. Возможные превращения веществ Тканевые элементы нервной системы
Тканевые элементы нервной системы Какие функции выполняет корень?
Какие функции выполняет корень? Лисиця. Представниця класу ссавці, ряду хижі звірі
Лисиця. Представниця класу ссавці, ряду хижі звірі Анатомия крысы
Анатомия крысы Зимующие птицы
Зимующие птицы ГМО в пищевой промышленности
ГМО в пищевой промышленности Зоология - наука о животных. Подготовка к ОГЭ и ЕГЭ по биологии
Зоология - наука о животных. Подготовка к ОГЭ и ЕГЭ по биологии Основные фосфолипиды и гликолипиды тканей человека
Основные фосфолипиды и гликолипиды тканей человека