- Молекулярные основы наследственности. Биосинтез белка. Транскрипция

Содержание
- 2. Процессинг РНК как компонент центральной догмы молекулярной биологии ДНК пре-РНК РНК БЕЛОК Репликация рекомбинация репарация репликация
- 3. 1. Особенности транскрипции.
- 4. Так выглядит транскрипция – синтез РНК для последующего производства белка http://www.vechnayamolodost.ru/pages/drugienaukiozhizni/rodkoodnrntrimndr80.html
- 5. Фермент РНК-полимераза (зеленые комки) ползет по молекуле ДНК (скрученный тяж) и «считывает» ее, синтезируя молекулу РНК
- 6. Транскрипция – биосинтез одноцепочечной молекулы РНК на матрице ДНК 1. Синтез молекул РНК идет в направлении
- 7. Для осуществления транскрипции необходимо наличие: 1. ДНК-матрицы; 2. Пула предшественников (аденин-, гуанин-, цитозин- и урацилтрифосфатнуклеотиды), 3.
- 8. Последовательность ДНК, транскрибируемая в одну молекулу РНК, начинающаяся промотором и заканчивающаяся терминатором, называется транскрипционной единицей, или
- 9. Транскрипция – матричный процесс, в котором выделяют три стадии
- 10. 2. Транскрипция у прокариот. Схема работы лактозного оперона.
- 11. Схема регуляции транскрипции у прокариот (гипотеза оперона) была предложена Ф. Жакобом и Ж. Моно в 1961
- 12. В структуре РНК-полимеразы присутствует канавка длиной 55 Å (5,5 нм) и шириной 25 Å (2,5 нм).
- 13. Бактериальная РНК-полимераза состоит из нескольких белковых-субъединиц: двух α-субъединиц (это маленькие субъединицы), β- и β΄-субъединиц (большие субъединицы)
- 15. Инициация транскрипции. Структура промоторов Сочетание последовательностей "–10«(ТАТА- или Прибнов-бокс) и "–35" назвали классическим промотором, т.к. она
- 16. Инициация транскрипции После того, как σ-субъединица в составе кор-фермента связалась с промотором, ДНК на этом участке
- 17. РНК наращивается на 3΄-конце. Присоединением каждого нуклеотида кор-фермент делает шаг по ДНК и сдвигается на один
- 18. После того, как σ-фактор диссоциирует с элонгирующей РНК-полимеразой, с ней начинают взаимодействовать несколько белков – факторов
- 19. Белки GreA и GreB узнают так называемые «арестованные» комплексы. Эти комплексы образуются если РНК-полимераза сдвигается против
- 21. Регуляция транскрипции у прокариот В состав оперона входит: Промотор Инициатор Ген-оператор Структурные гены Терминатор Ген-регулятор не
- 22. Схема работы лактозного оперона Оперон работает
- 23. Регуляция работы генов у прокариот http://bio.fizteh.ru/student/files/biology/biolections/lection11.html
- 24. 3. Транскрипция у эукариот.
- 25. Схема регуляции транскрипции у эукариот разработана Георгием Павловичем Георгиевым (1972 г.) и получила название гипотезы транскриптона.
- 26. Транскрипция у эукариот Транскриптон состоит из неинформативной (акцепторной) и информативной (структурной) зон. Неинформативная зона начинается промотором
- 27. http://rpp.nashaucheba.ru/docs/index-137904.html
- 28. Энхансеры Энхансер (усилители транскрипции, англ. enhance — увеличивать, усиливать) – это генетический элемент, обладающий усиливающим транскрипцию
- 29. Сайленсеры Сайленсер (ослабители транскрипции, англ. silense — заглушать) – это регуляторный участок ДНК, который подавляет активность
- 30. Инсуляторы Специфичность действия энхансеров и сайленсеров определяется инсуляторами, которые блокируют активность энхансера или сайленсера.
- 31. Биосинтез белка. Трансляция.
- 32. Биосинтез белков Наиболее сложный из генетических процессов; Наиболее энергоемкий процесс; Протекает с высокой скоростью (при 37˚
- 33. Строение рибосомы http://biouroki.ru/material/plants/kletka.html 1 — большая субъединица; 2 — малая субъединица.
- 34. Рибозимы — катализаторы древнего мира Уникальное свойства ферментов рибозимов — их способность к низкотемпературному катализу —
- 35. Рибосома: химически – рибонуклеопротеид, физически – компактная частица, диаметром около 30 нм, функционально – молекулярная машина,
- 36. А-сайт – место связывания очередной аа-тРНК. Р-сайт. В начале процесса трансляции с пептидильным центром связывается инициирующая
- 37. Полирибосомы (полисомы) – синтезирующие белок внутриклеточные комплексы, каждый из которых состоит из молекулы мРНК и многих
- 38. б). эпицикл трансляции: инициация, элонгация, терминация
- 39. Синтез белков молекулой РНК http://nnm.ru/blogs/rab44/
- 40. Трансляция Трансляция – матричный процесс, включающий в себя определенные стадии в определенной последовательности (инициация, элонгация, терминация).
- 41. Инициация трансляции Это серия молекулярных событий, происходящих с рибосомой, которая приводит к взаимодействию рибосомы с началом
- 42. У эукариот инициирующий (стартовый) кодон – кодон аминокислоты метионина – АУГ. У прокариот (а также в
- 43. Аминокислоты тРНК метионина может присоединять в результате ферментативной реакции формильную группу к аминокислоте: Такая формилметиониновая тРНК
- 44. Схема демонстрирующая участие двух рибосомных субчастиц — малой (желтая) и большой (красная) — в процессе инициации
- 45. В рибосоме имеются три различных участка, с которыми связываются РНК, - один для иРНК и два
- 46. Элонгация полипептидной цепи на рибосомах слагается из трех отдельных этапов На первом этапе молекула аминоацил-тРНК связывается
- 47. Элонгационный цикл составляют три последовательных шага: связывание аминоацил-тРНК, транспептидация и транслокация.
- 48. Элонгация
- 49. Схема элонгации пептида на рибосоме: а — инициаторная аминоацил-тРНК находится в P-участке и первая элонгаторная аминоацил-тРНК
- 50. Стадии трансляции: терминация Схема терминации трансляции: а — после добавления последнего аминокислотного остатка к растущему полипептиду,
- 51. Терминация — узнавание терминирующего кодона (стоп-кодона) и отделение продукта. Из 64 возможных кодонов мРНК три: УАА,
- 52. Свойства генетического кода Основное свойство генетического кода — его триплетность. Каждая аминокислота кодируется тройкой нуклеотидов. Избыточность
- 54. Скачать презентацию
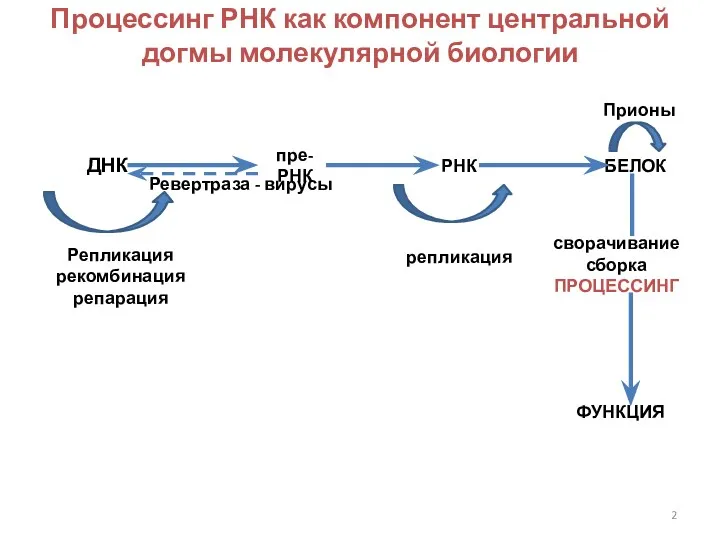
Процессинг РНК как компонент центральной догмы молекулярной биологии
ДНК
пре-РНК
РНК
БЕЛОК
Репликация
рекомбинация
репарация
репликация
сворачивание
сборка
ПРОЦЕССИНГ
ФУНКЦИЯ
Прионы
Ревертраза - вирусы
Процессинг РНК как компонент центральной догмы молекулярной биологии
ДНК
пре-РНК
РНК
БЕЛОК
Репликация
рекомбинация
репарация
репликация
сворачивание
сборка
ПРОЦЕССИНГ
ФУНКЦИЯ
Прионы
Ревертраза - вирусы

1. Особенности транскрипции.
1. Особенности транскрипции.
Так выглядит транскрипция – синтез РНК для последующего производства белка
http://www.vechnayamolodost.ru/pages/drugienaukiozhizni/rodkoodnrntrimndr80.html
Так выглядит транскрипция – синтез РНК для последующего производства белка
http://www.vechnayamolodost.ru/pages/drugienaukiozhizni/rodkoodnrntrimndr80.html
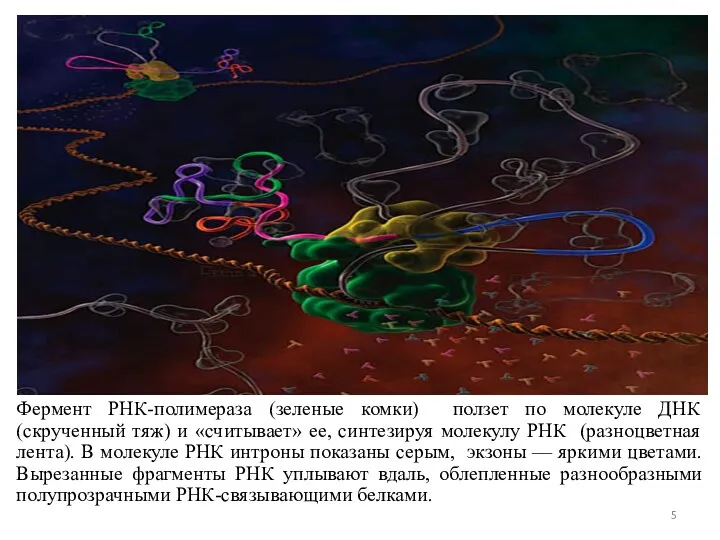
Фермент РНК-полимераза (зеленые комки) ползет по молекуле ДНК (скрученный тяж) и
Фермент РНК-полимераза (зеленые комки) ползет по молекуле ДНК (скрученный тяж) и
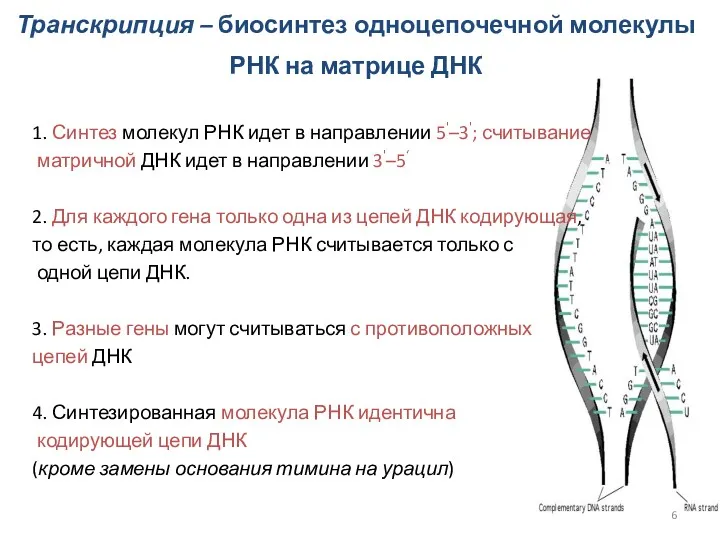
Транскрипция – биосинтез одноцепочечной молекулы РНК на матрице ДНК
1.
Транскрипция – биосинтез одноцепочечной молекулы РНК на матрице ДНК
1.
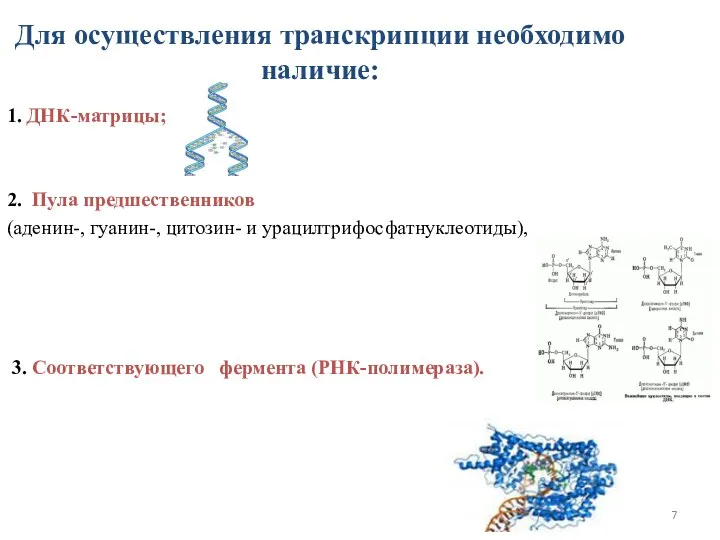
Для осуществления транскрипции необходимо наличие:
1. ДНК-матрицы;
2. Пула предшественников
(аденин-, гуанин-, цитозин- и
Для осуществления транскрипции необходимо наличие:
1. ДНК-матрицы;
2. Пула предшественников
(аденин-, гуанин-, цитозин- и
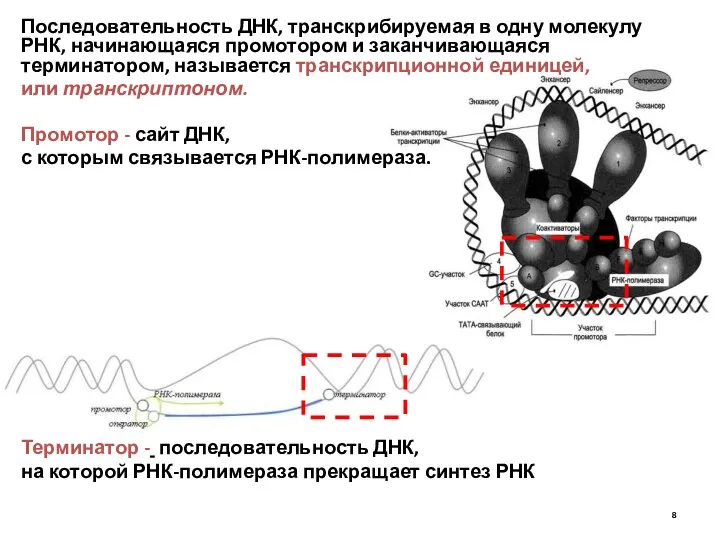
Последовательность ДНК, транскрибируемая в одну молекулу РНК, начинающаяся промотором и заканчивающаяся
Последовательность ДНК, транскрибируемая в одну молекулу РНК, начинающаяся промотором и заканчивающаяся

Транскрипция – матричный процесс, в котором выделяют три стадии
Транскрипция – матричный процесс, в котором выделяют три стадии

2. Транскрипция у прокариот. Схема работы лактозного оперона.
2. Транскрипция у прокариот. Схема работы лактозного оперона.

Схема регуляции транскрипции у прокариот (гипотеза оперона) была предложена Ф.
Схема регуляции транскрипции у прокариот (гипотеза оперона) была предложена Ф.

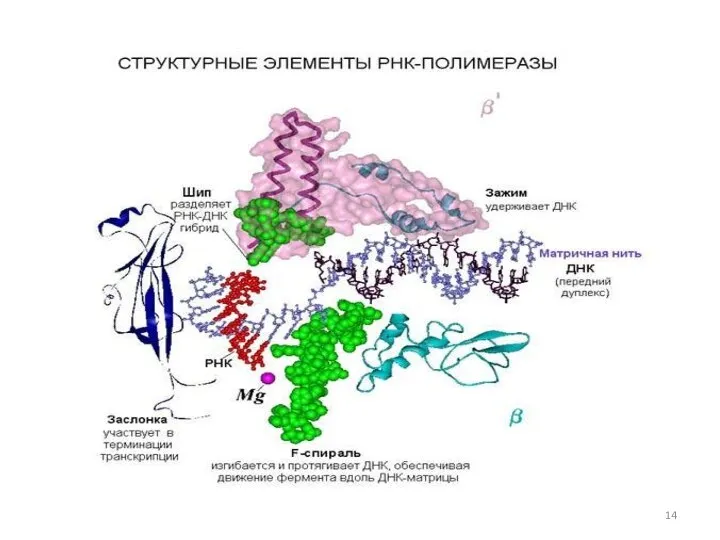
В структуре РНК-полимеразы присутствует канавка длиной 55 Å (5,5 нм) и
В структуре РНК-полимеразы присутствует канавка длиной 55 Å (5,5 нм) и

Бактериальная РНК-полимераза состоит из нескольких белковых-субъединиц: двух α-субъединиц (это маленькие субъединицы),
Бактериальная РНК-полимераза состоит из нескольких белковых-субъединиц: двух α-субъединиц (это маленькие субъединицы),
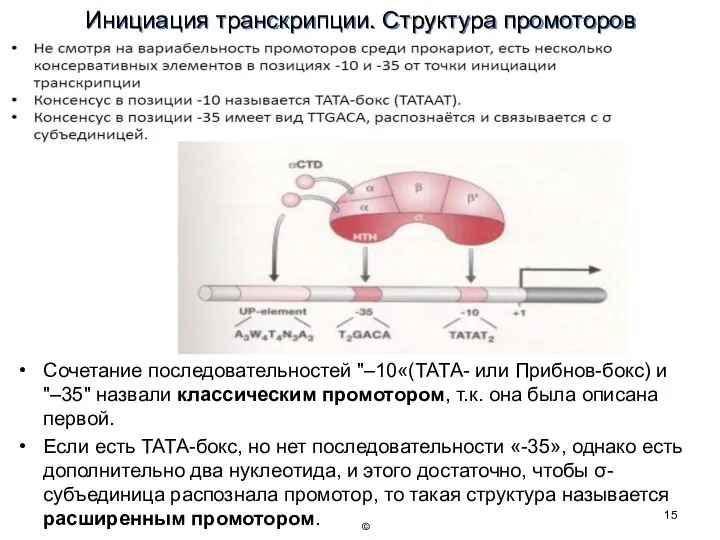
Инициация транскрипции. Структура промоторов
Сочетание последовательностей "–10«(ТАТА- или Прибнов-бокс) и "–35" назвали
Инициация транскрипции. Структура промоторов
Сочетание последовательностей "–10«(ТАТА- или Прибнов-бокс) и "–35" назвали
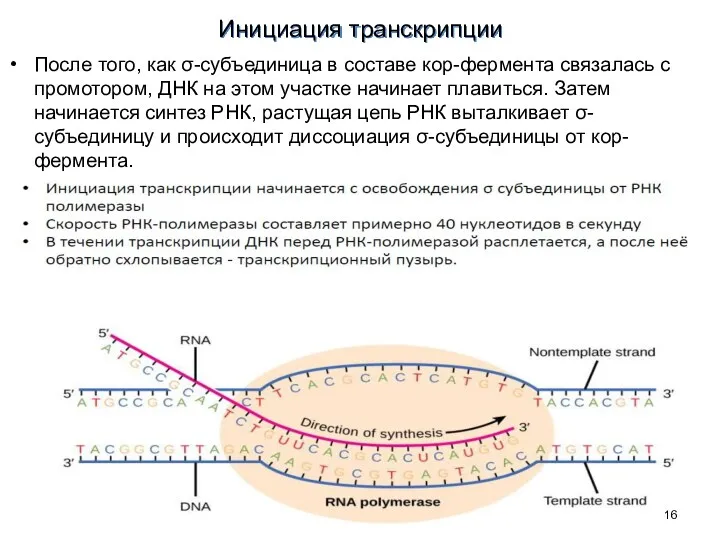
Инициация транскрипции
После того, как σ-субъединица в составе кор-фермента связалась с промотором,
Инициация транскрипции
После того, как σ-субъединица в составе кор-фермента связалась с промотором,
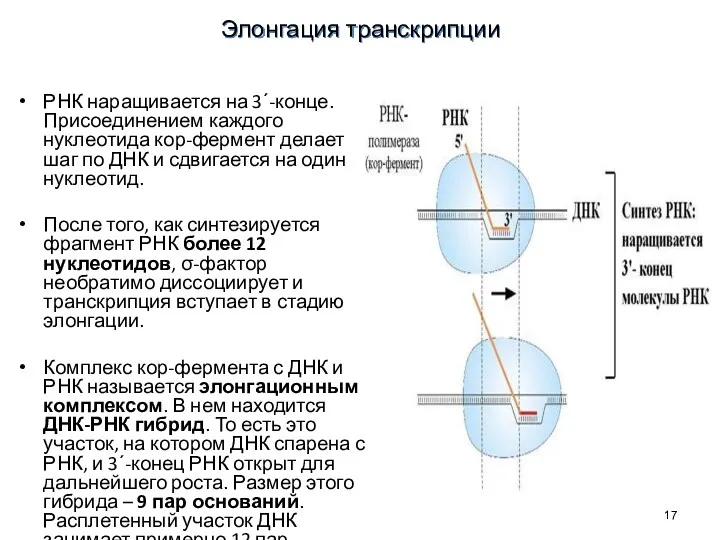
РНК наращивается на 3΄-конце. Присоединением каждого нуклеотида кор-фермент делает шаг по
РНК наращивается на 3΄-конце. Присоединением каждого нуклеотида кор-фермент делает шаг по
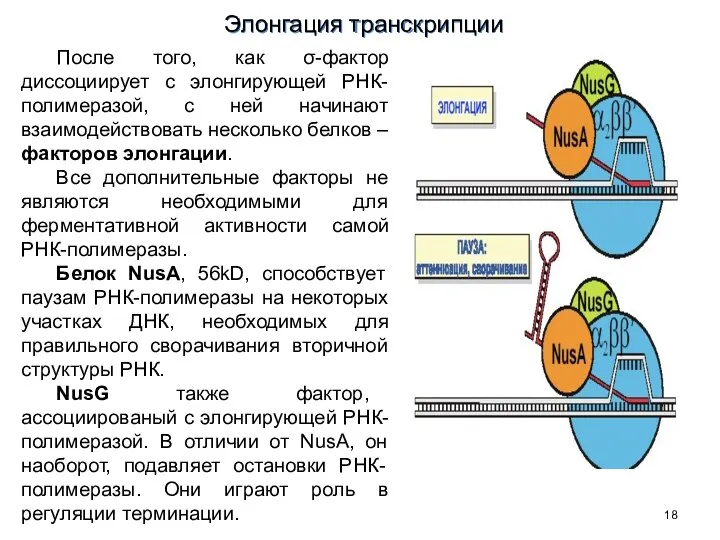
После того, как σ-фактор диссоциирует с элонгирующей РНК-полимеразой, с ней начинают
После того, как σ-фактор диссоциирует с элонгирующей РНК-полимеразой, с ней начинают
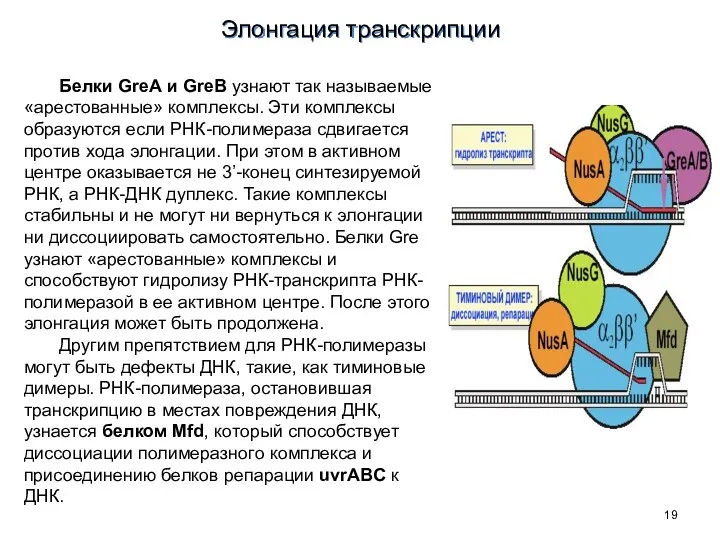
Белки GreA и GreB узнают так называемые «арестованные» комплексы. Эти комплексы
Белки GreA и GreB узнают так называемые «арестованные» комплексы. Эти комплексы


Регуляция транскрипции у прокариот
В состав оперона входит:
Промотор
Инициатор
Ген-оператор
Структурные гены
Регуляция транскрипции у прокариот
В состав оперона входит:
Промотор
Инициатор
Ген-оператор
Структурные гены
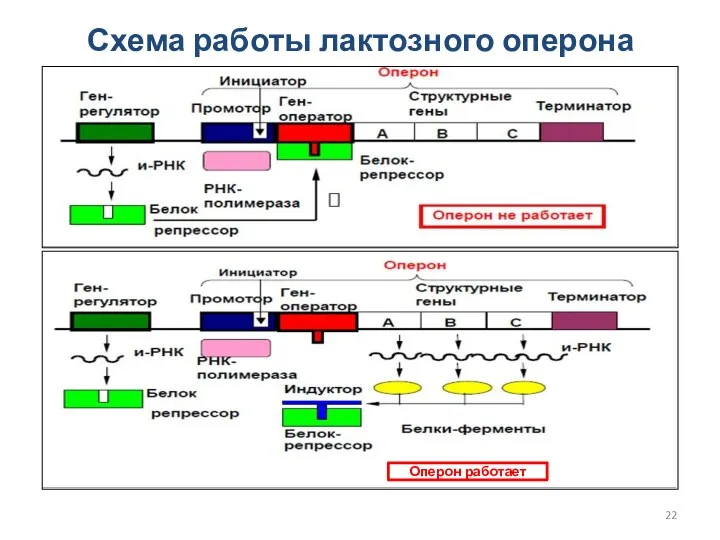
Схема работы лактозного оперона
Оперон работает
Схема работы лактозного оперона
Оперон работает
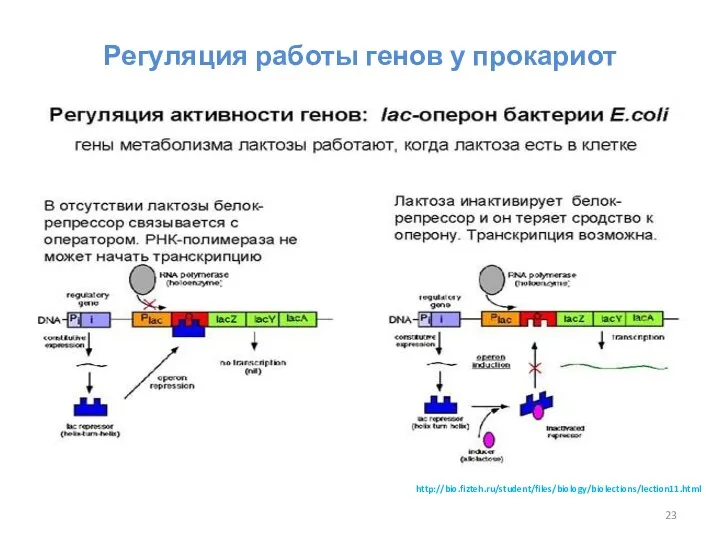
Регуляция работы генов у прокариот
http://bio.fizteh.ru/student/files/biology/biolections/lection11.html
Регуляция работы генов у прокариот
http://bio.fizteh.ru/student/files/biology/biolections/lection11.html

3. Транскрипция у эукариот.
3. Транскрипция у эукариот.

Схема регуляции транскрипции у эукариот разработана Георгием Павловичем Георгиевым (1972 г.)
Схема регуляции транскрипции у эукариот разработана Георгием Павловичем Георгиевым (1972 г.)
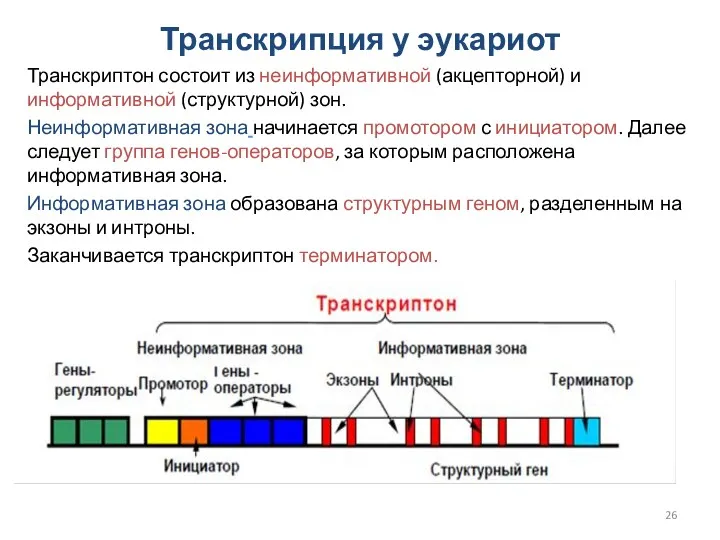
Транскрипция у эукариот
Транскриптон состоит из неинформативной (акцепторной) и информативной (структурной) зон.
Неинформативная
Транскрипция у эукариот
Транскриптон состоит из неинформативной (акцепторной) и информативной (структурной) зон.
Неинформативная
http://rpp.nashaucheba.ru/docs/index-137904.html
http://rpp.nashaucheba.ru/docs/index-137904.html
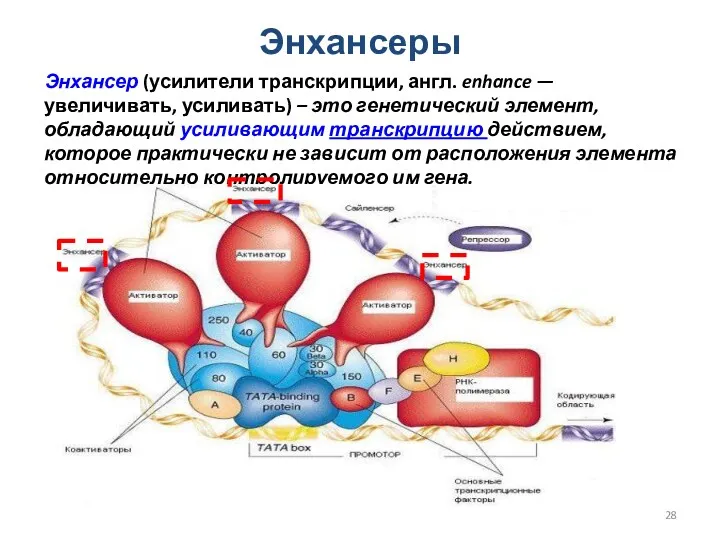
Энхансеры
Энхансер (усилители транскрипции, англ. enhance — увеличивать, усиливать) – это генетический
Энхансеры
Энхансер (усилители транскрипции, англ. enhance — увеличивать, усиливать) – это генетический
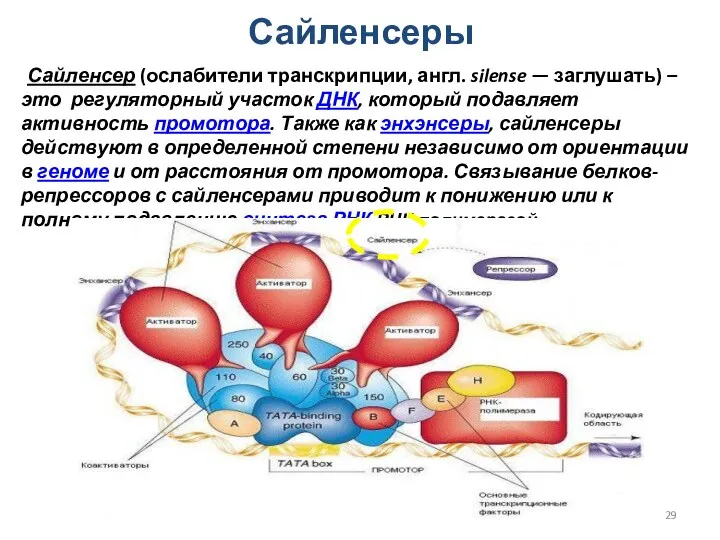
Сайленсеры
Сайленсер (ослабители транскрипции, англ. silense — заглушать) – это регуляторный
Сайленсеры
Сайленсер (ослабители транскрипции, англ. silense — заглушать) – это регуляторный
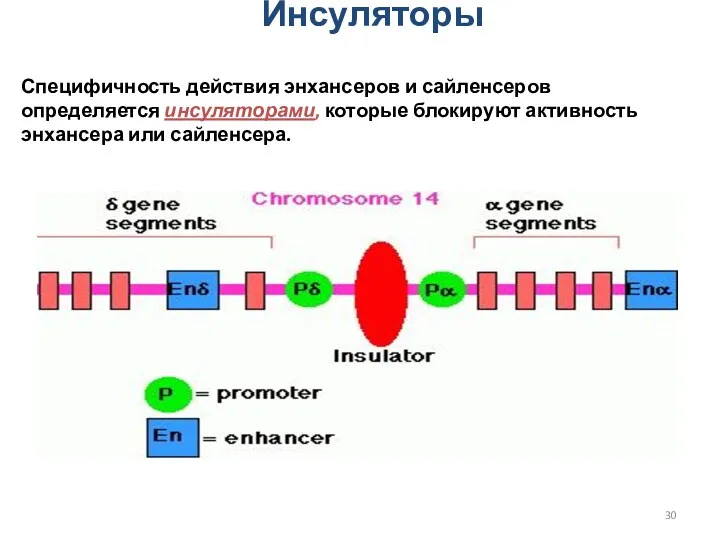
Инсуляторы
Специфичность действия энхансеров и сайленсеров определяется инсуляторами, которые блокируют активность
Инсуляторы
Специфичность действия энхансеров и сайленсеров определяется инсуляторами, которые блокируют активность
Биосинтез белка. Трансляция.
Биосинтез белка. Трансляция.

Биосинтез белков
Наиболее сложный из генетических процессов;
Наиболее энергоемкий процесс;
Протекает с
Биосинтез белков
Наиболее сложный из генетических процессов;
Наиболее энергоемкий процесс;
Протекает с
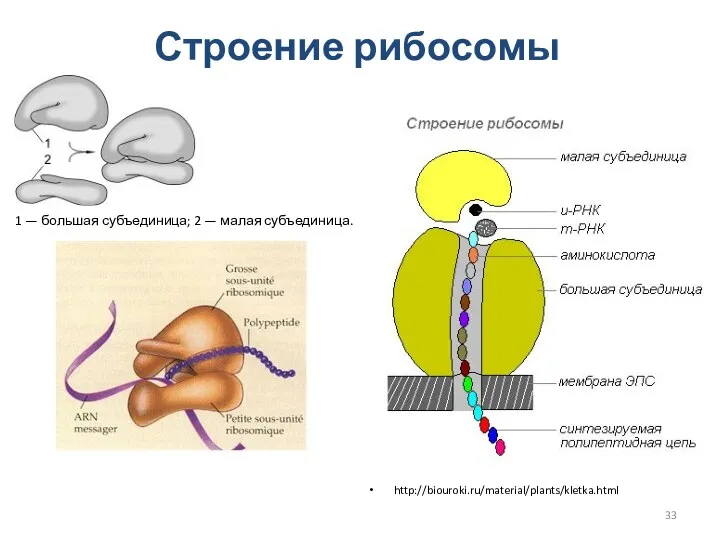
Строение рибосомы
http://biouroki.ru/material/plants/kletka.html
1 — большая субъединица; 2 — малая субъединица.
Строение рибосомы
http://biouroki.ru/material/plants/kletka.html
1 — большая субъединица; 2 — малая субъединица.
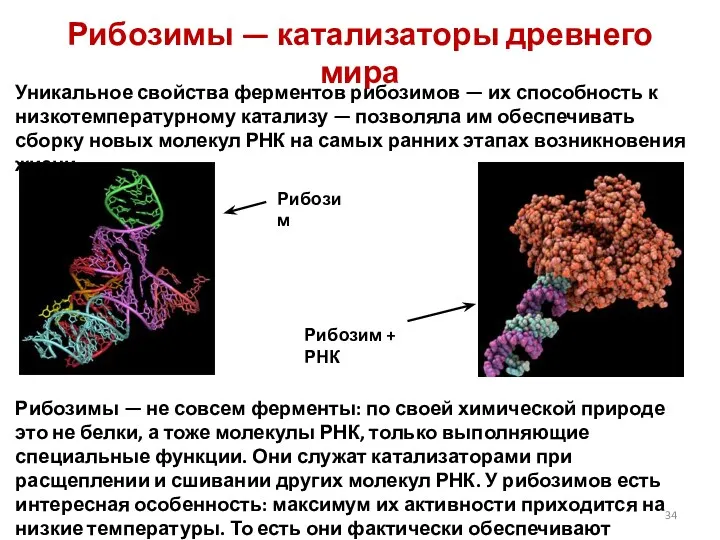
Рибозимы — катализаторы древнего мира
Уникальное свойства ферментов рибозимов — их способность
Рибозимы — катализаторы древнего мира
Уникальное свойства ферментов рибозимов — их способность

Рибосома:
химически – рибонуклеопротеид,
физически – компактная частица, диаметром около 30 нм,
функционально
Рибосома:
химически – рибонуклеопротеид,
физически – компактная частица, диаметром около 30 нм,
функционально
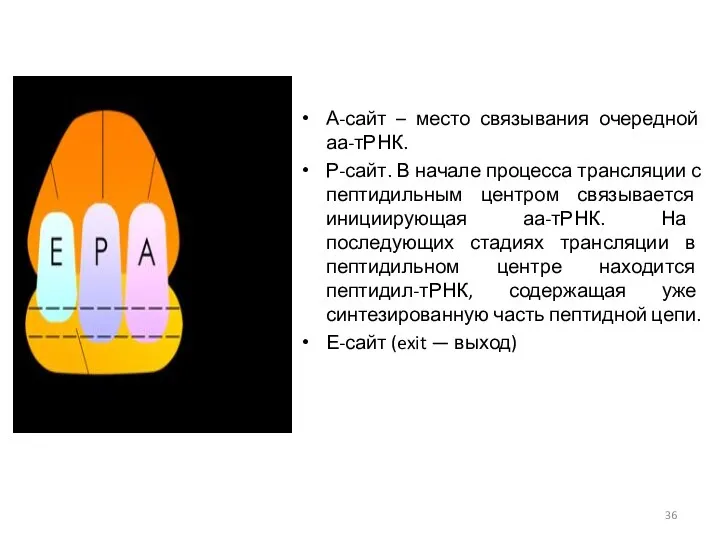
А-сайт – место связывания очередной аа-тРНК.
Р-сайт. В начале процесса трансляции с
А-сайт – место связывания очередной аа-тРНК.
Р-сайт. В начале процесса трансляции с
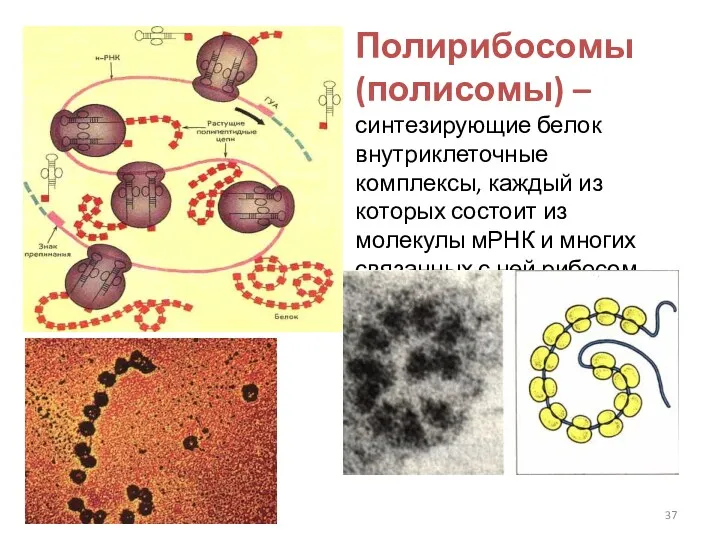
Полирибосомы (полисомы) – синтезирующие белок внутриклеточные комплексы, каждый из которых состоит
Полирибосомы (полисомы) – синтезирующие белок внутриклеточные комплексы, каждый из которых состоит

б). эпицикл трансляции:
инициация, элонгация, терминация
б). эпицикл трансляции:
инициация, элонгация, терминация

Синтез белков молекулой РНК
http://nnm.ru/blogs/rab44/
Синтез белков молекулой РНК
http://nnm.ru/blogs/rab44/
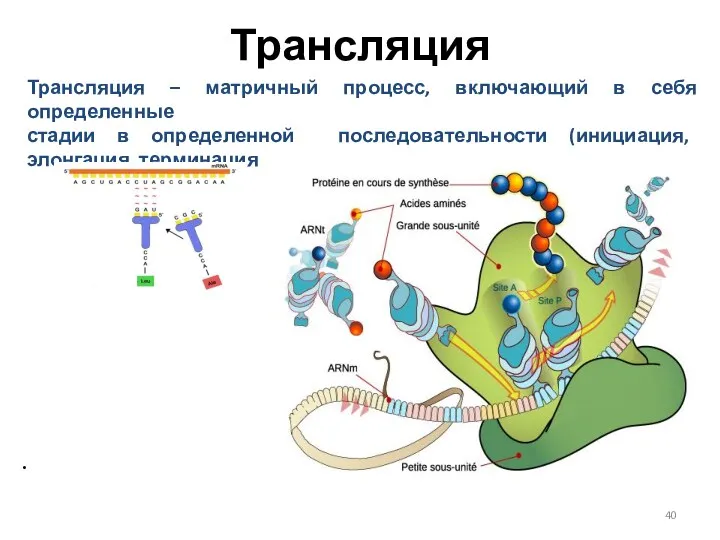
Трансляция
Трансляция – матричный процесс, включающий в себя определенные
стадии в определенной
Трансляция
Трансляция – матричный процесс, включающий в себя определенные
стадии в определенной

Инициация трансляции
Это серия молекулярных событий, происходящих с рибосомой, которая приводит к
Инициация трансляции
Это серия молекулярных событий, происходящих с рибосомой, которая приводит к

У эукариот инициирующий (стартовый) кодон – кодон аминокислоты метионина –
У эукариот инициирующий (стартовый) кодон – кодон аминокислоты метионина –
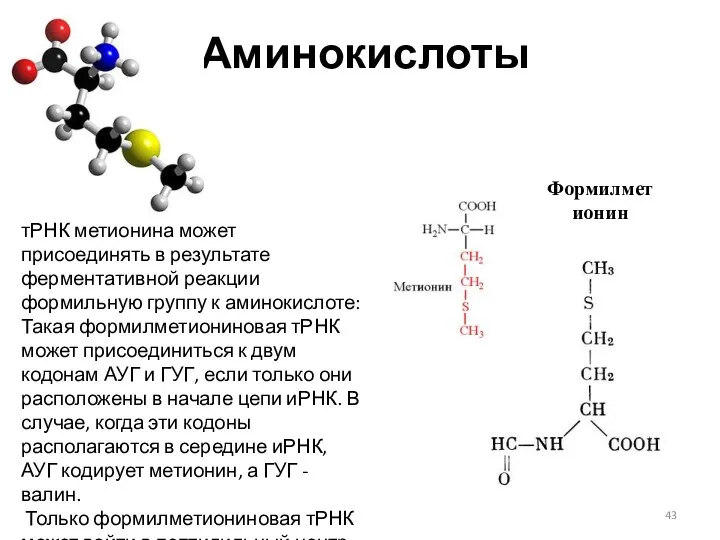
Аминокислоты
тРНК метионина может присоединять в результате ферментативной реакции формильную группу к
Аминокислоты
тРНК метионина может присоединять в результате ферментативной реакции формильную группу к
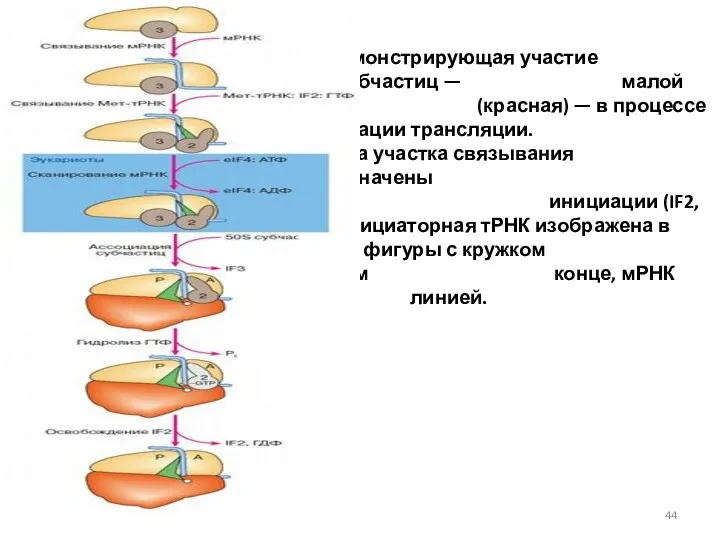
Схема демонстрирующая участие двух рибосомных субчастиц — малой (желтая) и
Схема демонстрирующая участие двух рибосомных субчастиц — малой (желтая) и
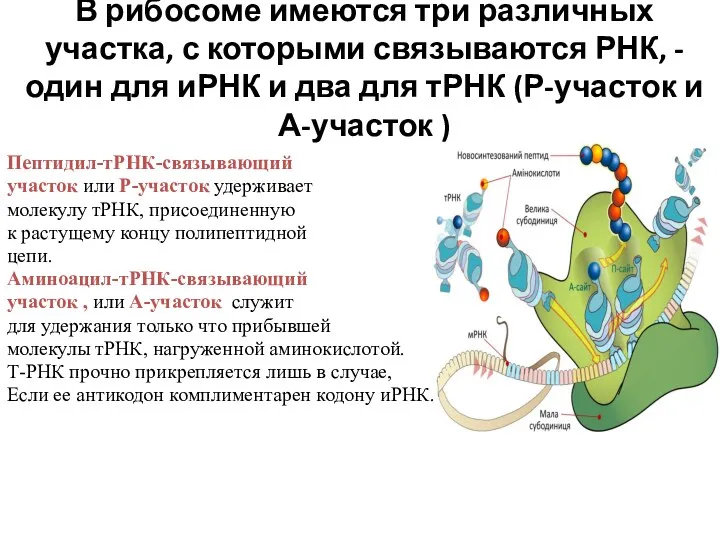
В рибосоме имеются три различных участка, с которыми связываются РНК, -
В рибосоме имеются три различных участка, с которыми связываются РНК, -

Элонгация полипептидной цепи на рибосомах слагается из трех отдельных этапов
На первом
Элонгация полипептидной цепи на рибосомах слагается из трех отдельных этапов
На первом

Элонгационный цикл составляют три последовательных шага: связывание аминоацил-тРНК, транспептидация и транслокация.
Элонгационный цикл составляют три последовательных шага: связывание аминоацил-тРНК, транспептидация и транслокация.
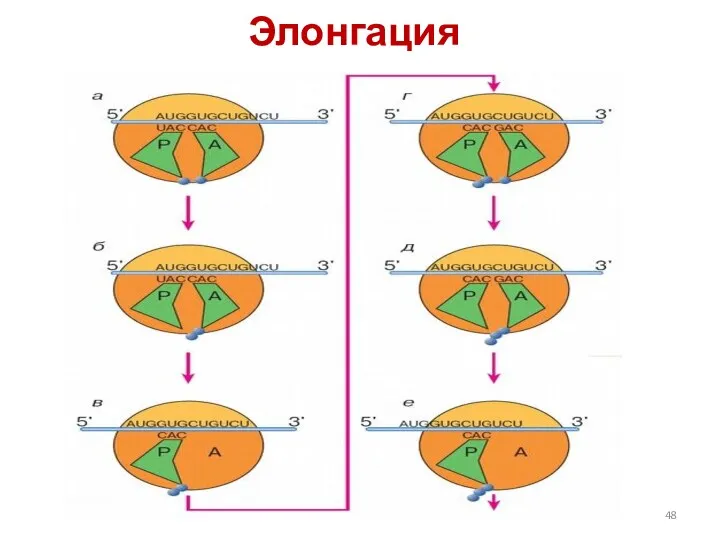
Элонгация
Элонгация

Схема элонгации пептида на рибосоме:
а — инициаторная аминоацил-тРНК находится в
Схема элонгации пептида на рибосоме:
а — инициаторная аминоацил-тРНК находится в
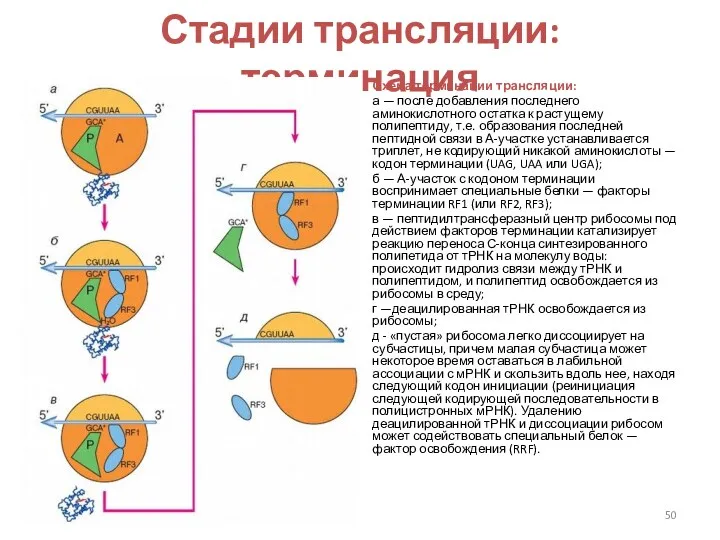
Стадии трансляции: терминация
Схема терминации трансляции:
а — после добавления последнего аминокислотного
Стадии трансляции: терминация
Схема терминации трансляции:
а — после добавления последнего аминокислотного

Терминация — узнавание терминирующего кодона (стоп-кодона) и отделение продукта.
Из 64 возможных
Терминация — узнавание терминирующего кодона (стоп-кодона) и отделение продукта.
Из 64 возможных

Свойства генетического кода
Основное свойство генетического кода — его триплетность. Каждая аминокислота
Свойства генетического кода
Основное свойство генетического кода — его триплетность. Каждая аминокислота
 Насекомое. Фото
Насекомое. Фото Класс Двудольные растения. Семейства Крестоцветные и Розоцветные
Класс Двудольные растения. Семейства Крестоцветные и Розоцветные Транскрипция РНК
Транскрипция РНК Семя, его строение и условия прорастания семян
Семя, его строение и условия прорастания семян Презентация к уроку биологии в 7 классе Отряд Чешуекрылые, или Бабочки
Презентация к уроку биологии в 7 классе Отряд Чешуекрылые, или Бабочки Гаструляция. Дробление и гаструляция в кишечнодышащих
Гаструляция. Дробление и гаструляция в кишечнодышащих V.A. Motsart hayoti va ijodi
V.A. Motsart hayoti va ijodi Клеточные и неклеточные формы жизни
Клеточные и неклеточные формы жизни Цитология. Клеточная теория
Цитология. Клеточная теория Восприятие ЭМИ разными видами животных. Особенности электрогенерирования, электро- и магниторецепции животных
Восприятие ЭМИ разными видами животных. Особенности электрогенерирования, электро- и магниторецепции животных Мед и пчелы
Мед и пчелы Развитие жизни на Земле. Многообразие органического мира. Классификация организмов
Развитие жизни на Земле. Многообразие органического мира. Классификация организмов Эволюционное учение Чарльза Дарвина. Естественный отбор, формы естественного отбора
Эволюционное учение Чарльза Дарвина. Естественный отбор, формы естественного отбора Семейство Тыквенные - бахчевые культуры. Семейство Крестоцветные (Карустные). Лабораторная работа № 9.
Семейство Тыквенные - бахчевые культуры. Семейство Крестоцветные (Карустные). Лабораторная работа № 9. Тайны глубины морей
Тайны глубины морей Методы диагностики, морфология, микроскопия, методы окраски бактерий
Методы диагностики, морфология, микроскопия, методы окраски бактерий Развитие жизни на Земле. Эры, периоды, эпохи, века
Развитие жизни на Земле. Эры, периоды, эпохи, века Подцарство Одноклеточные. Простейшие
Подцарство Одноклеточные. Простейшие Соседи по планете живущие в океанских глубинах
Соседи по планете живущие в океанских глубинах Biologics in Rheumatology
Biologics in Rheumatology Роль печени в обменных процессах
Роль печени в обменных процессах Бактериофаги
Бактериофаги Сахарный диабет. Гормоны – производные аминокислот
Сахарный диабет. Гормоны – производные аминокислот Строение растительной, животной, грибной и бактериальной клеток под микроскопом. Лабораторная работа
Строение растительной, животной, грибной и бактериальной клеток под микроскопом. Лабораторная работа Ботаника. Салалары
Ботаника. Салалары Презентация к уроку биологии в 6 классе по теме: Способы передвижения животных. Движение растений.
Презентация к уроку биологии в 6 классе по теме: Способы передвижения животных. Движение растений. Собственно соединительные ткани
Собственно соединительные ткани Способы размножения животных. Оплодотворение
Способы размножения животных. Оплодотворение