- Молекулярно-генетические методы диагностики в практике врача. Лекция 9

Содержание
- 2. В ядре каждой соматической клетки организма человека содержится 46 хромосом (44 аутосомы и 2 половые) или
- 3. Строение хромосом Материал, из которого построены хромосомы – хроматин – представляет собой комплекс ДНК и белков,
- 4. 1. Молекула ДНК - элементарная структурная ед. ДНК (мономер) – нуклеотид – состав азотистое основание [пурины:
- 5. 2. Через определенные промежутки ДНК обвивается 1¾ раза (145 п.о.) вокруг нуклеосомного кора (кор – гистоновый
- 6. Затем следует участок свободной ДНК 20 – 60 п.н. В результате образуется нуклеосомная цепь (волокно) -
- 7. Центромера – компактный участок хромосомы, к которому при делении клетки прикрепляются волокна веретена p, q-плечи Теломеры
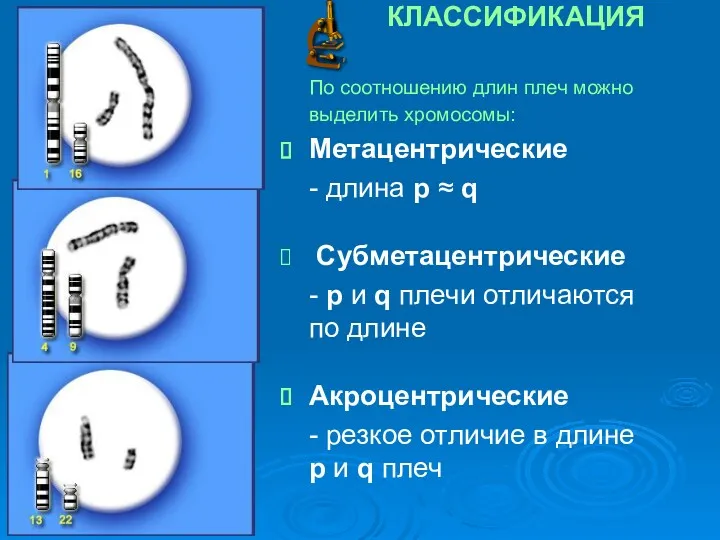
- 8. КЛАССИФИКАЦИЯ По соотношению длин плеч можно выделить хромосомы: Метацентрические - длина p ≈ q Субметацентрические -
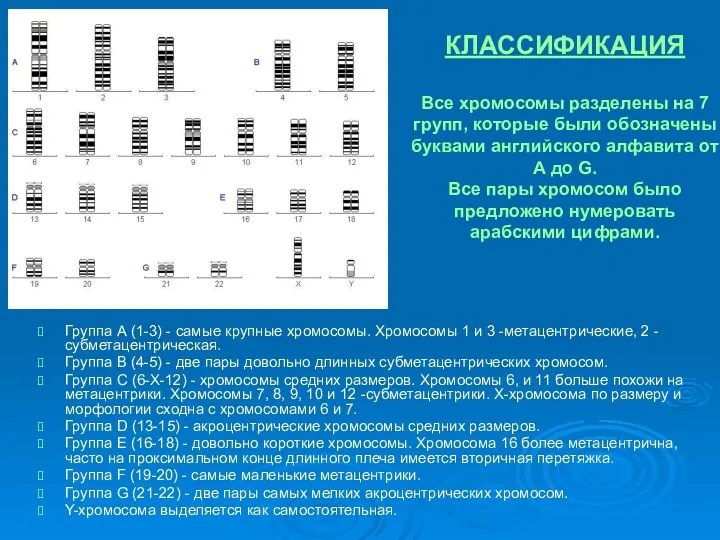
- 9. КЛАССИФИКАЦИЯ Все хромосомы разделены на 7 групп, которые были обозначены буквами английского алфавита от А до
- 10. Лучше и удобнее всего анализировать хромосомы в те моменты клеточного цикла, когда их можно визуализировать с
- 11. ОСНОВНЫЕ МЕТОДЫ ПРИГОТОВЛЕНИЯ ХРОМОСОМНЫХ ПРЕПАРАТОВ В качестве материала для получения хромосом человека и их исследования могут
- 12. Основные методы приготовления хромосомных препаратов ЭТАПЫ: 1) Забор материала – ткань, содержащая способные к делению клетки
- 13. Культивирование клеток периферической крови, постановка культуры лимфоцитов человека. Забор материала. Культивирование на питательной среде. Используются цельная
- 14. Культивирование клеток периферической крови, постановка культуры лимфоцитов человека. Остановка деления клеток на стадии метафазы. Гипотонизация. Фиксация.
- 15. Приготовление хромосомных препаратов Задачей процедур этого этапа является получение хорошо распластанных метафазных пластинок с сохранением целостности
- 16. ОКРАСКА Метод сплошной окраски (RV-метод). Окраска азур-эозином: 1) Готовится 0,1% раствор эозина и 0,1% раствор азура
- 17. Методы дифференциальных окрасок хромосом, выявляющих их линейную неоднородность. В практике широко применены следующие методы: метод окраски
- 18. G-окрашивание Препараты обрабатываются 0,25% р-ром трипсина Промываются в 70%, 96%, 100% этаноле Высушиваются Окрашиваются 5% р-ром
- 19. R-окрашивание «Состарившиеся» хромосомные препараты инкубируют при t=60oC в насыщенном растворе гидроокиси бария 3 – 5 мин.
- 20. Ряд методов окраски позволяет получить поперечную исчерченность хромосом (R-, G-, Q-band). Полосы размещаются по длине плеч
- 21. Полосы и участки нумеруются в направлении от центромеры к теломере по длине короткого и длинного плеча.
- 22. В любом обозначении хромосомы используются: 1) номер хромосомы, 2) символ плеча, 3) номер района (участка), 4)
- 23. Цитогенетическое исследование [G-band]
- 24. Основные принципы анализа хромосомных препаратов Цитогенетический анализ хромосом проводят на препаратах дифференциально окрашенных по длине (G-,
- 25. Внедрение методов дифференциальной окраски хромосом способствовало значительному прогрессу в цитогенетике человека и привело к крупным достижениям.
- 26. МОЛЕКУЛЯРНО-ЦИТОГЕНЕТИЧЕСКОЕ ИССЛЕДОВАНИЕ Базируется на технологии гибридизации ДНК. Позволяет увидеть хромосомную локализацию специфических последовательностей ДНК с помощью
- 27. МОЛЕКУЛЯРНО-ЦИТОГЕНЕТИЧЕСКОЕ ИССЛЕДОВАНИЕ ЭТАПЫ: 1) 1) – 6) Этапы аналогичны этапам цитогенетического исследования Денатурация образца и зонда
- 28. МОЛЕКУЛЯРНО-ЦИТОГЕНЕТИЧЕСКОЕ ИССЛЕДОВАНИЕ При наличии комплементарного зонду участка последовательности ДНК – визуализация (хромосомы, ядра с флуоресцентными сигналами)
- 29. МОЛЕКУЛЯРНО-ЦИТОГЕНЕТИЧЕСКОЕ ИССЛЕДОВАНИЕ – флуоресцентная гибридизация in situ (FISH).
- 30. МОЛЕКУЛЯРНО-ЦИТОГЕНЕТИЧЕСКОЕ ИССЛЕДОВАНИЕ Новые молекулярно-цитогенетические технологии – анализ многоцветно окрашенных хромосом – спектральное кариотипитование (SKY) и мультиплексная
- 31. МОЛЕКУЛЯРНО-ЦИТОГЕНЕТИЧЕСКОЕ ИССЛЕДОВАНИЕ Спектральное кариотипитование (SKY)
- 32. МОЛЕКУЛЯРНО-ЦИТОГЕНЕТИЧЕСКОЕ ИССЛЕДОВАНИЕ Мультиплексная FISH (M-FISH)
- 33. МОЛЕКУЛЯРНО-ЦИТОГЕНЕТИЧЕСКОЕ ИССЛЕДОВАНИЕ Сравнительная геномная гибридизация (CGH) – техника, позволяющая широко сканировать геном на потерю или увеличение
- 34. Контрольная геномная ДНК (известный нормальный кариотип) Тестируемая геномная ДНК (неизвестный кариотип) Сравнительная геномная гибридизация (CGH)
- 35. Сравнительная геномная гибридизация (CGH)
- 36. МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИЕ МЕТОДЫ: ГЕНЕТИЧЕСКАЯ ИНЖЕНЕРИЯ (ТЕХНОЛОГИЯ РЕКОМБИНАНТНЫХ ДНК) Методы клонирования ДНК Методы анализа первичной последовательности
- 37. Клонирование ДНК – позволяет из единичного фрагмента ДНК получить достаточно большое количество его копий, чтобы получить
- 38. МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИЕ МЕТОДЫ: Клонирование in vivo Фрагмент ДНК, вырезанный с помощью рестриктаз, соединяется с молекулой вектора с
- 39. Клонирование вне клеток. Полимеразная цепная реакция (ПЦР) Выделение ДНК Приготовление реакционной смеси -образец ДНК (матрица) -праймеры
- 40. МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИЕ МЕТОДЫ. Амплификация → увеличение количества повторов интересующего участка. Процесс – цикличное изменение температурного режима. -плавление
- 41. Фрагмент для размножения Цикл 2 Плавление и отжиг праймеров Элонгация - DNA Taq - полимераза Полимеразная
- 42. Полимеразная цепная реакция 300 bp 500 bp 20-40 циклов 1 2 1 2
- 43. МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИЕ МЕТОДЫ. Полимеразная цепная реакция Гель-электрофорез Детекция
- 44. МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИЕ МЕТОДЫ. Секвенирование фрагментов ДНК Секвенированием ДНК называют процесс определения последовательности нуклеотидов в ДНК. Метод секвенирования
- 45. МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИЕ МЕТОДЫ. Секвенирование фрагментов ДНК Метод Сэнгера (дидезокси-метод) Выделение ДНК Приготовление реакционной смеси -образец ДНК (матрица)
- 46. МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИЕ МЕТОДЫ. Секвенирование фрагментов ДНК После плавления ДНК и отжига праймера (праймер один), начинается синтез цепи.
- 48. Скачать презентацию
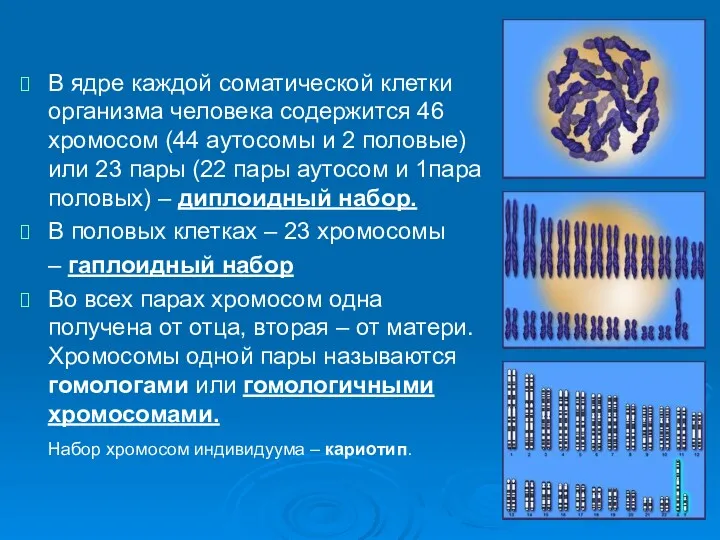
В ядре каждой соматической клетки организма человека содержится 46 хромосом (44

Строение
хромосом
Материал, из которого построены хромосомы – хроматин – представляет собой
Строение
хромосом
Материал, из которого построены хромосомы – хроматин – представляет собой
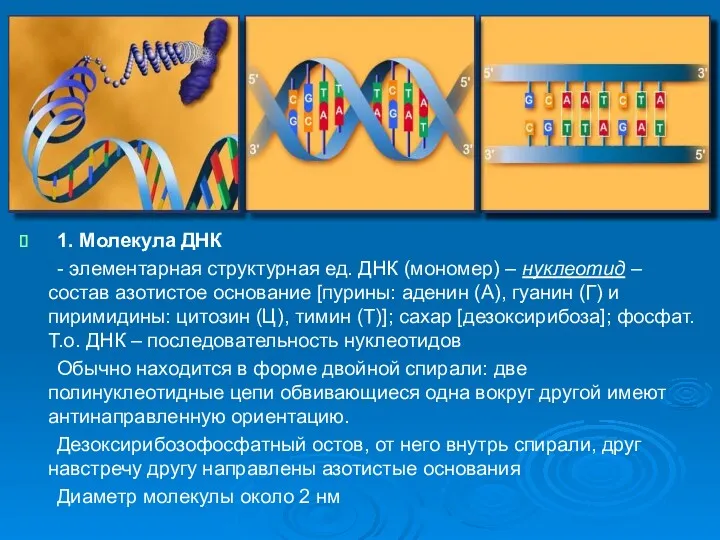
1. Молекула ДНК
- элементарная структурная ед. ДНК (мономер) – нуклеотид –
1. Молекула ДНК
- элементарная структурная ед. ДНК (мономер) – нуклеотид –
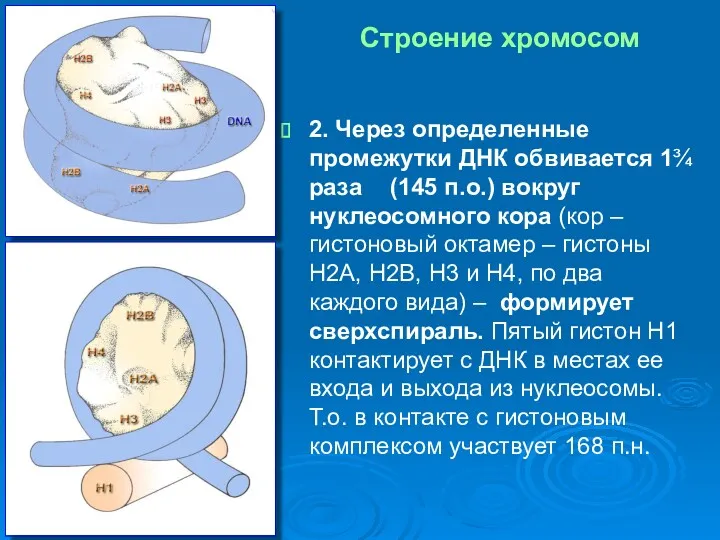
2. Через определенные промежутки ДНК обвивается 1¾ раза (145 п.о.) вокруг
2. Через определенные промежутки ДНК обвивается 1¾ раза (145 п.о.) вокруг
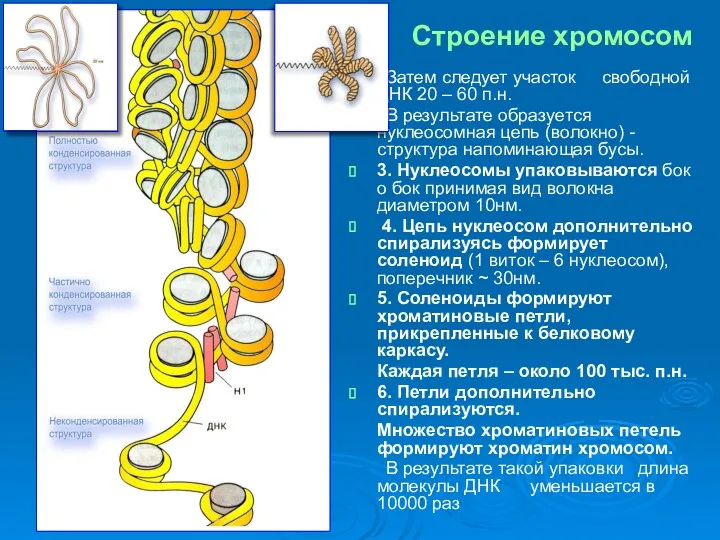
Затем следует участок свободной ДНК 20 – 60 п.н.
В результате образуется
Затем следует участок свободной ДНК 20 – 60 п.н.
В результате образуется

Центромера – компактный участок хромосомы, к которому при делении клетки прикрепляются
Центромера – компактный участок хромосомы, к которому при делении клетки прикрепляются
КЛАССИФИКАЦИЯ
По соотношению длин плеч можно выделить хромосомы:
Метацентрические
- длина p ≈
КЛАССИФИКАЦИЯ
По соотношению длин плеч можно выделить хромосомы:
Метацентрические
- длина p ≈
КЛАССИФИКАЦИЯ
Все хромосомы разделены на 7 групп, которые были обозначены буквами английского
КЛАССИФИКАЦИЯ Все хромосомы разделены на 7 групп, которые были обозначены буквами английского
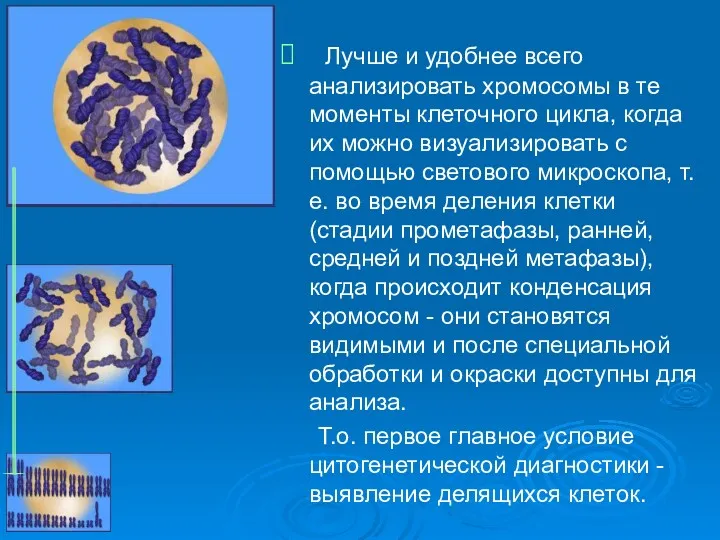
Лучше и удобнее всего анализировать хромосомы в те моменты клеточного
Лучше и удобнее всего анализировать хромосомы в те моменты клеточного

ОСНОВНЫЕ МЕТОДЫ ПРИГОТОВЛЕНИЯ ХРОМОСОМНЫХ ПРЕПАРАТОВ
В качестве материала для получения хромосом человека
ОСНОВНЫЕ МЕТОДЫ ПРИГОТОВЛЕНИЯ ХРОМОСОМНЫХ ПРЕПАРАТОВ
В качестве материала для получения хромосом человека

Основные методы приготовления хромосомных препаратов
ЭТАПЫ:
1) Забор материала – ткань, содержащая
Основные методы приготовления хромосомных препаратов
ЭТАПЫ:
1) Забор материала – ткань, содержащая

Культивирование клеток периферической крови, постановка культуры лимфоцитов человека.
Забор материала. Культивирование
Культивирование клеток периферической крови, постановка культуры лимфоцитов человека. Забор материала. Культивирование

Культивирование клеток периферической крови, постановка культуры лимфоцитов человека.
Остановка деления клеток на
Культивирование клеток периферической крови, постановка культуры лимфоцитов человека. Остановка деления клеток на

Приготовление хромосомных препаратов
Задачей процедур этого этапа является получение хорошо распластанных
Приготовление хромосомных препаратов
Задачей процедур этого этапа является получение хорошо распластанных
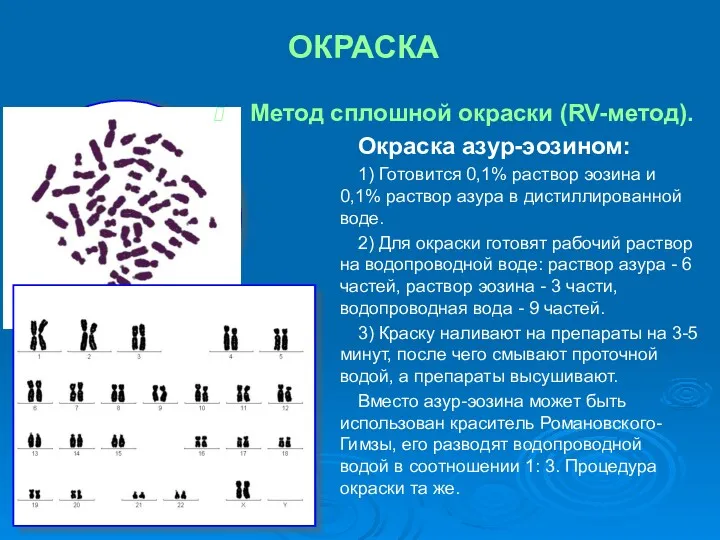
ОКРАСКА
Метод сплошной окраски (RV-метод).
Окраска азур-эозином:
1) Готовится 0,1% раствор эозина и
ОКРАСКА
Метод сплошной окраски (RV-метод).
Окраска азур-эозином:
1) Готовится 0,1% раствор эозина и

Методы дифференциальных окрасок хромосом, выявляющих их линейную неоднородность.
В практике широко применены
Методы дифференциальных окрасок хромосом, выявляющих их линейную неоднородность.
В практике широко применены
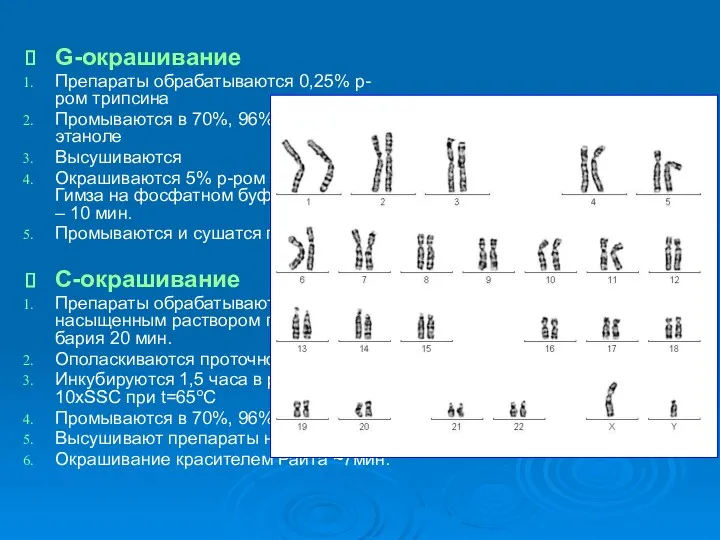
G-окрашивание
Препараты обрабатываются 0,25% р-ром трипсина
Промываются в 70%, 96%, 100% этаноле
Высушиваются
Окрашиваются
G-окрашивание
Препараты обрабатываются 0,25% р-ром трипсина
Промываются в 70%, 96%, 100% этаноле
Высушиваются
Окрашиваются

R-окрашивание
«Состарившиеся» хромосомные препараты инкубируют при t=60oC в насыщенном растворе гидроокиси бария
R-окрашивание
«Состарившиеся» хромосомные препараты инкубируют при t=60oC в насыщенном растворе гидроокиси бария
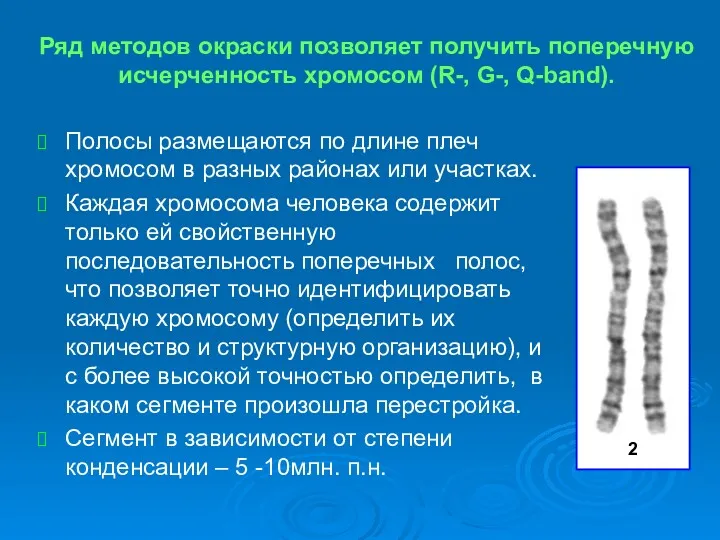
Ряд методов окраски позволяет получить поперечную исчерченность хромосом (R-, G-, Q-band).
Ряд методов окраски позволяет получить поперечную исчерченность хромосом (R-, G-, Q-band).

Полосы и участки нумеруются в направлении от центромеры к теломере по
Полосы и участки нумеруются в направлении от центромеры к теломере по

В любом обозначении хромосомы используются:
1) номер хромосомы,
2) символ плеча,
3)
В любом обозначении хромосомы используются:
1) номер хромосомы,
2) символ плеча,
3)
![Цитогенетическое исследование [G-band]](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/174994/slide-22.jpg)
Цитогенетическое исследование [G-band]
Цитогенетическое исследование [G-band]

Основные принципы анализа
хромосомных препаратов
Цитогенетический анализ хромосом проводят на препаратах
Основные принципы анализа
хромосомных препаратов
Цитогенетический анализ хромосом проводят на препаратах

Внедрение методов дифференциальной окраски хромосом способствовало значительному прогрессу в цитогенетике человека
Внедрение методов дифференциальной окраски хромосом способствовало значительному прогрессу в цитогенетике человека

МОЛЕКУЛЯРНО-ЦИТОГЕНЕТИЧЕСКОЕ ИССЛЕДОВАНИЕ
Базируется на технологии гибридизации ДНК.
Позволяет увидеть хромосомную локализацию специфических последовательностей
МОЛЕКУЛЯРНО-ЦИТОГЕНЕТИЧЕСКОЕ ИССЛЕДОВАНИЕ
Базируется на технологии гибридизации ДНК.
Позволяет увидеть хромосомную локализацию специфических последовательностей

МОЛЕКУЛЯРНО-ЦИТОГЕНЕТИЧЕСКОЕ ИССЛЕДОВАНИЕ
ЭТАПЫ:
1) 1) – 6) Этапы аналогичны этапам цитогенетического исследования
Денатурация образца
МОЛЕКУЛЯРНО-ЦИТОГЕНЕТИЧЕСКОЕ ИССЛЕДОВАНИЕ
ЭТАПЫ:
1) 1) – 6) Этапы аналогичны этапам цитогенетического исследования
Денатурация образца
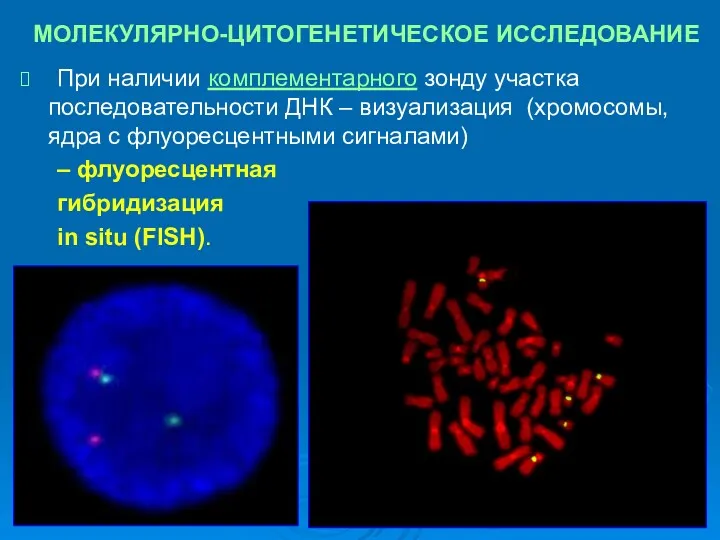
МОЛЕКУЛЯРНО-ЦИТОГЕНЕТИЧЕСКОЕ ИССЛЕДОВАНИЕ
При наличии комплементарного зонду участка последовательности ДНК – визуализация (хромосомы,
МОЛЕКУЛЯРНО-ЦИТОГЕНЕТИЧЕСКОЕ ИССЛЕДОВАНИЕ
При наличии комплементарного зонду участка последовательности ДНК – визуализация (хромосомы,
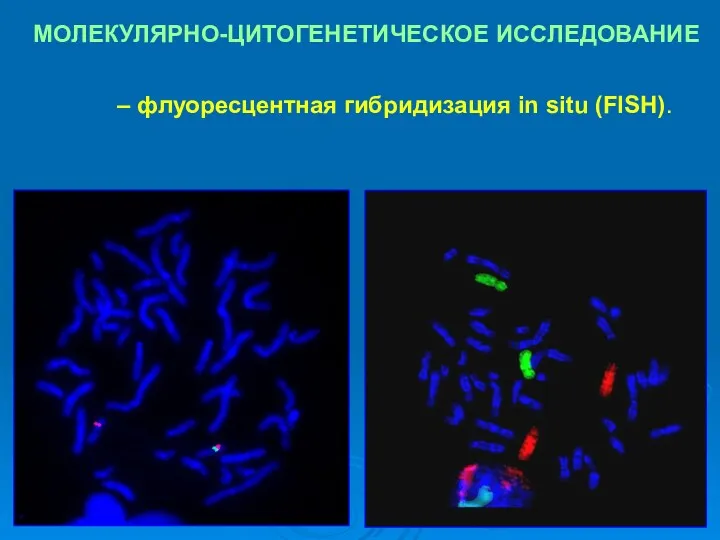
МОЛЕКУЛЯРНО-ЦИТОГЕНЕТИЧЕСКОЕ ИССЛЕДОВАНИЕ
– флуоресцентная гибридизация in situ (FISH).
МОЛЕКУЛЯРНО-ЦИТОГЕНЕТИЧЕСКОЕ ИССЛЕДОВАНИЕ
– флуоресцентная гибридизация in situ (FISH).

МОЛЕКУЛЯРНО-ЦИТОГЕНЕТИЧЕСКОЕ ИССЛЕДОВАНИЕ
Новые молекулярно-цитогенетические технологии – анализ многоцветно окрашенных хромосом – спектральное
МОЛЕКУЛЯРНО-ЦИТОГЕНЕТИЧЕСКОЕ ИССЛЕДОВАНИЕ
Новые молекулярно-цитогенетические технологии – анализ многоцветно окрашенных хромосом – спектральное
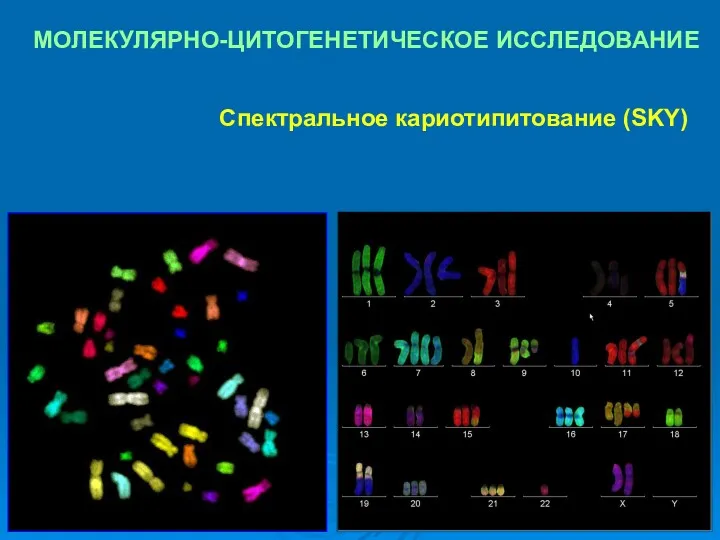
МОЛЕКУЛЯРНО-ЦИТОГЕНЕТИЧЕСКОЕ ИССЛЕДОВАНИЕ
Спектральное кариотипитование (SKY)
МОЛЕКУЛЯРНО-ЦИТОГЕНЕТИЧЕСКОЕ ИССЛЕДОВАНИЕ
Спектральное кариотипитование (SKY)
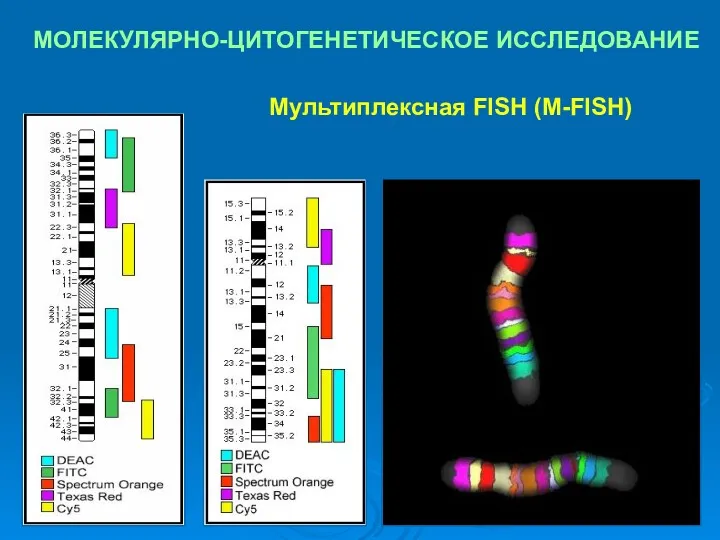
МОЛЕКУЛЯРНО-ЦИТОГЕНЕТИЧЕСКОЕ ИССЛЕДОВАНИЕ
Мультиплексная FISH (M-FISH)
МОЛЕКУЛЯРНО-ЦИТОГЕНЕТИЧЕСКОЕ ИССЛЕДОВАНИЕ
Мультиплексная FISH (M-FISH)
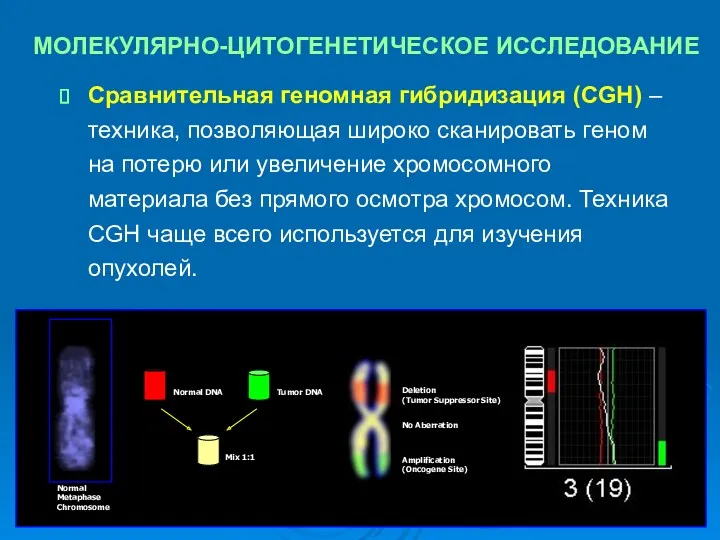
МОЛЕКУЛЯРНО-ЦИТОГЕНЕТИЧЕСКОЕ ИССЛЕДОВАНИЕ
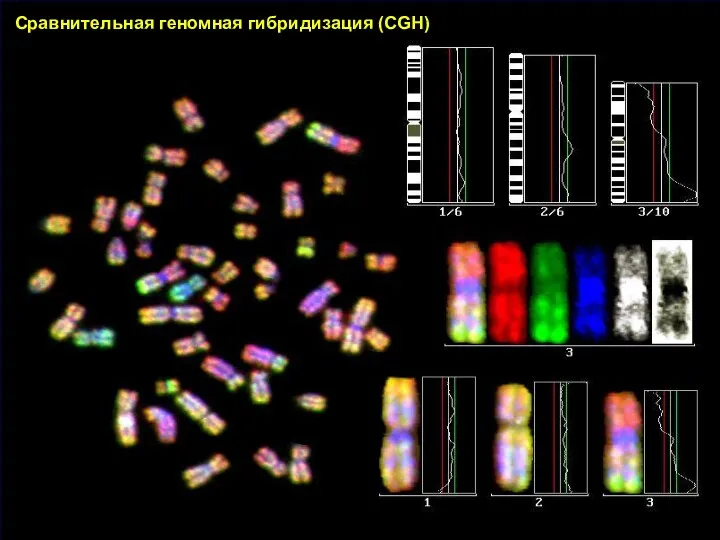
Сравнительная геномная гибридизация (CGH) – техника, позволяющая широко сканировать геном
МОЛЕКУЛЯРНО-ЦИТОГЕНЕТИЧЕСКОЕ ИССЛЕДОВАНИЕ
Сравнительная геномная гибридизация (CGH) – техника, позволяющая широко сканировать геном
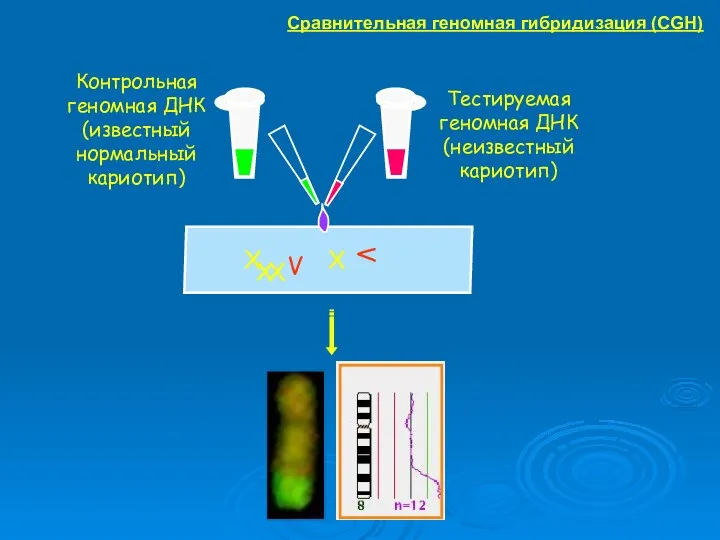
Контрольная геномная ДНК
(известный нормальный кариотип)
Тестируемая геномная ДНК
(неизвестный кариотип)
Сравнительная геномная гибридизация (CGH)
Контрольная геномная ДНК
(известный нормальный кариотип)
Тестируемая геномная ДНК
(неизвестный кариотип)
Сравнительная геномная гибридизация (CGH)
Сравнительная геномная гибридизация (CGH)
Сравнительная геномная гибридизация (CGH)

МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИЕ МЕТОДЫ:
ГЕНЕТИЧЕСКАЯ ИНЖЕНЕРИЯ (ТЕХНОЛОГИЯ РЕКОМБИНАНТНЫХ ДНК)
Методы клонирования ДНК
Методы анализа первичной последовательности
МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИЕ МЕТОДЫ:
ГЕНЕТИЧЕСКАЯ ИНЖЕНЕРИЯ (ТЕХНОЛОГИЯ РЕКОМБИНАНТНЫХ ДНК)
Методы клонирования ДНК
Методы анализа первичной последовательности

Клонирование ДНК
– позволяет из единичного фрагмента ДНК получить достаточно большое количество
Клонирование ДНК
– позволяет из единичного фрагмента ДНК получить достаточно большое количество

МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИЕ МЕТОДЫ:
Клонирование in vivo
Фрагмент ДНК, вырезанный с помощью рестриктаз, соединяется
МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИЕ МЕТОДЫ:
Клонирование in vivo
Фрагмент ДНК, вырезанный с помощью рестриктаз, соединяется

Клонирование вне клеток. Полимеразная цепная реакция (ПЦР)
Выделение ДНК
Приготовление реакционной смеси
Клонирование вне клеток. Полимеразная цепная реакция (ПЦР)
Выделение ДНК
Приготовление реакционной смеси

МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИЕ МЕТОДЫ.
Амплификация → увеличение количества повторов интересующего участка. Процесс –
МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИЕ МЕТОДЫ.
Амплификация → увеличение количества повторов интересующего участка. Процесс –
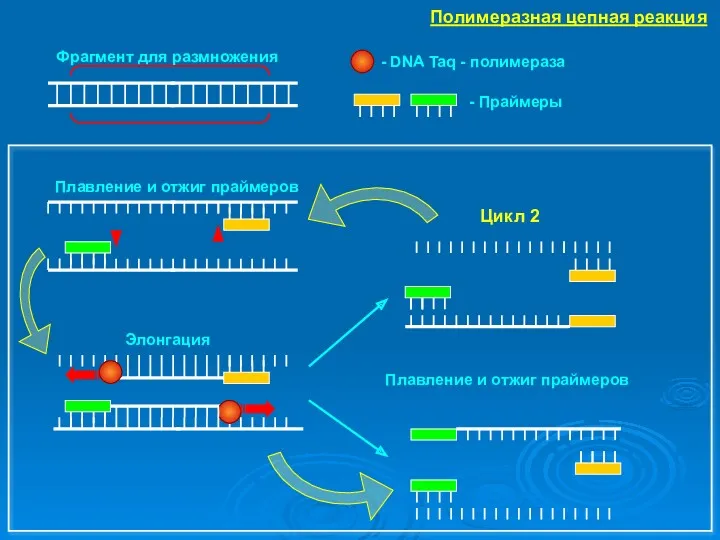
Фрагмент для размножения
Цикл 2
Плавление и отжиг праймеров
Элонгация
- DNA Taq - полимераза
Полимеразная
Фрагмент для размножения
Цикл 2
Плавление и отжиг праймеров
Элонгация
- DNA Taq - полимераза
Полимеразная
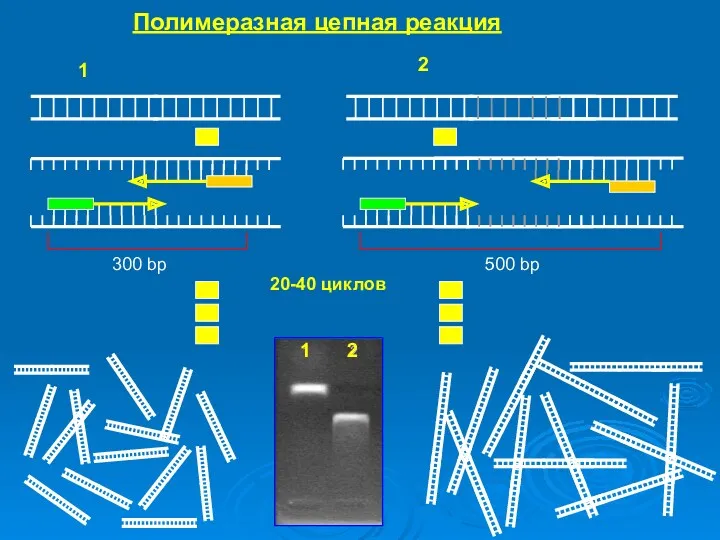
Полимеразная цепная реакция
300 bp
500 bp
20-40 циклов
1
2
1
2
Полимеразная цепная реакция
300 bp
500 bp
20-40 циклов
1
2
1
2
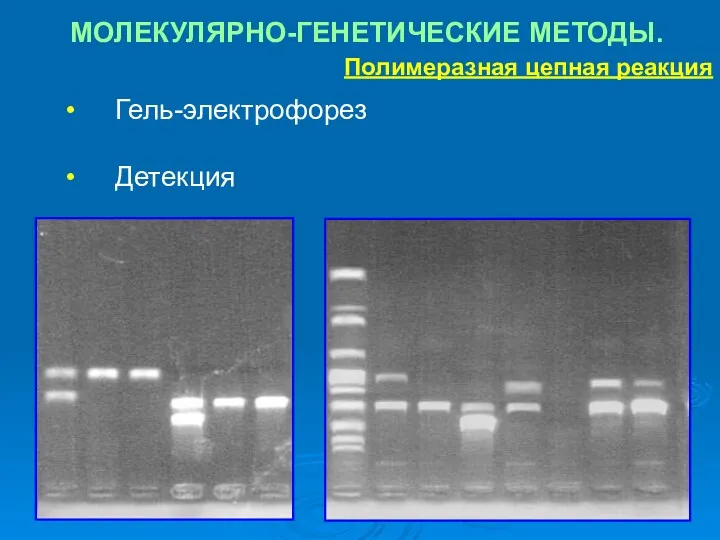
МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИЕ МЕТОДЫ.
Полимеразная цепная реакция
Гель-электрофорез
Детекция
МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИЕ МЕТОДЫ.
Полимеразная цепная реакция
Гель-электрофорез
Детекция

МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИЕ МЕТОДЫ. Секвенирование фрагментов ДНК
Секвенированием ДНК называют процесс определения последовательности нуклеотидов
МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИЕ МЕТОДЫ. Секвенирование фрагментов ДНК
Секвенированием ДНК называют процесс определения последовательности нуклеотидов

МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИЕ МЕТОДЫ. Секвенирование фрагментов ДНК
Метод Сэнгера (дидезокси-метод)
Выделение ДНК
Приготовление реакционной смеси
МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИЕ МЕТОДЫ. Секвенирование фрагментов ДНК
Метод Сэнгера (дидезокси-метод)
Выделение ДНК
Приготовление реакционной смеси

МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИЕ МЕТОДЫ. Секвенирование фрагментов ДНК
После плавления ДНК и отжига праймера
МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИЕ МЕТОДЫ. Секвенирование фрагментов ДНК
После плавления ДНК и отжига праймера
 Теории возникновения жизни
Теории возникновения жизни Высшие растения
Высшие растения Презнтация Электрические станции на службе человека.
Презнтация Электрические станции на службе человека. Культивирование бактерий. Изучение культуральных свойств
Культивирование бактерий. Изучение культуральных свойств Animals. Food chain
Animals. Food chain Побег, его строение и развитие
Побег, его строение и развитие Биологические мембраны. Структура, свойства и пути их изучения
Биологические мембраны. Структура, свойства и пути их изучения Щитовидная, паращитовидная, поджелудочная железы, надпочечники
Щитовидная, паращитовидная, поджелудочная железы, надпочечники Изучение строения скелета человека
Изучение строения скелета человека Презентация Карлики и великаны в животном мире
Презентация Карлики и великаны в животном мире Тірі ағзалардың көптүрлілігі
Тірі ағзалардың көптүрлілігі Проводящие пути: двигательные. Чувствительные
Проводящие пути: двигательные. Чувствительные Влияние абиотических факторов на выгонку луковичных растений к определенному времени
Влияние абиотических факторов на выгонку луковичных растений к определенному времени Строение крови и ее функции
Строение крови и ее функции Тип членистоногие, класс насекомые
Тип членистоногие, класс насекомые Промежуточный мозг
Промежуточный мозг Обмін БЖВ
Обмін БЖВ Презентация к уроку: Клетки крови.
Презентация к уроку: Клетки крови. Презенетация к уроку Строение цветка. Типы соцветий
Презенетация к уроку Строение цветка. Типы соцветий Старение, как заключительный этап онтогенеза
Старение, как заключительный этап онтогенеза Закономерности проведения информации. Соматосенсорная система
Закономерности проведения информации. Соматосенсорная система Простейшие, или эукариоты. (5 класс)
Простейшие, или эукариоты. (5 класс) White tigers
White tigers Открытый урок по темеЛист. Внешнее и внутреннее строение
Открытый урок по темеЛист. Внешнее и внутреннее строение Болезни с нетрадиционным наследованием
Болезни с нетрадиционным наследованием портфолио учителя биологии Канунниковой Л.Н.
портфолио учителя биологии Канунниковой Л.Н. Мой домашний питомец
Мой домашний питомец Теория антропосоциогенеза
Теория антропосоциогенеза