- Микробиологическая трансформация ксенобиотиков

Содержание
- 3. ОСНОВНЫЕ ИСТОЧНИКИ ЗАГРЯЗНЕНИЯ БИОСФЕРЫ химические и фармацевтические предприятия; предприятия целлюлозно-бумажной и печатной индустрии – основные потребители
- 4. САМООЧИЩЕНИЕ ОКРУЖАЮЩЕЙ СРЕДЫ Частичная нейтрализация последствий воздействия антропогенных факторов и самоочищение окружающей среды происходит благодаря мощному
- 5. Преимущества: высокая скорость и селективность ферментативных реакций; разнообразие ферментов, позволяющее микроорганизмам утилизировать широкий спектр субстратов и
- 6. Недостатки: При внедрении процесса биодеградации синтетического химического вещества необходимо исследовать всю цепочку его превращений, подобрать оптимальные
- 7. ПУТИ ОБЕЗВРЕЖИВАНИЯ КСЕНОБИОТИКОВ Минерализация (полная деструкция) происходит при использовании микроорганизмом ксенобиотика в качестве источника углерода и
- 8. БИОТРАНСФОРМАЦИЯ КСЕНОБИОТИКОВ 2 ступени: первичное биоразложение – трансформация микроорганизмами молекулы ксенобиотика с образованием соединений, не проявляющих
- 9. В зависимости от времени полураспада (t1/2) ксенобиотики делят на: легко доступные (t1/2 1–7 суток); умеренно доступные
- 10. БИОДОСТУПНОСТЬ НЕКОТОРЫХ КЛАССОВ КСЕНОБИОТИКОВ + - высокая; ± - умеренная; - - низкая
- 11. ФАКТОРЫ ВНЕШНЕЙ СРЕДЫ И БИОДОСТУПНОСТЬ КСЕНОБИОТИКОВ Биодоступность и активность биодеградации ксенобиотиков ограничивают следующие факторы: Неоптимальное значение
- 12. КОНЦЕНТРАЦИЯ КСЕНОБИОТИКА чем выше концентрация ксенобиотика, тем дольше он находится в окружающей среде; если концентрация ксенобиотика
- 13. НЕДОСТУПНОСТЬ АКЦЕПТОРОВ ЭЛЕКТРОНОВ ИЛИ КОСУБСТРАТОВ Деградация ксенобиотика микроорганизмами сопряжена с потреблением окислителя (акцептора электронов) или восстановителя
- 14. ТЕМПЕРАТУРА Для эффективных процессов биодеструкции температура – один из наиболее важных факторов окружающей среды. при повышении
- 15. АКТИВНАЯ КИСЛОТНОСТЬ оптимальная кислотность среды для большинства бактерий-деструкторов находится в интервале значений рН 6,0-8,0; эффект рН
- 16. ТОКСИЧЕСКОЕ ДЕЙСТВИЕ КСЕНОБИОТИКОВ НА МИКРООРГАНИЗМЫ Зависит от: природы ксенобиотика; концентрации ксенобиотика; продолжительности контакта с клеткой микроорганизма
- 17. МЕХАНИЗМЫ АДАПТАЦИИ МИКРООРГАНИЗМОВ К ТОКСИЧЕСКОМУ ДЕЙСТВИЮ КСЕНОБИОТИКОВ Механизмы адаптации: снижение скорости поступления ксенобиотика в клетку вследствие
- 18. ОСНОВНЫЕ СТАДИИ ВЗАИМОДЕЙСТВИЯ КСЕНОБИОТИКА С МИКРОБНОЙ КЛЕТКОЙ
- 19. 2. Транспорт ксенобиотика в клетку транспорт нейтральных и гидрофобных ксенобиотиков через мембрану осуществляется путем диффузии и
- 20. СТАДИИ ВЗАИМОДЕЙСТВИЯ КСЕНОБИОТИКА С МИКРОБНОЙ КЛЕТКОЙ 3. Первичная атака (периферийный метаболизм) - последовательная трансформация органического ксенобиотика
- 21. ПЕРИФЕРИЙНЫЙ МЕТАБОЛИЗМ Включает начальные этапы метаболизма ксенобиотиков Ферменты периферийного метаболизма: имеют широкую субстратную специфичность; являются индуцибельными
- 22. Включает консервативные метаболические пути, общие для большинства микроорганизмов: гликолиз; глюконеогенез; пентозофосфатный шунт; путь Энтнера-Дудорова; цикл Кребса;
- 23. МЕХАНИЗМЫ ДЕГРАДАЦИИ КСЕНОБИОТИКОВ Если синтетические ксенобиотики подобны по химическим свойствам природным соединениям, из трансформация проводится по
- 24. Бактерии, разрушающие негалогенированные ароматические соединения, превращают их в катехол или протокатехоат, а затем, в ходе нескольких
- 26. Пути ферментативного расщепления катехола и протокатехоата
- 27. Разрушение ароматических соединений содержащих сульфогруппу (эмульгаторы, смачиватели, компоненты красителей) протекает через предварительное удаление сульфогруппы, которая обеспечивает
- 28. Скорость и степень биодеградации нефтяных загрязнений определяется их компонентным составом. Легче всего разрушаются линейные алканы, хуже
- 29. Некоторые микроорганизмы обладают природной способностью к деградации различных ксенобиотиков, однако: ни один микроорганизм не может разрушать
- 30. «ПРИРОДНЫЕ» МИКРООРГАНИЗМЫ – ДЕСТРУКТОРЫ КСЕНОБИОТИКОВ Ведущая роль в биотрансформации органических ксенобиотиков принадлежит хемоорганотрофным (гетеротрофным) микроорганизмам, в
- 31. «ПРИРОДНЫЕ» МИКРООРГАНИЗМЫ – ДЕСТРУКТОРЫ КСЕНОБИОТИКОВ
- 32. «ПРИРОДНЫЕ» МИКРООРГАНИЗМЫ – ДЕСТРУКТОРЫ КСЕНОБИОТИКОВ
- 33. «ПРИРОДНЫЕ» МИКРООРГАНИЗМЫ – ДЕСТРУКТОРЫ КСЕНОБИОТИКОВ
- 34. СМЕШАННЫЕ ПОПУЛЯЦИИ МИКРООРГАНИЗМОВ – ДЕСТРУКТОРОВ КСЕНОБИОТИКОВ Смешанные популяции, как правило, быстрее и полнее разрушают органические ксенобиотики.
- 35. ПРИНЦИПЫ СЕЛЕКЦИИ МИКРООРГАНИЗМОВ – ДЕСТРУКТОРОВ КСЕНОБИОТИКОВ Выделять монокультуры или смешанные культуры микроорганизмов-деструкторов из сред, загрязненных конкретным
- 36. ПРИНЦИПЫ СЕЛЕКЦИИ МИКРООРГАНИЗМОВ – ДЕСТРУКТОРОВ КСЕНОБИОТИКОВ Ферменты подготовительного метаболизма микроорганизмов характеризуются низкой специфичностью, и метаболизм синтетических
- 37. ГЕННО-ИНЖЕНЕРНОЕ КОНСТРУИРОВАНИЕ МИКРООРГАНИЗМОВ – ДЕСТРУКТОРОВ КСЕНОБИОТИКОВ Конструирование рекомбинантных штаммов – деструкторов ксенобиотиков заключается в объединении нескольких
- 38. ПЛАЗМИДЫ ДЕГРАДАЦИИ (D-ПЛАЗМИДЫ) содержат гены, детерминирующие утилизацию ксенобиотиков; наиболее распространены у грамотрицательных бактерий.
- 40. В 1970-х гг. Чакрабарти и коллегами (США) был создан первый бактериальный штамм с широкими катаболическими возможностями,
- 42. ПРОБЛЕМЫ ПРИ ИСПОЛЬЗОВАНИИ ГЕННО-ИНЖЕНЕРНЫХ МИКРООРГАНИЗМОВ – ДЕСТРУКТОРОВ КСЕНОБИОТИКОВ генетическая нестабильность; низкая конкурентоспособность по сравнению с автохтонной
- 44. Скачать презентацию


ОСНОВНЫЕ ИСТОЧНИКИ ЗАГРЯЗНЕНИЯ БИОСФЕРЫ
химические и фармацевтические предприятия;
предприятия целлюлозно-бумажной и
ОСНОВНЫЕ ИСТОЧНИКИ ЗАГРЯЗНЕНИЯ БИОСФЕРЫ
химические и фармацевтические предприятия;
предприятия целлюлозно-бумажной и

САМООЧИЩЕНИЕ ОКРУЖАЮЩЕЙ СРЕДЫ
Частичная нейтрализация последствий воздействия антропогенных факторов и самоочищение
САМООЧИЩЕНИЕ ОКРУЖАЮЩЕЙ СРЕДЫ
Частичная нейтрализация последствий воздействия антропогенных факторов и самоочищение

Преимущества:
высокая скорость и селективность ферментативных реакций;
разнообразие ферментов, позволяющее микроорганизмам
Преимущества:
высокая скорость и селективность ферментативных реакций;
разнообразие ферментов, позволяющее микроорганизмам

Недостатки:
При внедрении процесса биодеградации синтетического химического вещества необходимо исследовать всю цепочку
Недостатки:
При внедрении процесса биодеградации синтетического химического вещества необходимо исследовать всю цепочку

ПУТИ ОБЕЗВРЕЖИВАНИЯ КСЕНОБИОТИКОВ
Минерализация (полная деструкция)
происходит при использовании микроорганизмом ксенобиотика в качестве
ПУТИ ОБЕЗВРЕЖИВАНИЯ КСЕНОБИОТИКОВ
Минерализация (полная деструкция)
происходит при использовании микроорганизмом ксенобиотика в качестве

БИОТРАНСФОРМАЦИЯ КСЕНОБИОТИКОВ
2 ступени:
первичное биоразложение – трансформация микроорганизмами молекулы ксенобиотика с образованием
БИОТРАНСФОРМАЦИЯ КСЕНОБИОТИКОВ
2 ступени:
первичное биоразложение – трансформация микроорганизмами молекулы ксенобиотика с образованием

В зависимости от времени полураспада (t1/2) ксенобиотики делят на:
легко доступные
В зависимости от времени полураспада (t1/2) ксенобиотики делят на:
легко доступные
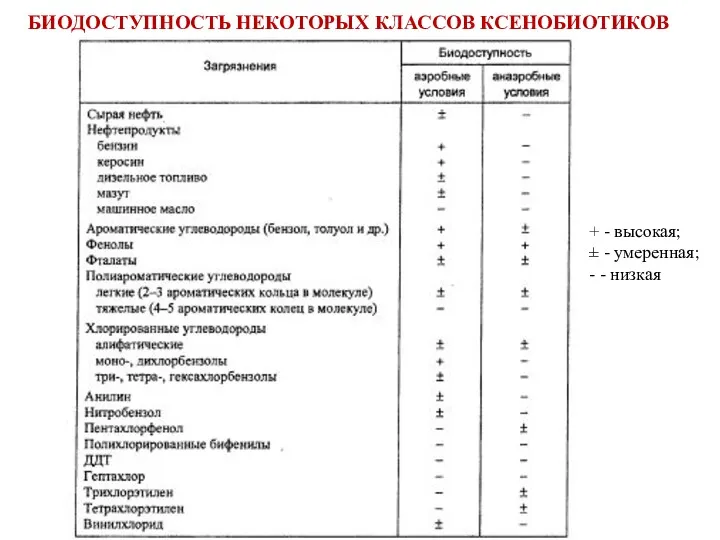
БИОДОСТУПНОСТЬ НЕКОТОРЫХ КЛАССОВ КСЕНОБИОТИКОВ
+ - высокая;
± - умеренная;
- - низкая
БИОДОСТУПНОСТЬ НЕКОТОРЫХ КЛАССОВ КСЕНОБИОТИКОВ
+ - высокая;
± - умеренная;
- - низкая

ФАКТОРЫ ВНЕШНЕЙ СРЕДЫ
И БИОДОСТУПНОСТЬ КСЕНОБИОТИКОВ
Биодоступность и активность биодеградации ксенобиотиков ограничивают
ФАКТОРЫ ВНЕШНЕЙ СРЕДЫ
И БИОДОСТУПНОСТЬ КСЕНОБИОТИКОВ
Биодоступность и активность биодеградации ксенобиотиков ограничивают

КОНЦЕНТРАЦИЯ КСЕНОБИОТИКА
чем выше концентрация ксенобиотика, тем дольше он находится в
КОНЦЕНТРАЦИЯ КСЕНОБИОТИКА
чем выше концентрация ксенобиотика, тем дольше он находится в

НЕДОСТУПНОСТЬ АКЦЕПТОРОВ ЭЛЕКТРОНОВ
ИЛИ КОСУБСТРАТОВ
Деградация ксенобиотика микроорганизмами сопряжена с потреблением окислителя
НЕДОСТУПНОСТЬ АКЦЕПТОРОВ ЭЛЕКТРОНОВ
ИЛИ КОСУБСТРАТОВ
Деградация ксенобиотика микроорганизмами сопряжена с потреблением окислителя

ТЕМПЕРАТУРА
Для эффективных процессов биодеструкции температура – один из наиболее важных факторов
ТЕМПЕРАТУРА
Для эффективных процессов биодеструкции температура – один из наиболее важных факторов

АКТИВНАЯ КИСЛОТНОСТЬ
оптимальная кислотность среды для большинства бактерий-деструкторов находится в интервале значений
АКТИВНАЯ КИСЛОТНОСТЬ
оптимальная кислотность среды для большинства бактерий-деструкторов находится в интервале значений

ТОКСИЧЕСКОЕ ДЕЙСТВИЕ КСЕНОБИОТИКОВ
НА МИКРООРГАНИЗМЫ
Зависит от:
природы ксенобиотика;
концентрации ксенобиотика;
продолжительности контакта с клеткой
ТОКСИЧЕСКОЕ ДЕЙСТВИЕ КСЕНОБИОТИКОВ
НА МИКРООРГАНИЗМЫ
Зависит от:
природы ксенобиотика;
концентрации ксенобиотика;
продолжительности контакта с клеткой

МЕХАНИЗМЫ АДАПТАЦИИ МИКРООРГАНИЗМОВ
К ТОКСИЧЕСКОМУ ДЕЙСТВИЮ КСЕНОБИОТИКОВ
Механизмы адаптации:
снижение скорости поступления ксенобиотика
МЕХАНИЗМЫ АДАПТАЦИИ МИКРООРГАНИЗМОВ
К ТОКСИЧЕСКОМУ ДЕЙСТВИЮ КСЕНОБИОТИКОВ
Механизмы адаптации:
снижение скорости поступления ксенобиотика
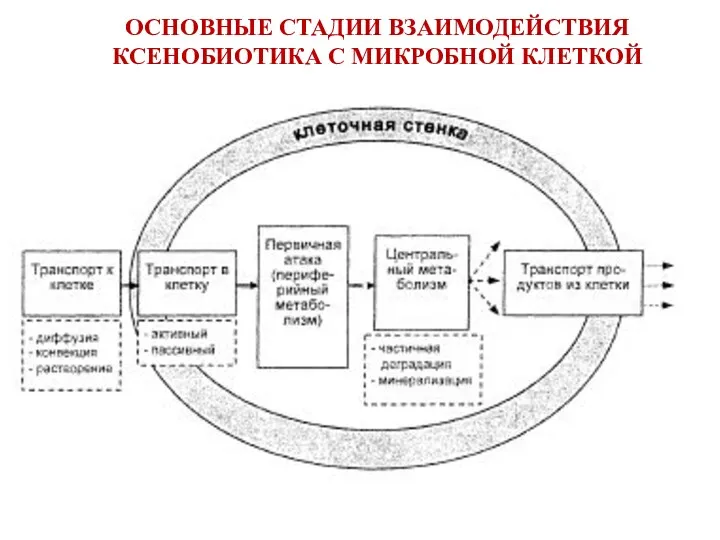
ОСНОВНЫЕ СТАДИИ ВЗАИМОДЕЙСТВИЯ КСЕНОБИОТИКА С МИКРОБНОЙ КЛЕТКОЙ
ОСНОВНЫЕ СТАДИИ ВЗАИМОДЕЙСТВИЯ КСЕНОБИОТИКА С МИКРОБНОЙ КЛЕТКОЙ

2. Транспорт ксенобиотика в клетку
транспорт нейтральных и гидрофобных ксенобиотиков через
2. Транспорт ксенобиотика в клетку
транспорт нейтральных и гидрофобных ксенобиотиков через

СТАДИИ ВЗАИМОДЕЙСТВИЯ КСЕНОБИОТИКА С МИКРОБНОЙ КЛЕТКОЙ
3. Первичная атака (периферийный метаболизм) -
СТАДИИ ВЗАИМОДЕЙСТВИЯ КСЕНОБИОТИКА С МИКРОБНОЙ КЛЕТКОЙ
3. Первичная атака (периферийный метаболизм) -

ПЕРИФЕРИЙНЫЙ МЕТАБОЛИЗМ
Включает начальные этапы метаболизма ксенобиотиков
Ферменты периферийного метаболизма:
имеют
ПЕРИФЕРИЙНЫЙ МЕТАБОЛИЗМ
Включает начальные этапы метаболизма ксенобиотиков
Ферменты периферийного метаболизма:
имеют
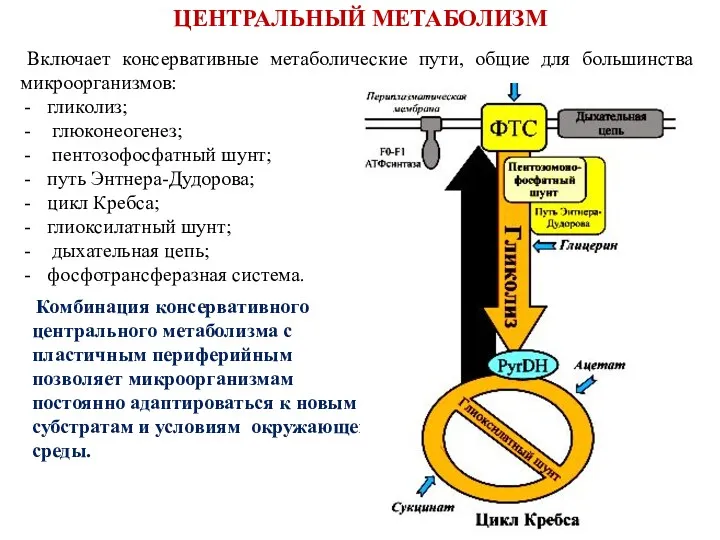
Включает консервативные метаболические пути, общие для большинства микроорганизмов:
гликолиз;
глюконеогенез;
пентозофосфатный
Включает консервативные метаболические пути, общие для большинства микроорганизмов:
гликолиз;
глюконеогенез;
пентозофосфатный

МЕХАНИЗМЫ ДЕГРАДАЦИИ КСЕНОБИОТИКОВ
Если синтетические ксенобиотики подобны по химическим свойствам природным
МЕХАНИЗМЫ ДЕГРАДАЦИИ КСЕНОБИОТИКОВ
Если синтетические ксенобиотики подобны по химическим свойствам природным

Бактерии, разрушающие негалогенированные ароматические соединения, превращают их в катехол или протокатехоат,
Бактерии, разрушающие негалогенированные ароматические соединения, превращают их в катехол или протокатехоат,

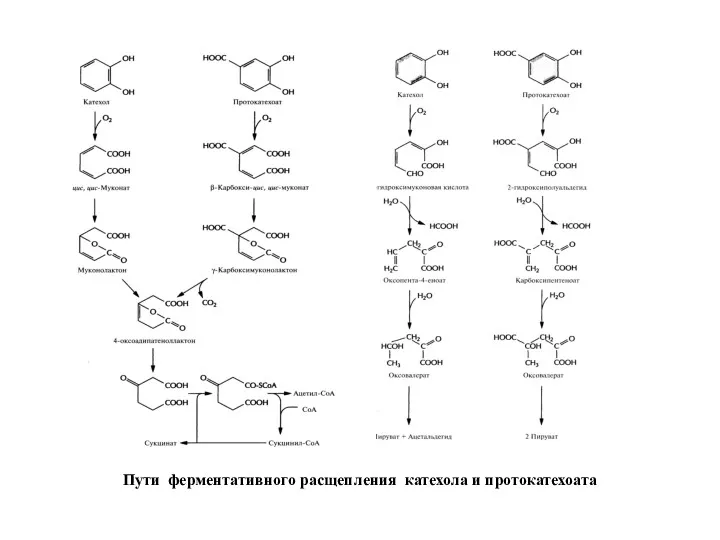
Пути ферментативного расщепления катехола и протокатехоата
Пути ферментативного расщепления катехола и протокатехоата
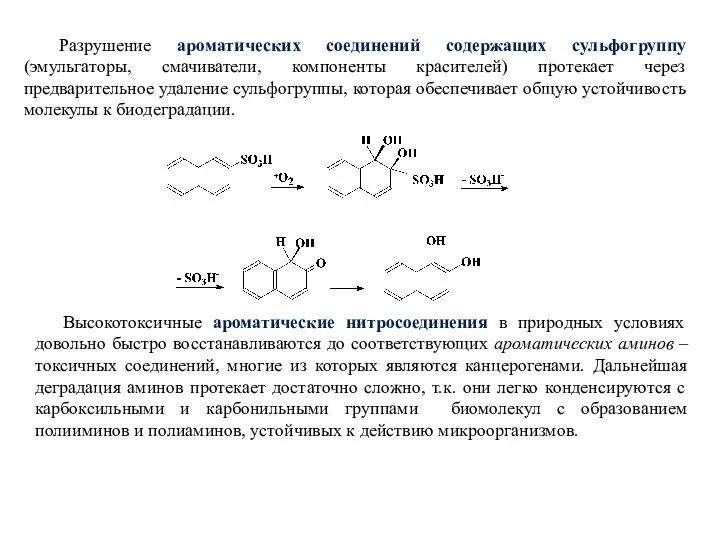
Разрушение ароматических соединений содержащих сульфогруппу (эмульгаторы, смачиватели, компоненты красителей) протекает через
Разрушение ароматических соединений содержащих сульфогруппу (эмульгаторы, смачиватели, компоненты красителей) протекает через

Скорость и степень биодеградации нефтяных загрязнений определяется их компонентным составом. Легче
Скорость и степень биодеградации нефтяных загрязнений определяется их компонентным составом. Легче

Некоторые микроорганизмы обладают природной способностью к деградации различных ксенобиотиков, однако:
ни
Некоторые микроорганизмы обладают природной способностью к деградации различных ксенобиотиков, однако:
ни

«ПРИРОДНЫЕ» МИКРООРГАНИЗМЫ –
ДЕСТРУКТОРЫ КСЕНОБИОТИКОВ
Ведущая роль в биотрансформации органических ксенобиотиков принадлежит
«ПРИРОДНЫЕ» МИКРООРГАНИЗМЫ –
ДЕСТРУКТОРЫ КСЕНОБИОТИКОВ
Ведущая роль в биотрансформации органических ксенобиотиков принадлежит

«ПРИРОДНЫЕ» МИКРООРГАНИЗМЫ –
ДЕСТРУКТОРЫ КСЕНОБИОТИКОВ
«ПРИРОДНЫЕ» МИКРООРГАНИЗМЫ –
ДЕСТРУКТОРЫ КСЕНОБИОТИКОВ

«ПРИРОДНЫЕ» МИКРООРГАНИЗМЫ –
ДЕСТРУКТОРЫ КСЕНОБИОТИКОВ
«ПРИРОДНЫЕ» МИКРООРГАНИЗМЫ –
ДЕСТРУКТОРЫ КСЕНОБИОТИКОВ

«ПРИРОДНЫЕ» МИКРООРГАНИЗМЫ –
ДЕСТРУКТОРЫ КСЕНОБИОТИКОВ
«ПРИРОДНЫЕ» МИКРООРГАНИЗМЫ –
ДЕСТРУКТОРЫ КСЕНОБИОТИКОВ
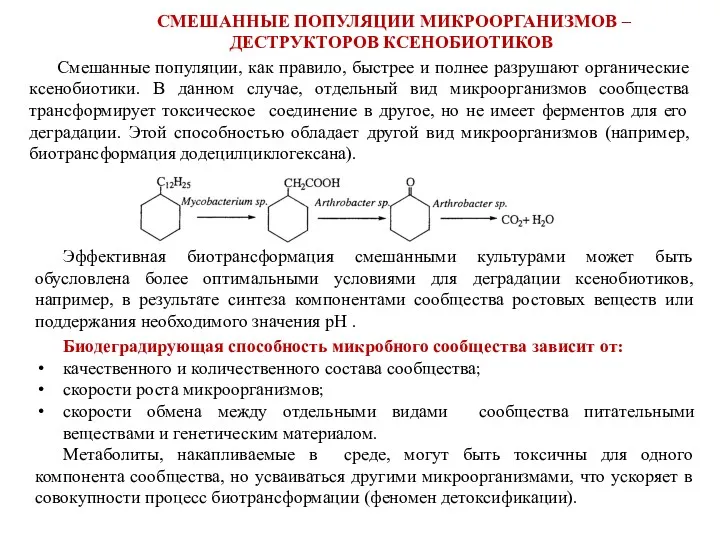
СМЕШАННЫЕ ПОПУЛЯЦИИ МИКРООРГАНИЗМОВ –
ДЕСТРУКТОРОВ КСЕНОБИОТИКОВ
Смешанные популяции, как правило, быстрее и
СМЕШАННЫЕ ПОПУЛЯЦИИ МИКРООРГАНИЗМОВ –
ДЕСТРУКТОРОВ КСЕНОБИОТИКОВ
Смешанные популяции, как правило, быстрее и

ПРИНЦИПЫ СЕЛЕКЦИИ МИКРООРГАНИЗМОВ –
ДЕСТРУКТОРОВ КСЕНОБИОТИКОВ
Выделять монокультуры или смешанные культуры микроорганизмов-деструкторов
ПРИНЦИПЫ СЕЛЕКЦИИ МИКРООРГАНИЗМОВ –
ДЕСТРУКТОРОВ КСЕНОБИОТИКОВ
Выделять монокультуры или смешанные культуры микроорганизмов-деструкторов

ПРИНЦИПЫ СЕЛЕКЦИИ МИКРООРГАНИЗМОВ –
ДЕСТРУКТОРОВ КСЕНОБИОТИКОВ
Ферменты подготовительного метаболизма микроорганизмов характеризуются низкой
ПРИНЦИПЫ СЕЛЕКЦИИ МИКРООРГАНИЗМОВ –
ДЕСТРУКТОРОВ КСЕНОБИОТИКОВ
Ферменты подготовительного метаболизма микроорганизмов характеризуются низкой

ГЕННО-ИНЖЕНЕРНОЕ КОНСТРУИРОВАНИЕ МИКРООРГАНИЗМОВ – ДЕСТРУКТОРОВ КСЕНОБИОТИКОВ
Конструирование рекомбинантных штаммов – деструкторов ксенобиотиков
ГЕННО-ИНЖЕНЕРНОЕ КОНСТРУИРОВАНИЕ МИКРООРГАНИЗМОВ – ДЕСТРУКТОРОВ КСЕНОБИОТИКОВ
Конструирование рекомбинантных штаммов – деструкторов ксенобиотиков
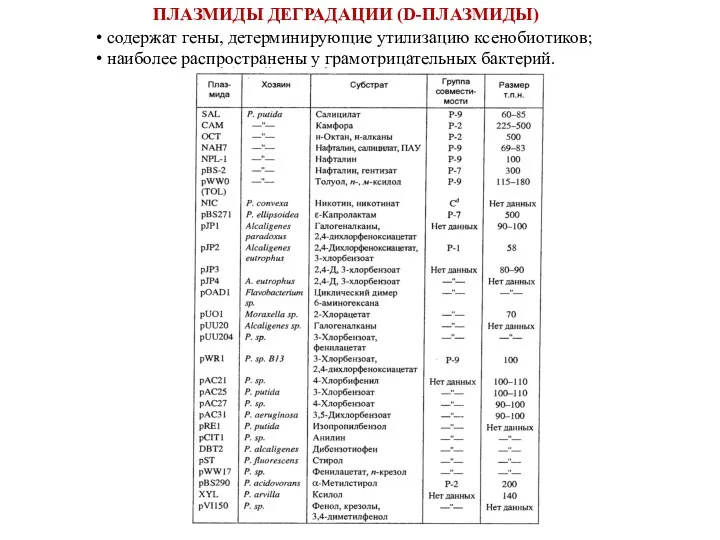
ПЛАЗМИДЫ ДЕГРАДАЦИИ (D-ПЛАЗМИДЫ)
содержат гены, детерминирующие утилизацию ксенобиотиков;
наиболее распространены у
ПЛАЗМИДЫ ДЕГРАДАЦИИ (D-ПЛАЗМИДЫ)
содержат гены, детерминирующие утилизацию ксенобиотиков;
наиболее распространены у
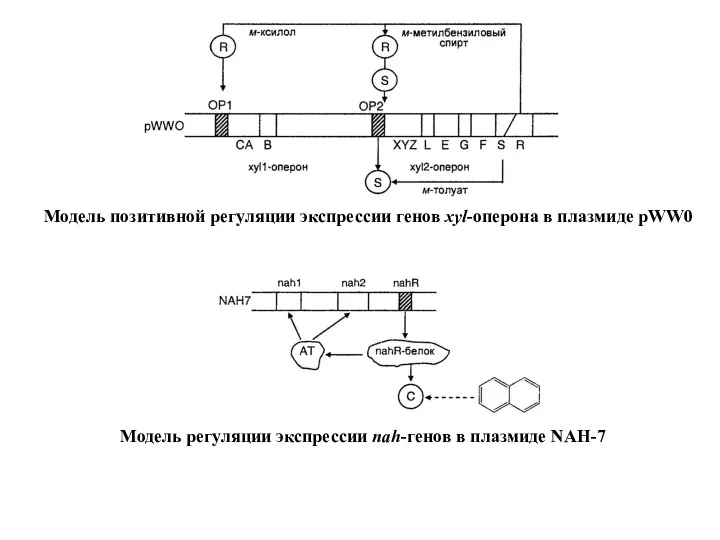

В 1970-х гг. Чакрабарти и коллегами (США) был создан первый бактериальный
В 1970-х гг. Чакрабарти и коллегами (США) был создан первый бактериальный


ПРОБЛЕМЫ ПРИ ИСПОЛЬЗОВАНИИ ГЕННО-ИНЖЕНЕРНЫХ МИКРООРГАНИЗМОВ – ДЕСТРУКТОРОВ КСЕНОБИОТИКОВ
генетическая нестабильность;
низкая
ПРОБЛЕМЫ ПРИ ИСПОЛЬЗОВАНИИ ГЕННО-ИНЖЕНЕРНЫХ МИКРООРГАНИЗМОВ – ДЕСТРУКТОРОВ КСЕНОБИОТИКОВ
генетическая нестабильность;
низкая
 От микроскопа к клетке.
От микроскопа к клетке. Подцарство Одноклеточные. Простейшие
Подцарство Одноклеточные. Простейшие Ген инженериясының мәселелері мен міндеттері
Ген инженериясының мәселелері мен міндеттері Деревья. Строение дерева
Деревья. Строение дерева Интеллектуально - развлекательная игра В здоровом теле здоровый дух
Интеллектуально - развлекательная игра В здоровом теле здоровый дух Удивительные факты из жизни рыб
Удивительные факты из жизни рыб Отдел голосеменные. ОГЭ
Отдел голосеменные. ОГЭ Отряд Непарнокопытные (Perissodactyla)
Отряд Непарнокопытные (Perissodactyla) Онтогенез нервной системы
Онтогенез нервной системы Происхождение человека
Происхождение человека Ферменты. Свойства. Функции. Механизм действия
Ферменты. Свойства. Функции. Механизм действия Жануарлардың жүйке жүйелерінің типтері. Жүйке жүйесінің қызметі мен оның құрылымдық компонентерінің атау
Жануарлардың жүйке жүйелерінің типтері. Жүйке жүйесінің қызметі мен оның құрылымдық компонентерінің атау Микориза (грибокорень)
Микориза (грибокорень) Вирусы – неклеточная форма жизни…
Вирусы – неклеточная форма жизни… Професія дощового черв'яка
Професія дощового черв'яка Утконос. Особенности и среда обитания
Утконос. Особенности и среда обитания Sequoia sempervirens
Sequoia sempervirens Формирование культуры здорового питания школьников на уроках биологии.
Формирование культуры здорового питания школьников на уроках биологии. Кожа рыб и ее производные
Кожа рыб и ее производные Презентация Пресмыкающиеся. Кроссворд
Презентация Пресмыкающиеся. Кроссворд Викторина по теме Биотические связи в природе
Викторина по теме Биотические связи в природе Генетичні основи селекції. Тема 8
Генетичні основи селекції. Тема 8 Урок-презентация В бобовом царстве, мотыльковом государстве
Урок-презентация В бобовом царстве, мотыльковом государстве Размножение и оплодотворение растений
Размножение и оплодотворение растений Периодизация и продолжительность жизни у животных
Периодизация и продолжительность жизни у животных Декоративные кролики
Декоративные кролики Родючість грунту
Родючість грунту Биологические системы. Строение организмов. Основные процессы, происходящие в организме
Биологические системы. Строение организмов. Основные процессы, происходящие в организме