- Модели биологических аналогий

Содержание
- 2. Средства и методы моделирования Методы моделирования – методы динамической теории систем Средства – дифференциальные и разностные
- 3. Пример. Рост колонии микроорганизмов За время прирост численности равен , где R – число родившихся и
- 4. Пример. Рост колонии микроорганизмов В простейшем случае, когда рождаемость и смертность пропорциональны численности: или Получим экспоненциальную
- 5. Пример. Рост колонии микроорганизмов График функции при положительных (размножение) и отрицательных (вымирание) значениях константы скорости роста
- 6. Уравнение Ферхюльста Логистическое уравнение было предложено Ферхюльстом в 1838 г. При малых х численность x возрастает,
- 7. Динамика численности в логистической модели при разных начальных значениях численности популяции Если начальное значение , кривая
- 8. Системы двух автономных дифференциальных уравнений Рассмотрим систему двух автономных дифф. Уравнений P(x, y), Q(x, y) -
- 9. Фазовая плоскость Плоскость всех точек M(x,y) называется фазовой плоскости и изображает совокупность всех состояний системы. Точка
- 10. Фазовый портрет Совокупность фазовых траекторий при различных начальных значениях переменных дает фазовый портрет системы Для изображения
- 11. Направление вектора dy/dx
- 12. Фазовые траекторий в аналитическом виде дает семейство интегральных кривых уравнения - фазовых траекторий системы на плоскости
- 13. Линейные системы Рассмотрим систему a, b, c, d = const
- 14. Вид фазовых траекторий
- 15. Математические модели взаимодействия двух видов. Гипотезы Вольтерра 1. Пища либо имеется в неограниченном количестве, либо ее
- 16. Математические модели взаимодействия двух видов. Гипотезы Вольтерра 4. Если имеется пища в ограниченном количестве и несколько
- 17. Уравнения Вольтерра x1, x2 – численности видов ai – константы собственной скорости роста видов, ci –
- 18. Основные типы взаимодействий
- 19. Уравнения конкуренции
- 20. Стационарные решения
- 21. Условие сосуществования видов Необходимое условие устойчивости
- 22. Расположение главных изоклин на фазовом портрете Выживание x1 Выживание x2 Сосуществование видов Триггерная система
- 23. Система «хищник-жертва»
- 24. Стационарные решения системы
- 25. Расположение главных изоклин на фазовом портрете Устойчивое сосуществование Популяция хищников вымерла Точки 1 и 3 неустойчивые
- 26. Моделирование динамики популяций с помощью уравнений Лотка-Вольтерра Модель взаимодействия хищников и их добычи, когда между особями
- 27. Решение системы Лотка-Вольтерра
- 28. Уравнения Лотка-Вольтерра с логистической поправкой Модель конкурирующих видов с логистической поправкой
- 29. Графики решений и фазовая кривая, α>0
- 30. Графики решений и фазовая кривая, α
- 31. Модель Холлинга-Тэннера Скорость роста популяции жертв dx1⁄dt в этой модели равна сумме трех величин
- 32. Модель Холлинга-Тэннера Скорость роста популяции хищников dx2⁄dt строится так же, как в модели Вольтерра–Лотка, в предположении,
- 33. Модель Холлинга-Тэннера При на фазовом портрете системы будет устойчивый предельный цикл
- 34. Графики решений и фазовый портрет при
- 35. Графики решений и фазовый портрет при
- 36. Обобщенная модель взаимодействия двух видов А. Н. Колмогорова
- 37. Обобщенная модель взаимодействия двух видов А. Н. Колмогорова 1) Хищники не взаимодействуют друг с другом, т.е.
- 38. Обобщенная модель взаимодействия двух видов А. Н. Колмогорова 4) dk1/dx 5) dk2/dx > 0, k1(0)
- 40. Скачать презентацию

Средства и методы моделирования
Методы моделирования – методы динамической теории систем
Средства –
Средства и методы моделирования
Методы моделирования – методы динамической теории систем
Средства –

Пример. Рост колонии микроорганизмов
За время прирост численности равен ,
где R –
Пример. Рост колонии микроорганизмов
За время прирост численности равен ,
где R –

Пример. Рост колонии микроорганизмов
В простейшем случае, когда рождаемость и смертность пропорциональны
Пример. Рост колонии микроорганизмов
В простейшем случае, когда рождаемость и смертность пропорциональны
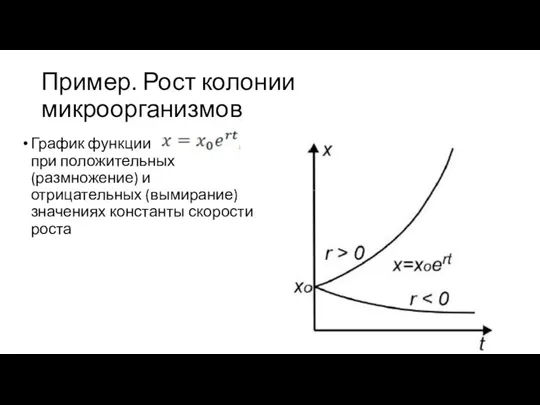
Пример. Рост колонии микроорганизмов
График функции при положительных (размножение) и отрицательных (вымирание)
Пример. Рост колонии микроорганизмов
График функции при положительных (размножение) и отрицательных (вымирание)

Уравнение Ферхюльста
Логистическое уравнение было предложено Ферхюльстом в 1838 г.
При малых х
Уравнение Ферхюльста
Логистическое уравнение было предложено Ферхюльстом в 1838 г.
При малых х
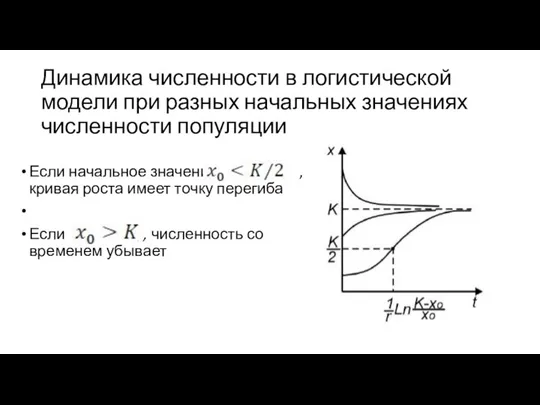
Динамика численности в логистической модели при разных начальных значениях численности популяции
Если
Динамика численности в логистической модели при разных начальных значениях численности популяции
Если

Системы двух автономных
дифференциальных уравнений
Рассмотрим систему двух автономных дифф. Уравнений
P(x,
Системы двух автономных
дифференциальных уравнений
Рассмотрим систему двух автономных дифф. Уравнений
P(x,

Фазовая плоскость
Плоскость всех точек M(x,y) называется фазовой плоскости и изображает совокупность
Фазовая плоскость
Плоскость всех точек M(x,y) называется фазовой плоскости и изображает совокупность

Фазовый портрет
Совокупность фазовых траекторий при различных начальных значениях переменных дает фазовый
Фазовый портрет
Совокупность фазовых траекторий при различных начальных значениях переменных дает фазовый

Направление вектора dy/dx
Направление вектора dy/dx

Фазовые траекторий в аналитическом виде
дает семейство интегральных кривых уравнения - фазовых
Фазовые траекторий в аналитическом виде
дает семейство интегральных кривых уравнения - фазовых

Линейные системы
Рассмотрим систему
a, b, c, d = const
Линейные системы
Рассмотрим систему
a, b, c, d = const
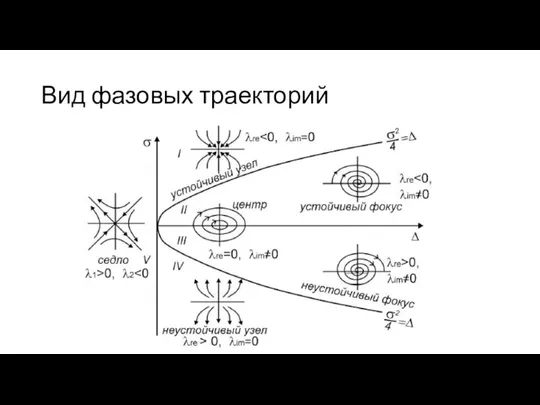
Вид фазовых траекторий
Вид фазовых траекторий

Математические модели взаимодействия двух видов. Гипотезы Вольтерра
1. Пища либо имеется в
Математические модели взаимодействия двух видов. Гипотезы Вольтерра
1. Пища либо имеется в

Математические модели взаимодействия двух видов. Гипотезы Вольтерра
4. Если имеется пища в
Математические модели взаимодействия двух видов. Гипотезы Вольтерра
4. Если имеется пища в

Уравнения Вольтерра
x1, x2 – численности видов
ai – константы собственной скорости роста
Уравнения Вольтерра
x1, x2 – численности видов
ai – константы собственной скорости роста

Основные типы взаимодействий
Основные типы взаимодействий

Уравнения конкуренции
Уравнения конкуренции

Стационарные решения
Стационарные решения

Условие сосуществования видов
Необходимое условие устойчивости
Условие сосуществования видов
Необходимое условие устойчивости

Расположение главных изоклин на фазовом портрете
Выживание x1
Выживание x2
Сосуществование видов
Триггерная система
Расположение главных изоклин на фазовом портрете
Выживание x1
Выживание x2
Сосуществование видов
Триггерная система

Система «хищник-жертва»
Система «хищник-жертва»

Стационарные решения системы
Стационарные решения системы

Расположение главных изоклин на фазовом портрете
Устойчивое сосуществование
Популяция хищников вымерла
Точки 1
Расположение главных изоклин на фазовом портрете
Устойчивое сосуществование
Популяция хищников вымерла
Точки 1

Моделирование динамики популяций с помощью уравнений Лотка-Вольтерра
Модель взаимодействия хищников и их
Моделирование динамики популяций с помощью уравнений Лотка-Вольтерра
Модель взаимодействия хищников и их
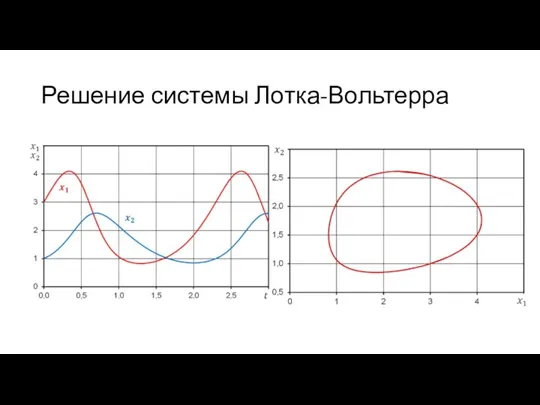
Решение системы Лотка-Вольтерра
Решение системы Лотка-Вольтерра

Уравнения Лотка-Вольтерра с логистической поправкой
Модель конкурирующих видов с логистической поправкой
Уравнения Лотка-Вольтерра с логистической поправкой
Модель конкурирующих видов с логистической поправкой
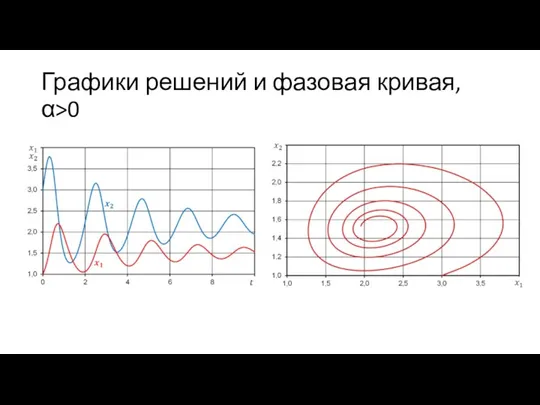
Графики решений и фазовая кривая, α>0
Графики решений и фазовая кривая, α>0
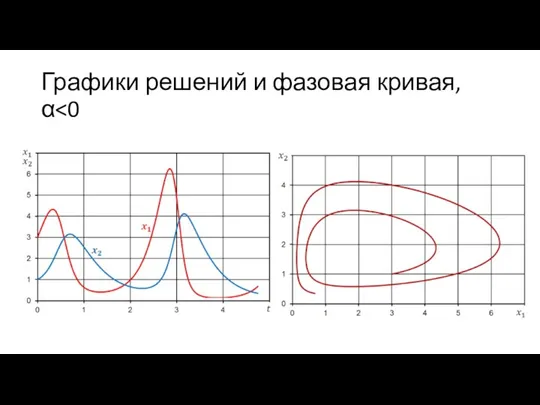
Графики решений и фазовая кривая, α<0
Графики решений и фазовая кривая, α<0

Модель Холлинга-Тэннера
Скорость роста популяции жертв dx1⁄dt в этой модели равна сумме
Модель Холлинга-Тэннера
Скорость роста популяции жертв dx1⁄dt в этой модели равна сумме

Модель Холлинга-Тэннера
Скорость роста популяции хищников dx2⁄dt строится так же, как в
Модель Холлинга-Тэннера
Скорость роста популяции хищников dx2⁄dt строится так же, как в

Модель Холлинга-Тэннера
При на фазовом портрете системы будет устойчивый предельный цикл
Модель Холлинга-Тэннера
При на фазовом портрете системы будет устойчивый предельный цикл
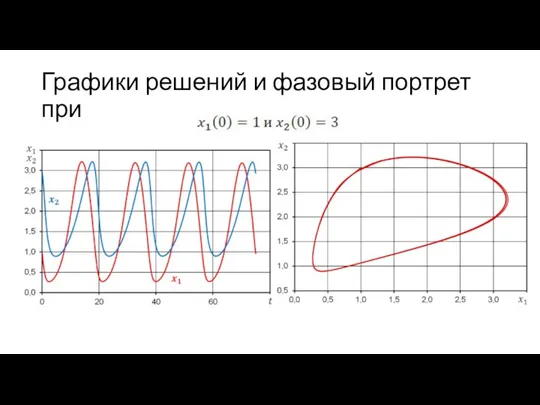
Графики решений и фазовый портрет при
Графики решений и фазовый портрет при
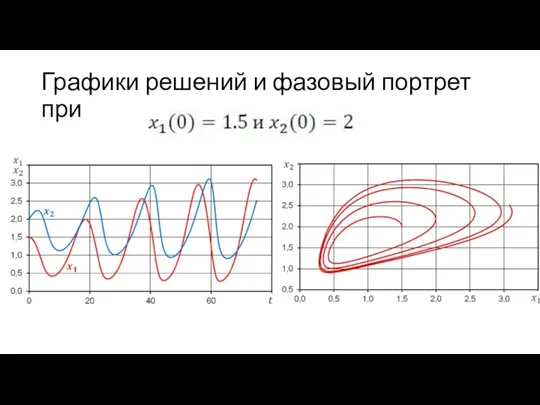
Графики решений и фазовый портрет при
Графики решений и фазовый портрет при

Обобщенная модель взаимодействия двух видов А. Н. Колмогорова
Обобщенная модель взаимодействия двух видов А. Н. Колмогорова

Обобщенная модель взаимодействия двух видов А. Н. Колмогорова
1) Хищники не взаимодействуют
Обобщенная модель взаимодействия двух видов А. Н. Колмогорова
1) Хищники не взаимодействуют

Обобщенная модель взаимодействия двух видов А. Н. Колмогорова
4) dk1/dx < 0.
Обобщенная модель взаимодействия двух видов А. Н. Колмогорова
4) dk1/dx < 0.
 Генетика пола. Наследование, сцепленное с полом
Генетика пола. Наследование, сцепленное с полом урок биологии в 7 классе Внешнее и внутреннее строение насекомых
урок биологии в 7 классе Внешнее и внутреннее строение насекомых Породы кошек
Породы кошек Происхождение человека. Расы и расогенез. Адаптивные типы людей
Происхождение человека. Расы и расогенез. Адаптивные типы людей Цветковые (покрытосемянные) растения
Цветковые (покрытосемянные) растения Многообразие организмов. Вирусы
Многообразие организмов. Вирусы Ядовитые грибы
Ядовитые грибы Птицы Ненецкого Автономного округа
Птицы Ненецкого Автономного округа Структура и динамика популяций. Внутривидовые и межвидовые взаимоотношения в популяциях, гомеостаз и экологические стратегии
Структура и динамика популяций. Внутривидовые и межвидовые взаимоотношения в популяциях, гомеостаз и экологические стратегии Психология и социобиология половых различий
Психология и социобиология половых различий Органы чувств. Строение и функции аппаратов глаза. Строение и гистофизиология органа обоняния
Органы чувств. Строение и функции аппаратов глаза. Строение и гистофизиология органа обоняния Факты о генетике
Факты о генетике Разнообразие зеленых водорослей
Разнообразие зеленых водорослей Животный мир Мещёры
Животный мир Мещёры Анатомия и морфология рыб
Анатомия и морфология рыб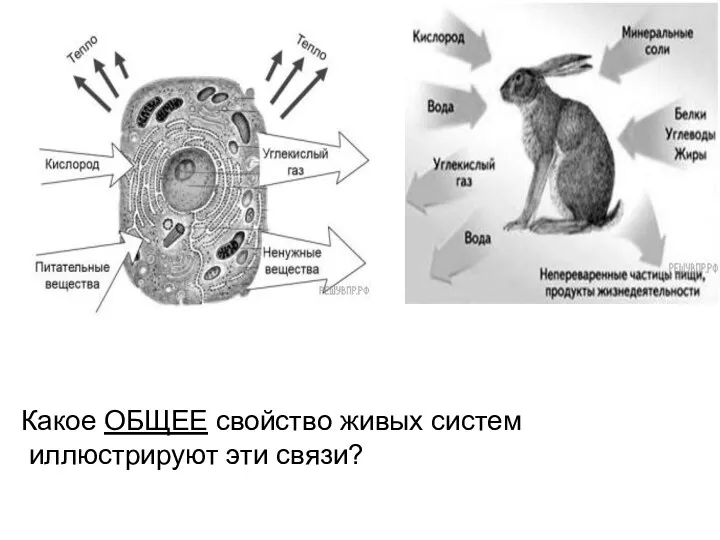 Обмен веществ и энергии
Обмен веществ и энергии Проблемы Антропогенеза
Проблемы Антропогенеза Мхи, хвощи, папоротники, плауны
Мхи, хвощи, папоротники, плауны Физиология гладких мышц. (Лекция 7)
Физиология гладких мышц. (Лекция 7) Морфология сельскохозяйственных животных. Аппарат движения. Остеология, миология
Морфология сельскохозяйственных животных. Аппарат движения. Остеология, миология Высшая нервная деятельность
Высшая нервная деятельность Презентация Лист. внешнее и внутреннее строение.
Презентация Лист. внешнее и внутреннее строение. Промежуточный мозг
Промежуточный мозг Углеводы. Классификация углеводов
Углеводы. Классификация углеводов Эмбриогенез печени и желчевыводящих путей
Эмбриогенез печени и желчевыводящих путей Транскрипция у эукариот
Транскрипция у эукариот Первая и вторая сигнальные системы человека
Первая и вторая сигнальные системы человека Презентация к внеклассному мероприятию Муравьи- общественные насекомые
Презентация к внеклассному мероприятию Муравьи- общественные насекомые