- Молекулярні основи стійкості рослин

Содержание
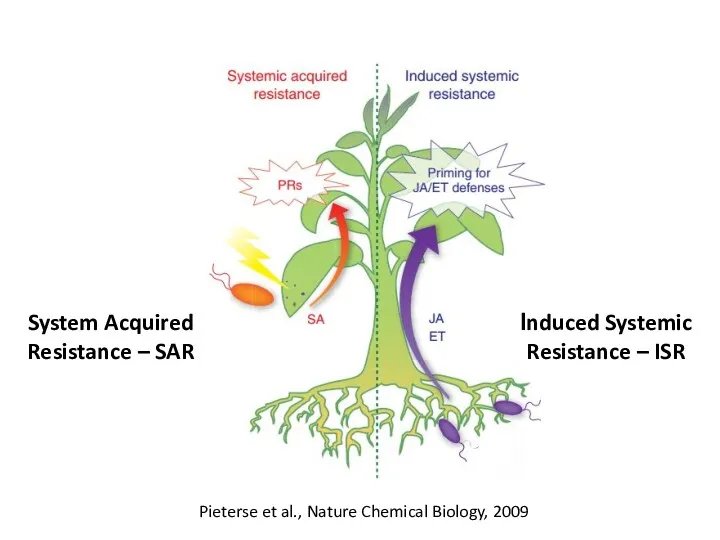
- 2. ІНДУКОВАНА (НЕСПЕЦИФІЧНА НАБУТА) СТІЙКІСТЬ РОСЛИН System Acquired Resistance – SAR + Іnduced Systemic Resistance – ISR
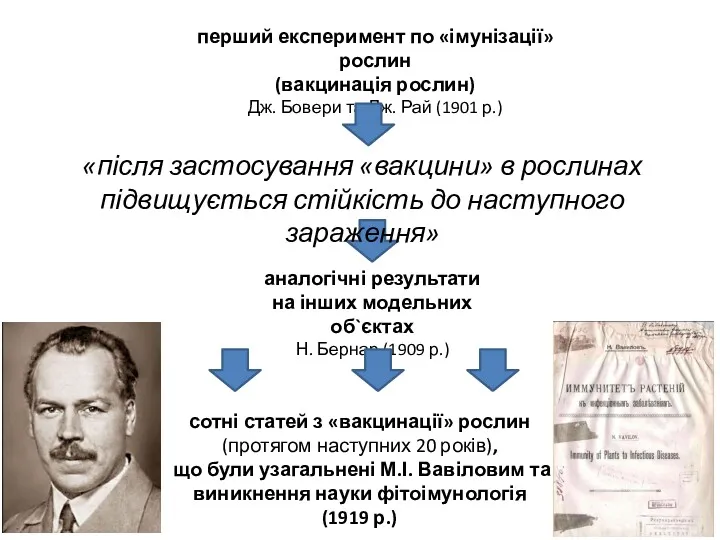
- 3. перший експеримент по «імунізації» рослин (вакцинація рослин) Дж. Бовери та Дж. Рай (1901 р.) аналогічні результати
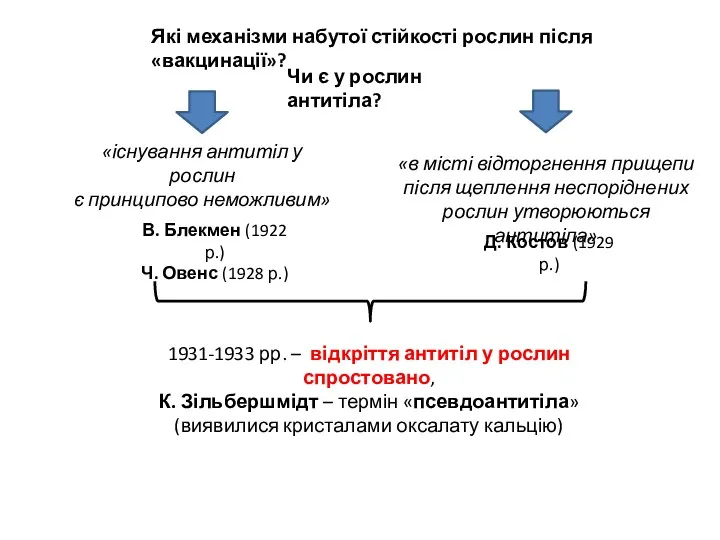
- 4. Які механізми набутої стійкості рослин після «вакцинації»? Чи є у рослин антитіла? В. Блекмен (1922 р.)
- 5. Станом на 1930-і рр. було встановлено, що стійкість рослин, що виникає після впливу «вакцини»: КОРОТКОСТРОКОВА (стійкий
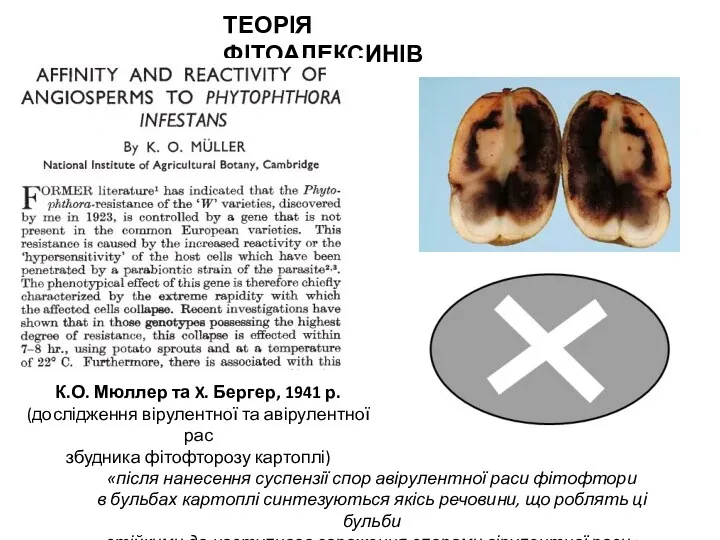
- 6. ТЕОРІЯ ФІТОАЛЕКСИНІВ К.О. Мюллер та X. Бергер, 1941 р. (дослідження вірулентної та авірулентної рас збудника фітофторозу
- 7. Фітоалексини відсутні у здоровій рослині і синтезуються (de novo) у відповідь на зараження Фітоалексини – неспецифічні
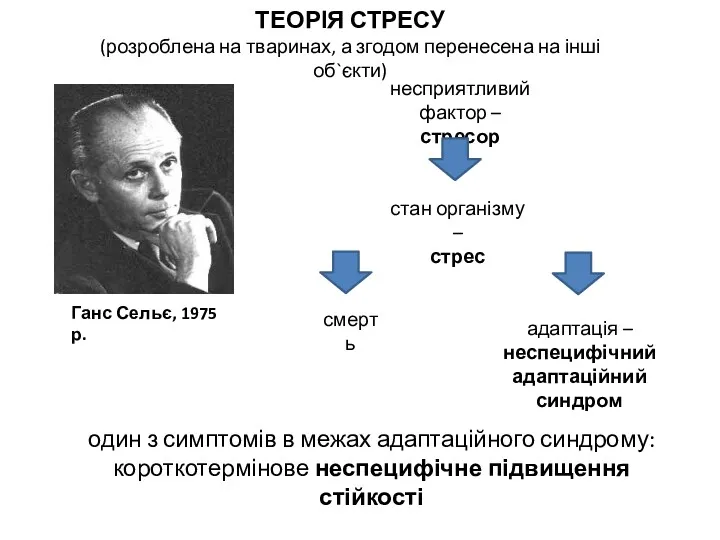
- 8. ТЕОРІЯ СТРЕСУ (розроблена на тваринах, а згодом перенесена на інші об`єкти) Ганс Сельє, 1975 р. несприятливий
- 9. Індукована стійкість рослин подібна до мобілізації під час військового стану, коли значна частина ресурсів замість нормальних
- 10. Елісітор – біотичний або абіотичний фактор, що індукує в рослинах неспецифічний комплекс захисних реакцій (неспецифічний адаптаційний
- 11. ОСНОВНІ ШЛЯХИ ЗАПУСКУ ІНДУКОВАНОЇ СТІЙКОСТІ саліцилатний (SA) жасмонатний (JA) у відповідь на дію біотрофних паразитів та
- 12. Іnduced Systemic Resistance – ISR System Acquired Resistance – SAR Pieterse et al., Nature Chemical Biology,
- 14. Simplified schematic representation of the SA, JA and ET signaling pathways SA mediates a change in
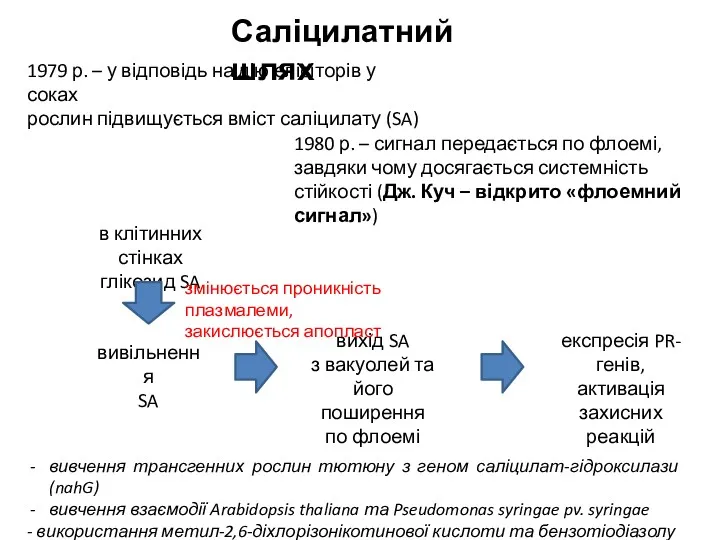
- 15. в клітинних стінках глікозид SA Саліцилатний шлях 1979 р. – у відповідь на дію елісіторів у
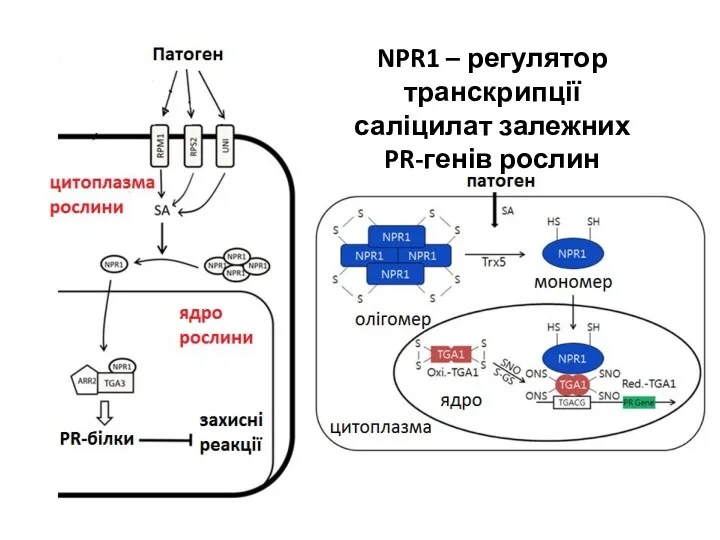
- 16. NPR1 – регулятор транскрипції саліцилат залежних PR-генів рослин
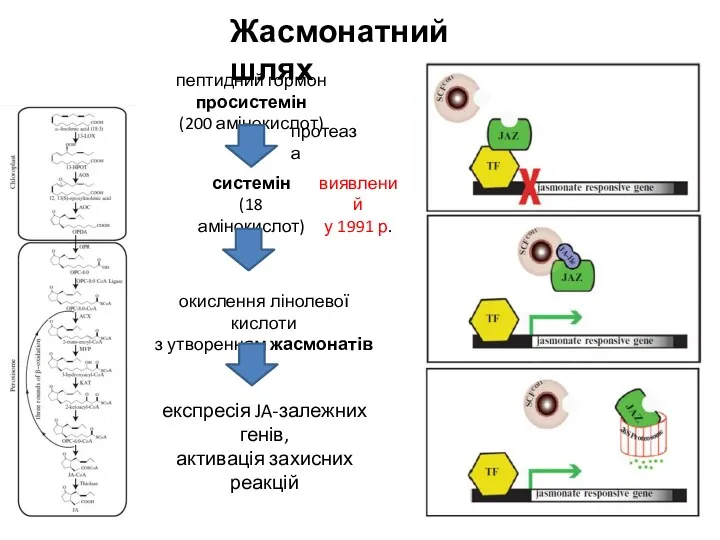
- 17. Жасмонатний шлях пептидний гормон просистемін (200 амінокислот) системін (18 амінокислот) окислення лінолевої кислоти з утворенням жасмонатів
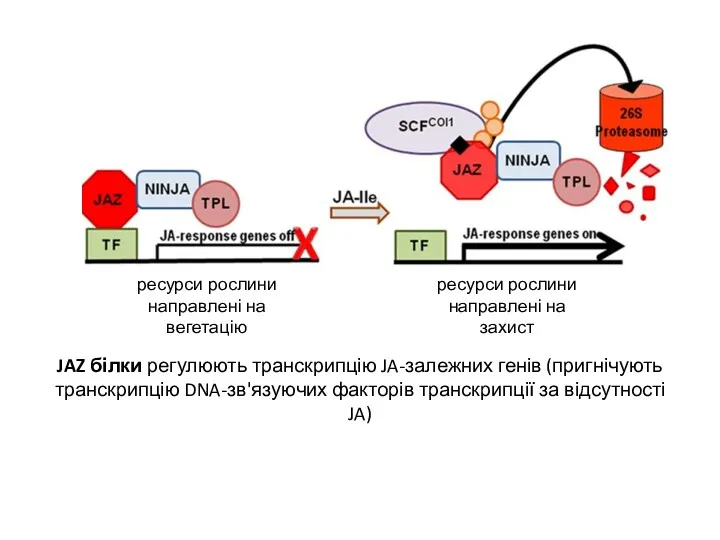
- 18. JAZ білки регулюють транскрипцію JA-залежних генів (пригнічують транскрипцію DNA-зв'язуючих факторів транскрипції за відсутності JA) JA-isoleucine
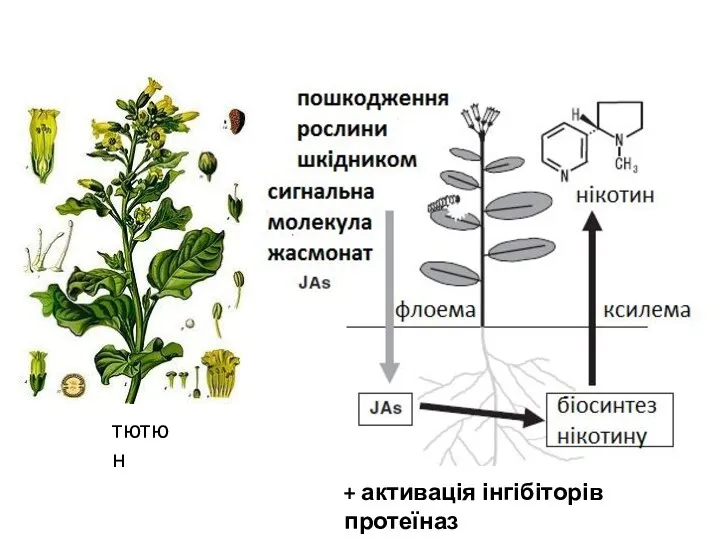
- 19. + активація інгібіторів протеїназ тютюн
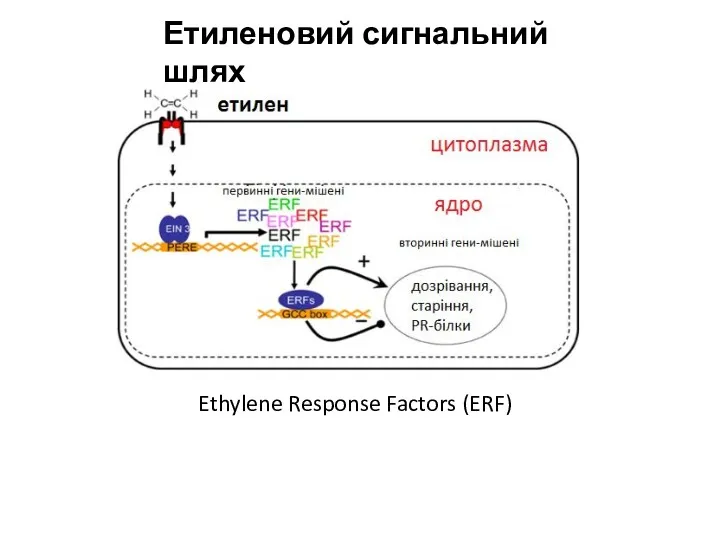
- 20. Етиленовий сигнальний шлях Ethylene Response Factors (ERF)
- 21. 2012 р.
- 23. Harold Henry Flor (1900 – 1991) СПЕЦИФІЧНА СТІЙКІСТЬ РОСЛИН Теорія «ген-на-ген» (1940-1955 рр.) іржа льону (пасмо)
- 24. Збудник іржі льону має багато (понад 100) рас Певні сорти (сорти-диференціатори) здатні розпізнавати певні раси паразита
- 26. Протягом тривалого часу продукти R- та Vir-генів залишалися невідомими, тому механізми расоспецифічної (моногенної) стійкості рослин залишалися
- 27. гени вірулентності перейменовані на гени авірулентності Avr-гени
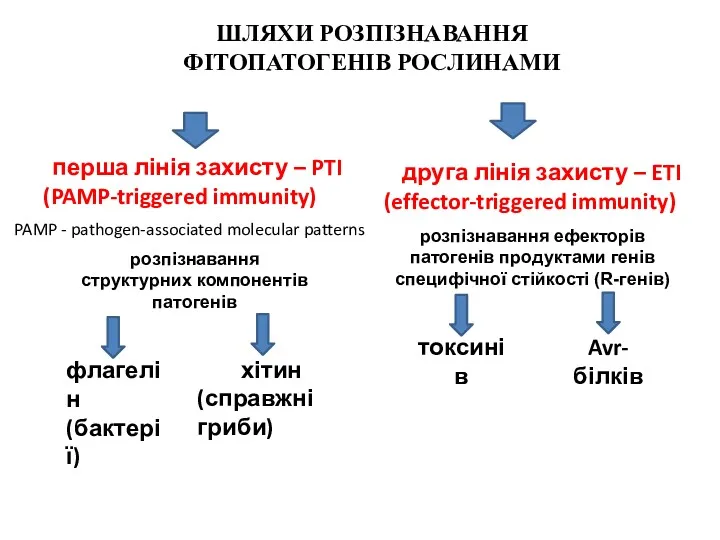
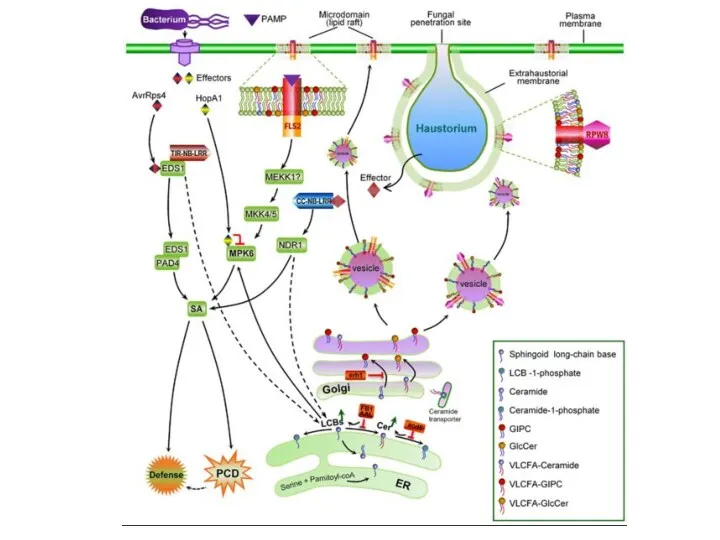
- 28. ШЛЯХИ РОЗПІЗНАВАННЯ ФІТОПАТОГЕНІВ РОСЛИНАМИ перша лінія захисту – PTI (PAMP-triggered immunity) розпізнавання структурних компонентів патогенів друга
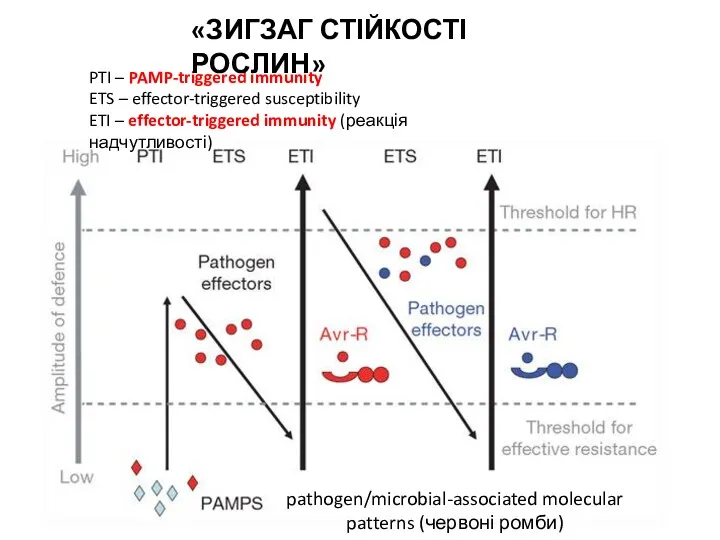
- 29. pathogen/microbial-associated molecular patterns (червоні ромби) PTI – PAMP-triggered immunity ETS – effector-triggered susceptibility ETI – effector-triggered
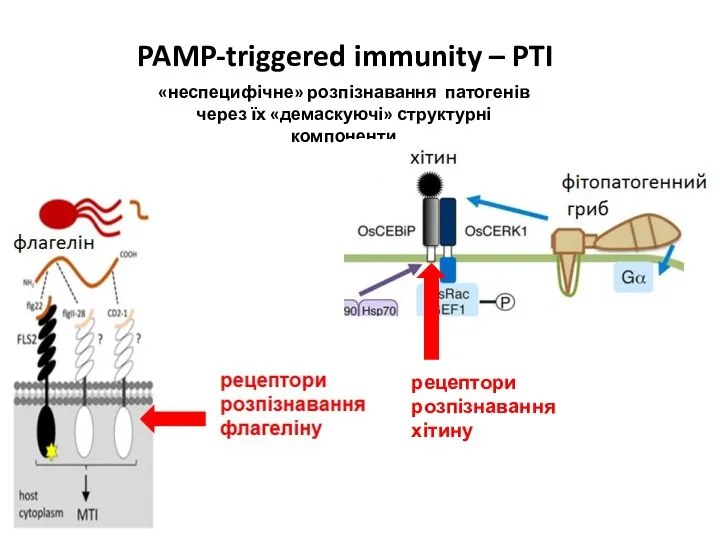
- 30. «неспецифічне» розпізнавання патогенів через їх «демаскуючі» структурні компоненти PAMP-triggered immunity – PTI
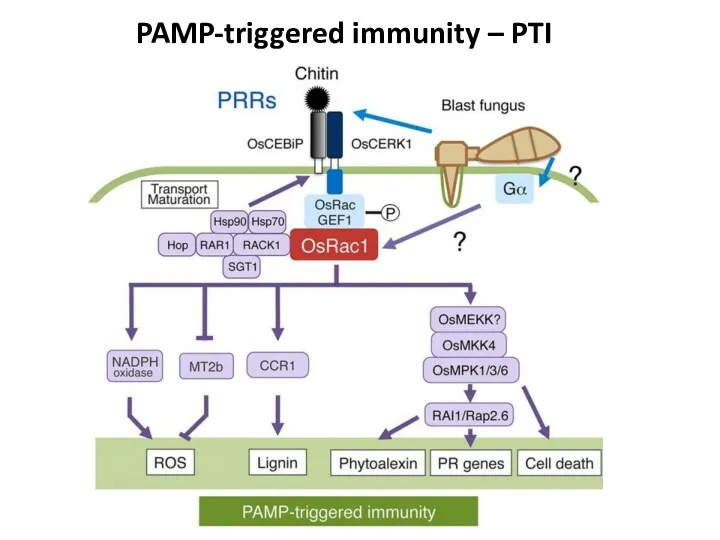
- 31. PAMP-triggered immunity – PTI
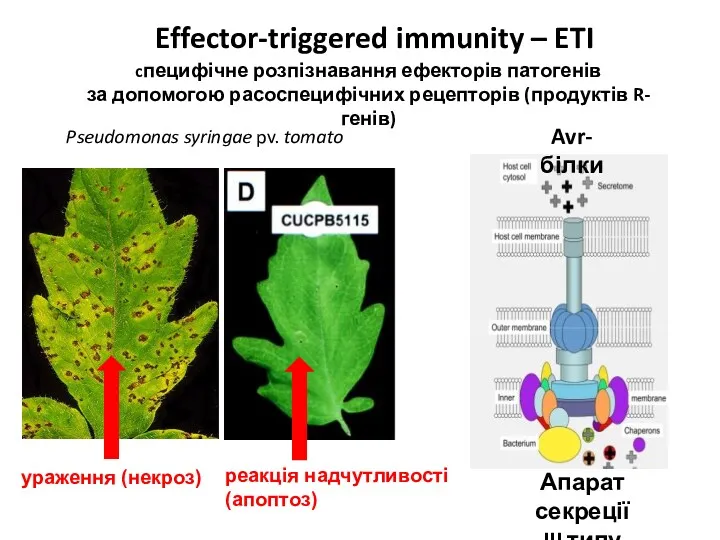
- 32. Effector-triggered immunity – ETI cпецифічне розпізнавання ефекторів патогенів за допомогою расоспецифічних рецепторів (продуктів R-генів) Pseudomonas syringae
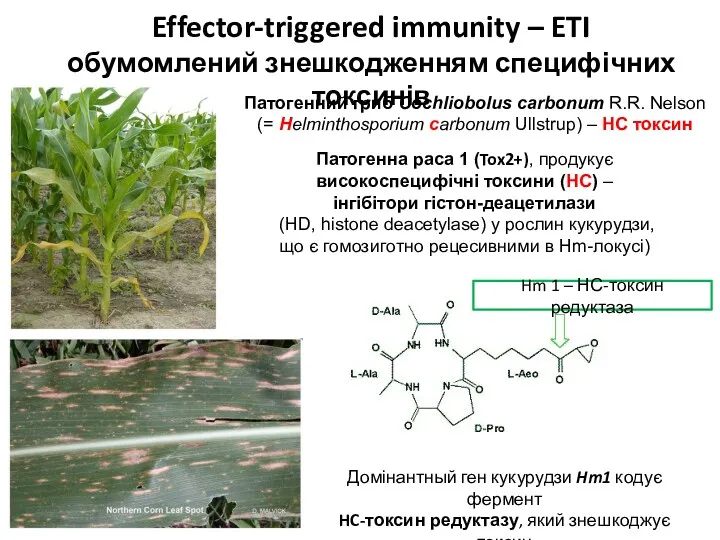
- 33. Патогенний гриб Cochliobolus carbonum R.R. Nelson (= Helminthosporium carbonum Ullstrup) – HC токсин Патогенна раса 1
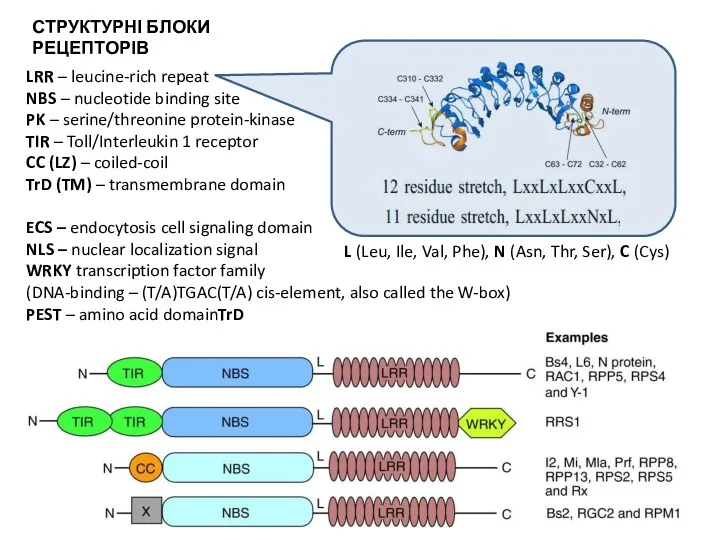
- 34. СТРУКТУРА СПЕЦИФІЧНИХ РЕЦЕПТОРІВ РОСЛИН
- 35. LRR – leucine-rich repeat NBS – nucleotide binding site PK – serine/threonine protein-kinase TIR – Toll/Interleukin
- 38. НОВІТНЯ КЛАСИФИКАЦІЯ СПЕЦИФІЧНИХ РЕЦЕПТОРІВ РОСЛИН
- 41. Приклади сигнальних шляхів при запуску реакції надчутливості окислювальний спалах сигнальна система NO активується аргінін-залежна NO-синтаза (NOS)
- 42. каскади MAP-кіназ
- 46. СИГНАЛЬНА СИСТЕМА NO 1992
- 47. NO атмосфери дуже токсичний для рослин, але присутній там у надмалих кількостях 1996-97 рр. – перші
- 48. L-аргінін + кисень = L-цитрулін + NO СИГНАЛЬНА СИСТЕМА NO
- 49. пероксінітріт нітрозо-глутатіон (флоемний сигнал)
- 50. потовщення клітинних стінок синтез PR-білків окислювальний спалах запрограмована гибель клітин… інактивація асоційованих з хлоропластами генів, зниження
- 51. Генетическая природа устойчивости растений Полигенная концепция (Р. Фишер, 1930; К. Мазер, 1943) Моногенная концепция (Биффен, 1930;
- 52. Теория горизонтальной и вертикальной устойчивости (1963-1985 гг.) фитофтороз картофеля, гриб Phytophthora infestans Ван дер Планк Я.Е.
- 53. Горизонтальная устойчивость - неспецифическое взаимодействие сорта со всеми расами патогена, имеет невысокий (количественный) уровень проявления, сильно
- 55. Скачать презентацию
ІНДУКОВАНА (НЕСПЕЦИФІЧНА НАБУТА) СТІЙКІСТЬ РОСЛИН
System Acquired Resistance – SAR
+
Іnduced Systemic Resistance – ISR
Пауль
ІНДУКОВАНА (НЕСПЕЦИФІЧНА НАБУТА) СТІЙКІСТЬ РОСЛИН
System Acquired Resistance – SAR
+
Іnduced Systemic Resistance – ISR
Пауль

перший експеримент по «імунізації» рослин
(вакцинація рослин)
Дж. Бовери та Дж. Рай (1901 р.)
аналогічні
перший експеримент по «імунізації» рослин
(вакцинація рослин)
Дж. Бовери та Дж. Рай (1901 р.)
аналогічні
Які механізми набутої стійкості рослин після «вакцинації»?
Чи є у рослин антитіла?
В. Блекмен (1922
Які механізми набутої стійкості рослин після «вакцинації»?
Чи є у рослин антитіла?
В. Блекмен (1922
Станом на 1930-і рр. було встановлено, що стійкість рослин,
що виникає після впливу
Станом на 1930-і рр. було встановлено, що стійкість рослин,
що виникає після впливу

ТЕОРІЯ ФІТОАЛЕКСИНІВ
К.О. Мюллер та X. Бергер, 1941 р.
(дослідження вірулентної та авірулентної рас
збудника фітофторозу
ТЕОРІЯ ФІТОАЛЕКСИНІВ
К.О. Мюллер та X. Бергер, 1941 р.
(дослідження вірулентної та авірулентної рас
збудника фітофторозу
Фітоалексини відсутні у здоровій рослині і синтезуються (de novo) у відповідь на зараження
Фітоалексини
Фітоалексини відсутні у здоровій рослині і синтезуються (de novo) у відповідь на зараження
Фітоалексини

ТЕОРІЯ СТРЕСУ
(розроблена на тваринах, а згодом перенесена на інші об`єкти)
Ганс Сельє, 1975
ТЕОРІЯ СТРЕСУ
(розроблена на тваринах, а згодом перенесена на інші об`єкти)
Ганс Сельє, 1975
Індукована стійкість рослин подібна до мобілізації
під час військового стану, коли значна частина ресурсів
Індукована стійкість рослин подібна до мобілізації
під час військового стану, коли значна частина ресурсів

Елісітор – біотичний або абіотичний фактор, що індукує
в рослинах неспецифічний комплекс захисних
Елісітор – біотичний або абіотичний фактор, що індукує
в рослинах неспецифічний комплекс захисних

ОСНОВНІ ШЛЯХИ ЗАПУСКУ ІНДУКОВАНОЇ СТІЙКОСТІ
саліцилатний (SA)
жасмонатний (JA)
у відповідь на дію
біотрофних паразитів
та сисинх (флоемних)
шкідників
у
ОСНОВНІ ШЛЯХИ ЗАПУСКУ ІНДУКОВАНОЇ СТІЙКОСТІ
саліцилатний (SA)
жасмонатний (JA)
у відповідь на дію
біотрофних паразитів
та сисинх (флоемних)
шкідників
у

Іnduced Systemic Resistance – ISR
System Acquired Resistance – SAR
Pieterse et al., Nature Chemical
Іnduced Systemic Resistance – ISR
System Acquired Resistance – SAR
Pieterse et al., Nature Chemical
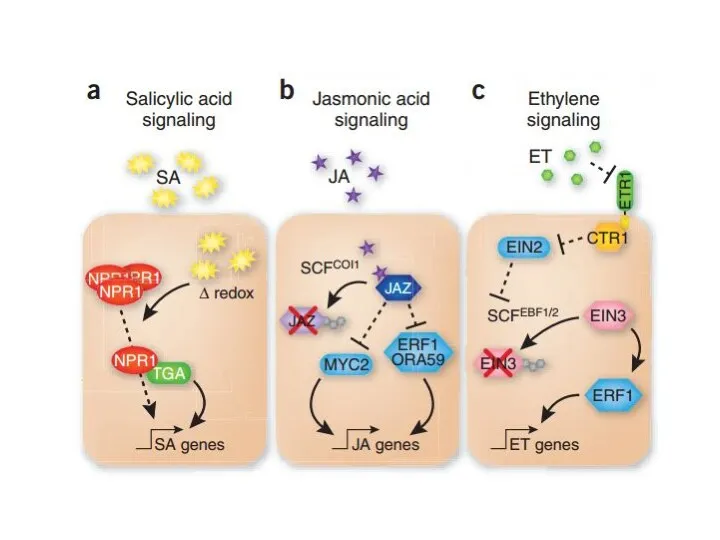
Simplified schematic representation of the SA, JA and ET signaling pathways
SA mediates a
Simplified schematic representation of the SA, JA and ET signaling pathways
SA mediates a

в клітинних стінках
глікозид SA
Саліцилатний шлях
1979 р. – у відповідь на дію елісіторів у
в клітинних стінках
глікозид SA
Саліцилатний шлях
1979 р. – у відповідь на дію елісіторів у
NPR1 – регулятор транскрипції
саліцилат залежних
PR-генів рослин
NPR1 – регулятор транскрипції
саліцилат залежних
PR-генів рослин
Жасмонатний шлях
пептидний гормон просистемін
(200 амінокислот)
системін
(18 амінокислот)
окислення лінолевої кислоти
з утворенням жасмонатів
експресія
Жасмонатний шлях
пептидний гормон просистемін
(200 амінокислот)
системін
(18 амінокислот)
окислення лінолевої кислоти
з утворенням жасмонатів
експресія
JAZ білки регулюють транскрипцію JA-залежних генів (пригнічують транскрипцію DNA-зв'язуючих факторів транскрипції за відсутності
JAZ білки регулюють транскрипцію JA-залежних генів (пригнічують транскрипцію DNA-зв'язуючих факторів транскрипції за відсутності
+ активація інгібіторів протеїназ
тютюн
+ активація інгібіторів протеїназ
тютюн
Етиленовий сигнальний шлях
Ethylene Response Factors (ERF)
Етиленовий сигнальний шлях
Ethylene Response Factors (ERF)
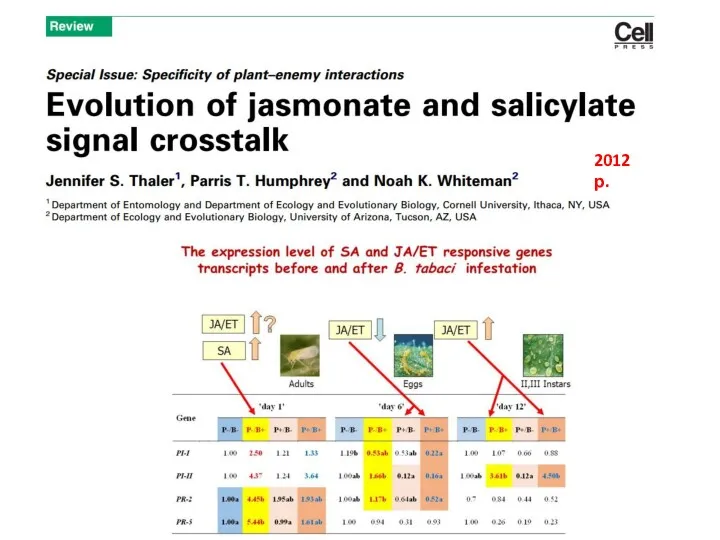
2012 р.
2012 р.

Harold Henry Flor
(1900 – 1991)
СПЕЦИФІЧНА СТІЙКІСТЬ РОСЛИН
Теорія «ген-на-ген» (1940-1955 рр.)
іржа льону (пасмо) →
Harold Henry Flor
(1900 – 1991)
СПЕЦИФІЧНА СТІЙКІСТЬ РОСЛИН
Теорія «ген-на-ген» (1940-1955 рр.)
іржа льону (пасмо) →

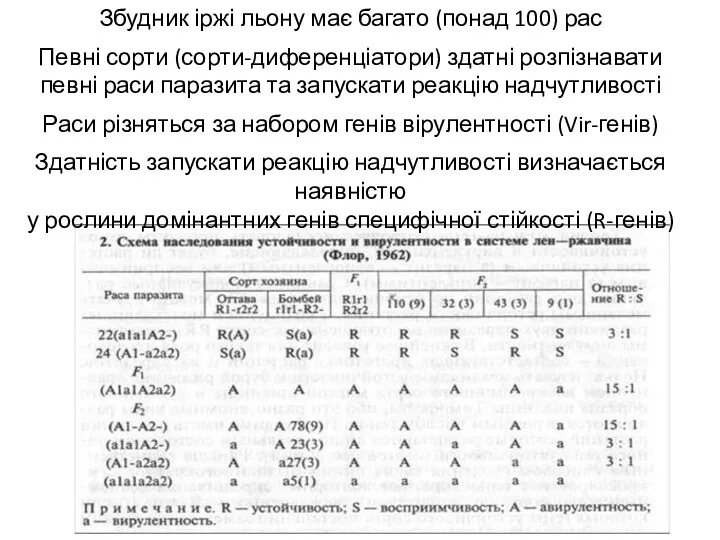
Збудник іржі льону має багато (понад 100) рас
Певні сорти (сорти-диференціатори) здатні розпізнавати
певні
Збудник іржі льону має багато (понад 100) рас
Певні сорти (сорти-диференціатори) здатні розпізнавати
певні
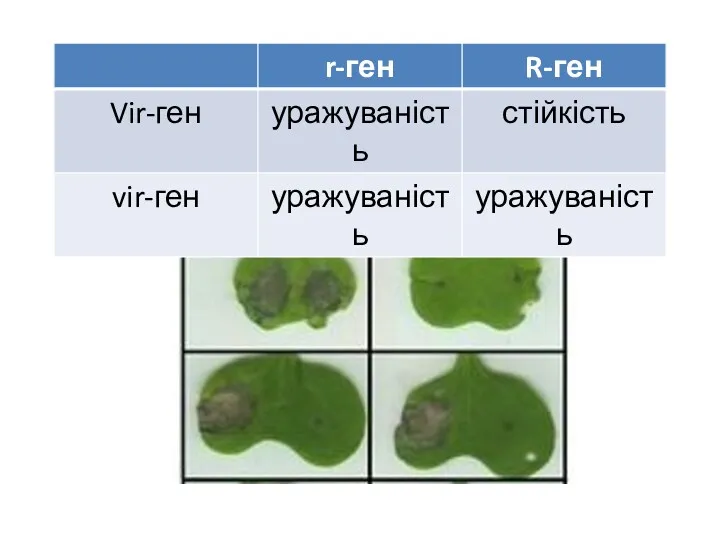
Протягом тривалого часу продукти R- та Vir-генів залишалися невідомими,
тому механізми расоспецифічної (моногенної)
Протягом тривалого часу продукти R- та Vir-генів залишалися невідомими,
тому механізми расоспецифічної (моногенної)

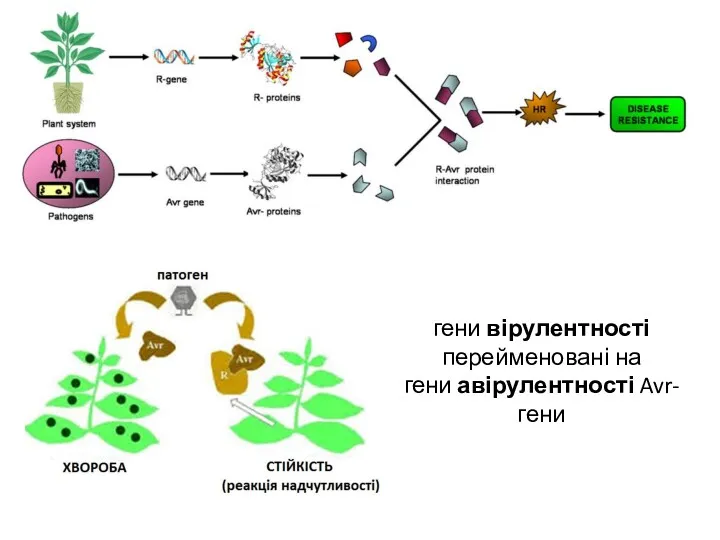
гени вірулентності
перейменовані на
гени авірулентності Avr-гени
гени вірулентності
перейменовані на
гени авірулентності Avr-гени
ШЛЯХИ РОЗПІЗНАВАННЯ
ФІТОПАТОГЕНІВ РОСЛИНАМИ
перша лінія захисту – PTI
(PAMP-triggered immunity)
розпізнавання
структурних компонентів
патогенів
ШЛЯХИ РОЗПІЗНАВАННЯ
ФІТОПАТОГЕНІВ РОСЛИНАМИ
перша лінія захисту – PTI
(PAMP-triggered immunity)
розпізнавання
структурних компонентів
патогенів
pathogen/microbial-associated molecular patterns (червоні ромби)
PTI – PAMP-triggered immunity
ETS – effector-triggered susceptibility
ETI – effector-triggered
pathogen/microbial-associated molecular patterns (червоні ромби)
PTI – PAMP-triggered immunity
ETS – effector-triggered susceptibility
ETI – effector-triggered
«неспецифічне» розпізнавання патогенів через їх «демаскуючі» структурні компоненти
PAMP-triggered immunity – PTI
«неспецифічне» розпізнавання патогенів через їх «демаскуючі» структурні компоненти
PAMP-triggered immunity – PTI
PAMP-triggered immunity – PTI
PAMP-triggered immunity – PTI
Effector-triggered immunity – ETI
cпецифічне розпізнавання ефекторів патогенів
за допомогою расоспецифічних рецепторів (продуктів R-генів)
Pseudomonas
Effector-triggered immunity – ETI
cпецифічне розпізнавання ефекторів патогенів
за допомогою расоспецифічних рецепторів (продуктів R-генів)
Pseudomonas
Патогенний гриб Cochliobolus carbonum R.R. Nelson
(= Helminthosporium carbonum Ullstrup) – HC токсин
Патогенна раса 1 (Tox2+),
Патогенний гриб Cochliobolus carbonum R.R. Nelson
(= Helminthosporium carbonum Ullstrup) – HC токсин
Патогенна раса 1 (Tox2+),
СТРУКТУРА СПЕЦИФІЧНИХ РЕЦЕПТОРІВ РОСЛИН
СТРУКТУРА СПЕЦИФІЧНИХ РЕЦЕПТОРІВ РОСЛИН

LRR – leucine-rich repeat
NBS – nucleotide binding site
PK – serine/threonine protein-kinase
TIR – Toll/Interleukin
LRR – leucine-rich repeat
NBS – nucleotide binding site
PK – serine/threonine protein-kinase
TIR – Toll/Interleukin
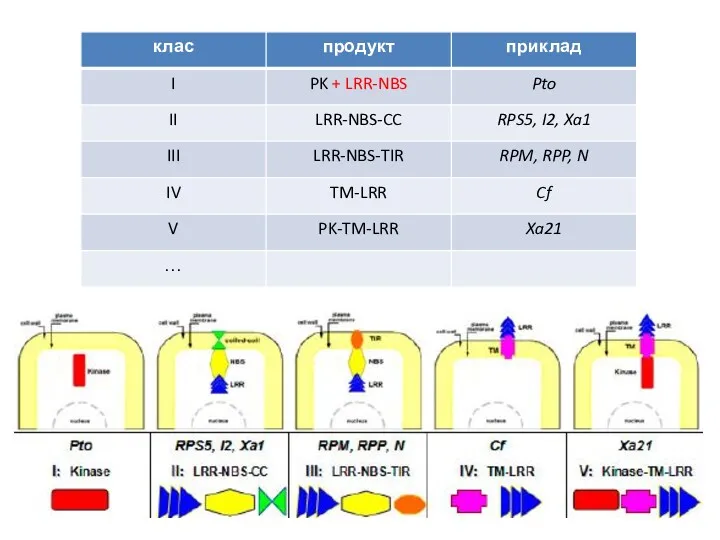
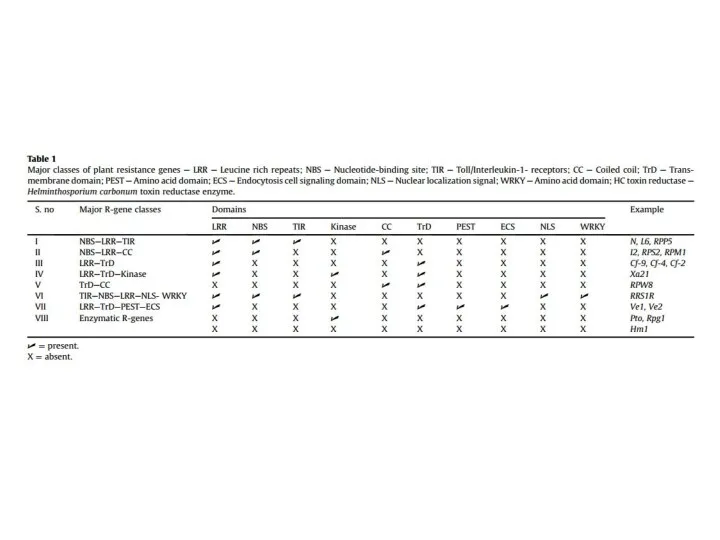
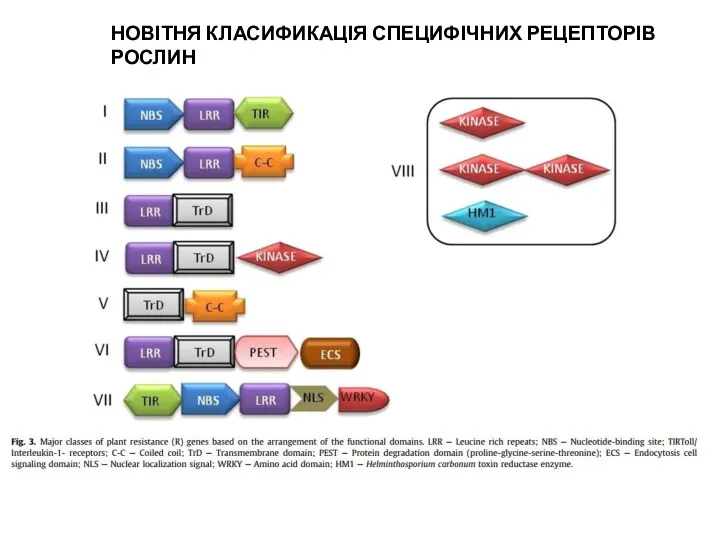
НОВІТНЯ КЛАСИФИКАЦІЯ СПЕЦИФІЧНИХ РЕЦЕПТОРІВ РОСЛИН
НОВІТНЯ КЛАСИФИКАЦІЯ СПЕЦИФІЧНИХ РЕЦЕПТОРІВ РОСЛИН

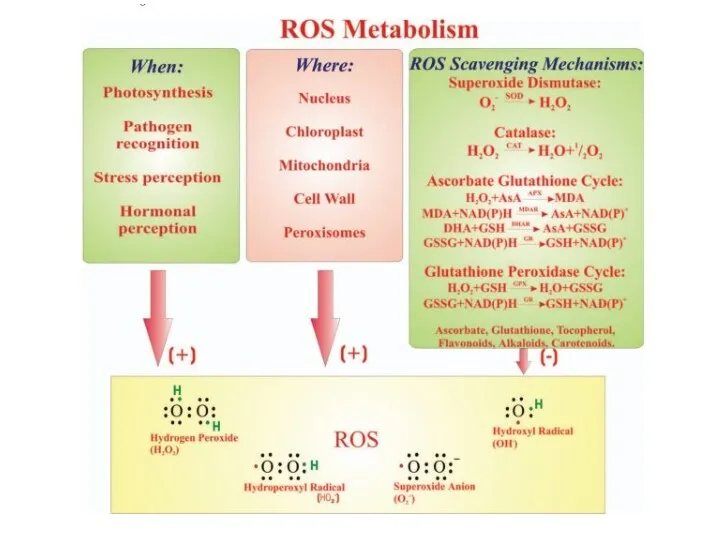
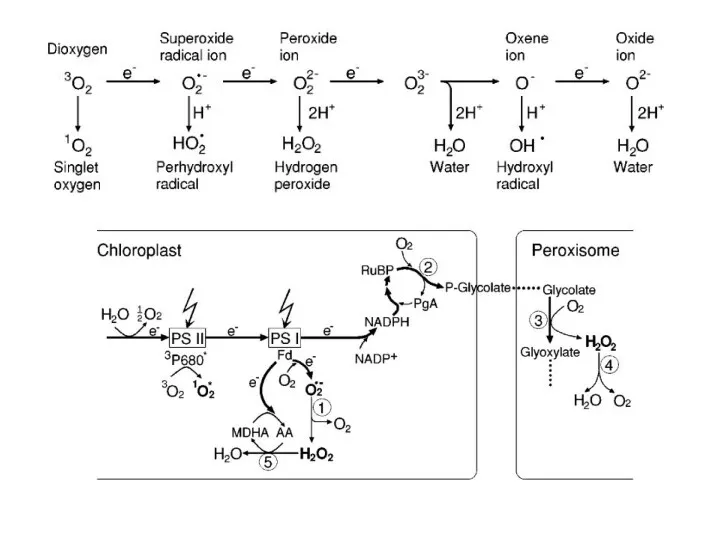
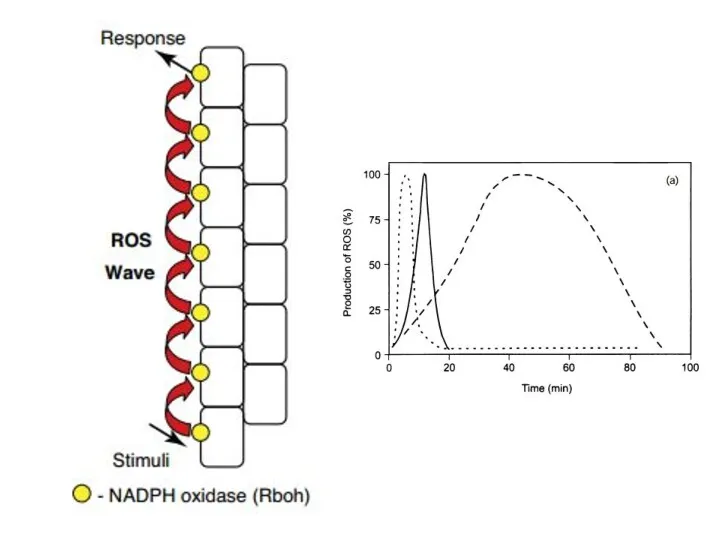
Приклади сигнальних шляхів при запуску реакції надчутливості
окислювальний
спалах
сигнальна
система NO
активується
аргінін-залежна
NO-синтаза (NOS)
активується
НАДФН-оксидаза
(NADPH-oxidase)
каскади
Приклади сигнальних шляхів при запуску реакції надчутливості
окислювальний
спалах
сигнальна
система NO
активується
аргінін-залежна
NO-синтаза (NOS)
активується
НАДФН-оксидаза
(NADPH-oxidase)
каскади
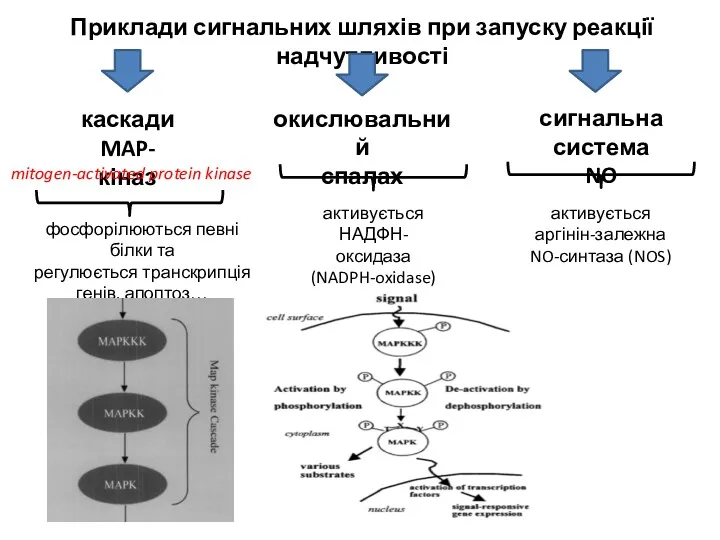
каскади MAP-кіназ
каскади MAP-кіназ
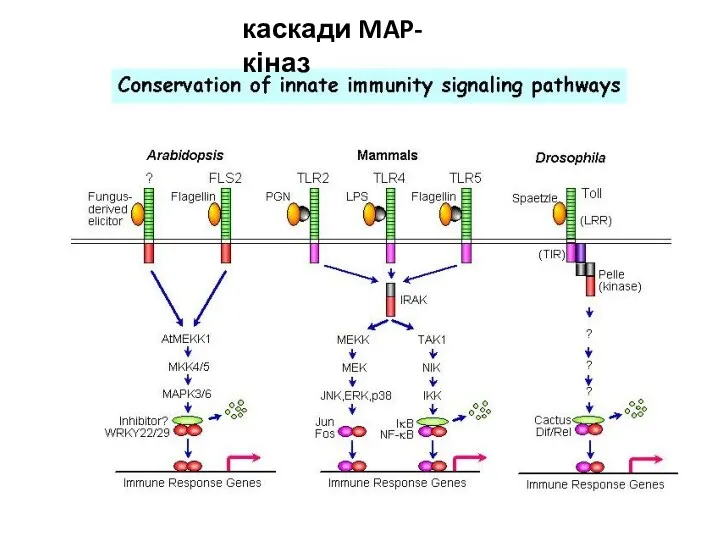
СИГНАЛЬНА СИСТЕМА NO
1992
СИГНАЛЬНА СИСТЕМА NO
1992

NO атмосфери дуже токсичний для рослин,
але присутній там у надмалих кількостях
1996-97 рр.
NO атмосфери дуже токсичний для рослин,
але присутній там у надмалих кількостях
1996-97 рр.

L-аргінін + кисень =
L-цитрулін + NO
СИГНАЛЬНА СИСТЕМА NO
L-аргінін + кисень =
L-цитрулін + NO
СИГНАЛЬНА СИСТЕМА NO
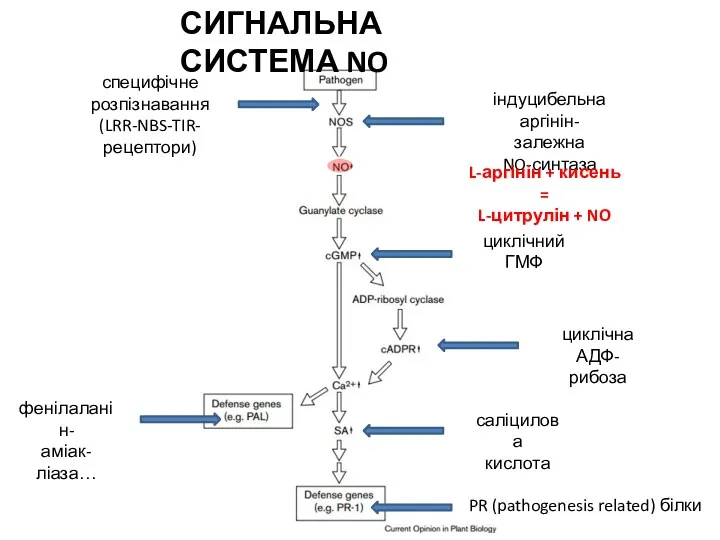
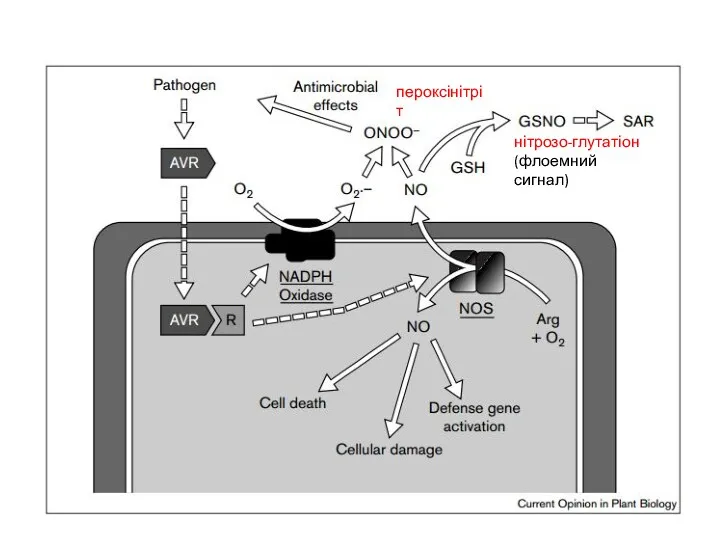
пероксінітріт
нітрозо-глутатіон
(флоемний сигнал)
пероксінітріт
нітрозо-глутатіон
(флоемний сигнал)
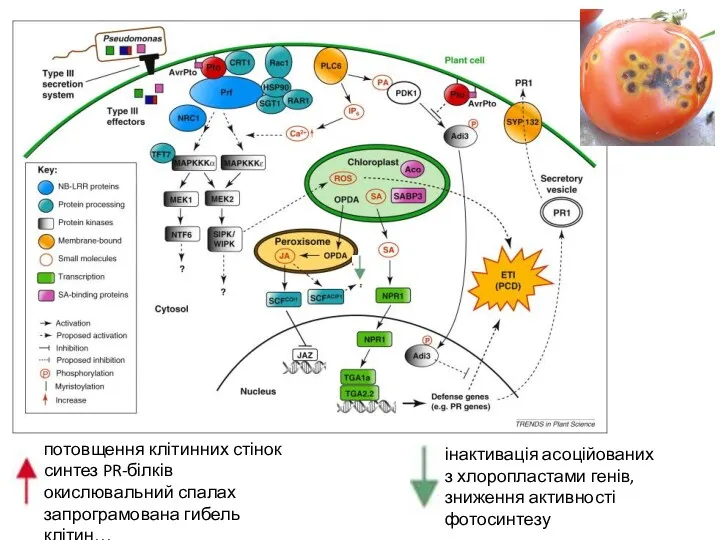
потовщення клітинних стінок
синтез PR-білків
окислювальний спалах
запрограмована гибель клітин…
інактивація асоційованих
з хлоропластами генів,
зниження активності фотосинтезу
потовщення клітинних стінок
синтез PR-білків
окислювальний спалах
запрограмована гибель клітин…
інактивація асоційованих
з хлоропластами генів,
зниження активності фотосинтезу
Генетическая природа устойчивости растений
Полигенная концепция
(Р. Фишер, 1930; К. Мазер, 1943)
Моногенная концепция
(Биффен,
Генетическая природа устойчивости растений
Полигенная концепция
(Р. Фишер, 1930; К. Мазер, 1943)
Моногенная концепция
(Биффен,

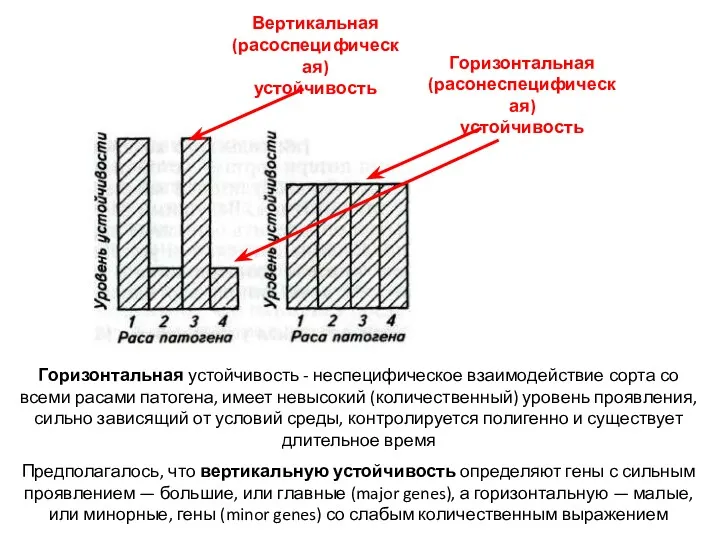
Теория горизонтальной и вертикальной устойчивости (1963-1985 гг.)
фитофтороз картофеля, гриб Phytophthora infestans
Ван дер Планк Я.Е.
Теория горизонтальной и вертикальной устойчивости (1963-1985 гг.)
фитофтороз картофеля, гриб Phytophthora infestans
Ван дер Планк Я.Е.

Горизонтальная устойчивость - неспецифическое взаимодействие сорта со всеми расами патогена, имеет невысокий (количественный)
Горизонтальная устойчивость - неспецифическое взаимодействие сорта со всеми расами патогена, имеет невысокий (количественный)
 Класс млекопитающие
Класс млекопитающие Жизнь домашних животных
Жизнь домашних животных Эпителиальная ткань
Эпителиальная ткань Анаэробный обмен углеводов
Анаэробный обмен углеводов Плазмиды
Плазмиды Птахи нашої місцевості
Птахи нашої місцевості Лимбическая система
Лимбическая система Технология выращивания астры китайской
Технология выращивания астры китайской Кайнозойская эра
Кайнозойская эра Психические познавательные процессы
Психические познавательные процессы Водоём - дом из воды
Водоём - дом из воды Человеческий мозг
Человеческий мозг Эпителий ұлпаларының жүйелері
Эпителий ұлпаларының жүйелері Группа Лишайники
Группа Лишайники Постэмбриональное развитие животных
Постэмбриональное развитие животных Реактивний рух у природі
Реактивний рух у природі Нейрон,түрлері,қызметі. Нейроглия.жүйке талшығынан қозудың өту механизмі. Синапс,қозудың өту механизмі
Нейрон,түрлері,қызметі. Нейроглия.жүйке талшығынан қозудың өту механизмі. Синапс,қозудың өту механизмі Птицы нашей местности
Птицы нашей местности Птицы в Санкт-Петербурге
Птицы в Санкт-Петербурге Клеточное строение растений
Клеточное строение растений Презентация к внеклассному мероприятию Деградация почв
Презентация к внеклассному мероприятию Деградация почв Genetics. Mendelian principles
Genetics. Mendelian principles Первоцветы. Биологические особенности раннецветущих растений
Первоцветы. Биологические особенности раннецветущих растений Растительный организм и его особенности. Тема 16
Растительный организм и его особенности. Тема 16 Генетика бастауы – Морган заңдылықтары
Генетика бастауы – Морган заңдылықтары Животные национальные символы стран мира
Животные национальные символы стран мира Ягуар 2
Ягуар 2 Какие функции выполняет корень?
Какие функции выполняет корень?