- Мышечная ткань. Классификация, морфофункциональная характеристика

Содержание
- 2. Мышечная ткань Мышечные ткани – это группа тканей, имеющая различное происхождение и строение, но объединенная по
- 3. Функции мышечной ткани Выполняют сократительную функцию Приводят в движение рычаги скелета, Обеспечивают ритмическую деятельность миокарда и
- 4. Общие свойства мышечных тканей 1. Структурные элементы мышечных тканей (клетки, волокна) обладают удлиненной формой; 2. Наличие
- 5. Общие свойства мышечных тканей Содержат много митохондрий. Имеют запасы углеводов в виде гранул гликогена и запасы
- 6. Общие свойства мышечных тканей Сокращение происходит по принципу скольжения толстых и тонких миофиламентов друг относительно друга
- 7. Классификация Классификация мышечных тканей основана на их признаках: Строения и функции (морфофункциональная классификация) Происхождения (гистогенетическая классификация).
- 8. Морфологическая классификация Гладкие Поперечно-полосатые Скелетная Сердечная
- 9. Поперечно-полосатые мышечные ткани Сердечная мышечная ткань - из миоэпикардиальной пластинки (находящейся в составе висцерального листка спланхнотома).
- 10. Гистогенетическая классификация Соматического типа (миотомы) Целомического типа (миоэпикардиальная пластинка) Мезенхимного происхождения (стенки сосудов и внутренних органов
- 11. Происхождение мышечных тканей Гладкие (неисчерченные) мышечные ткани Мезенхимного происхождения - гладкая мышечная ткань сосудов и внутренних
- 12. Гладкая мышечная ткань Гладкая мышечная ткань образована гладкими миоцитами. Развивается из мезенхимы по следующей схеме: мезенхимная
- 13. Строение гладкого миоцита гладкие миоциты - клетки веретеновидной и звёздчатой формы (миоэпителиальные клетки) Длиной 20-500 мкм,
- 14. Строение гладкого миоцита В гладких миоцитах хорошо выражена гранулярная ЭПС и КГ. Кроме сократительной функции, могут
- 15. Строение гладкого миоцита Миофибриллы лишены регулярной организации, поэтому у клеток в нет поперечной исчерченности - термин
- 16. Гладкая мышечная ткань Несмотря на разное строение, плотные пластинки плазмолеммы и плотные тельца цитоплазмы содержат те
- 17. Гладкая мышечная ткань Толстые же (миозиновые) миофиламенты в состоянии покоя распадаются на фрагменты или даже отдельные
- 18. Строение гладкого миоцита Плазмолнмма образуют кавеолы (впячивания) – в них Са2+ Ионы Ca2+ поступают в цитозоль
- 19. Гладкая мышечная ткань Функциональные особенности: Иннервируется вегетативной нервной системой и потому не может напрямую управляться волей
- 20. Гладкая мышечная ткань
- 21. Гладкая мышечная ткань
- 22. Механизм сокращения Под влиянием нервного импульса из внешней среды в клетку с помощью кавеол и через
- 23. Механизм сокращения Толстые миофиламенты внедряются между тонкими – образуются временные миофибриллы. Миофиламенты перемещаются навстречу друг другу
- 24. Регенерация и обновление В обычных условиях новообразования гладких миоцитов не происходит. Обновление ткани совершается на субклеточном
- 25. Регенерация и обновление Это происходит двумя способами: путём гипертрофии (увеличения размеров) миоцитов и за счёт гиперплазии
- 26. Скелетная мышечная ткань Локализация скелетной мышечной ткани: образует скелетные мышцы. составляют 25-50% от общей массы тела
- 27. Образование мышечных волокон Скелетная мышечная ткань - из клеток миотома образуются промиобласты и миобласты, которые активно
- 28. Скелетная мышечная ткань Основной элемент скелетной мышечной ткани – мышечные волокна. Волокно включает 2 компонента: миосимпласт
- 29. Скелетная поперечнополосатая мышечная ткань Размеры волокон: Диаметр мышечных волокон – 50-70 мкм, что почти в 10
- 30. Скелетная поперечнополосатая мышечная ткань Мышечные волокна отличаются высокой оксифилией: они интенсивно красятся эозином в ярко-розовый цвет,
- 31. Скелетная мышечная ткань В отношении миосимпластов вместо термина "цитоплазма" используется термин "саркоплазма" (греч. sarcos – мясо).
- 32. Не надо путать три уровня нитевидных структур в мышечных тканях: мышечные волокна (истинные или функциональные) -
- 33. Поперечная исчерченность Благодаря особой укладке миофиламентов, миофибриллы имеют поперечную исчерченность: в них регулярно чередуются светлые и
- 34. Скелетная мышечная ткань
- 35. Поперечная исчерченность Поперечная исчерченность мышечного волокна обусловлена исчерченностью отдельных миофибрилл: В каждой миофибрилле закономерно чередуются светлые
- 36. Тонкие миофиламенты Тонкие, или актиновые, миофиламенты образованы глобулярным белком актином. Примерно 350 молекул последнего объединяются в
- 37. Тонкие миофиламенты Функциональная роль тропонина и тропомиозина состоит в том, что они влияют на взаимодействие актина
- 38. Тонкие миофиламенты Тонкие филаменты прикрепляются к телофрагме (т.е. к белку актинину) с обеих её сторон. Таким
- 39. САРКОМЕР
- 40. Толстые миофиламенты Толстые (миозиновые) миофиламенты образованы белком миозином. Молекула миозина состоит из нескольких пептидных цепей и
- 41. Толстые миофиламенты Толстые миофиламенты крепятся своей срединной частью к мезофрагме (образованной М-белком). Длина толстых миофиламентов равна
- 42. Саркомер Саркомер – это участок миофибриллы между двумя соседними телофрагмами. Саркомер включает два полудиска I (прилегающие
- 43. САРКОМЕР
- 44. Саркомер
- 45. Всего в саркомере – около 1400 толстых миофиламентов, что в 4 раза меньше общего количества тонких
- 46. Таким образом, упорядоченность расположения миофиламентов в миофибрилле и миофибрилл в миосимпласте поддерживается с помощью ряда опорных
- 47. Телофрагма В центре светлого диска располагается телофрагма (видимая как Z-линия) - это сетчатая пластинка из белковых
- 48. Мезофрагма В центре темного диска А находится относительно более светлая Н-зона (шириной в покое 0,5 мкм),
- 49. Промежуточные филаменты образованы в миосимпластах белком десмином, которые связывают соседние телофрагмы миофибриллы и миофибриллы с мембранными
- 50. Для передачи возбуждения от плазмолеммы к миофибриллам в миосимпластах существуют специальные мембранные структуры: Т-трубочки и L-канальцы
- 51. Т-трубочки и L-канальцы
- 52. Аккумуляция ионов Са2+ в цистернах За счет - Са2+-насосов и Са2+-каналов: Са2+-насос закачивает внутрь цистерн ионы
- 53. Механизм сокращения При возбуждении мышечного волокна в сарколемме возрастает содержание ионов Са2+. В тонких миофиламентах меняется
- 54. АДФ и фосфат вытесняются актином из связи с миозиновыми головками. Hапряжённая конформация миозиновых головок создаёт силу,
- 55. Размыкание мостиков с помощью АТФ - головки связывают новые молекулы АТФ – и это приводит к
- 56. В итоге в саркомерах миофибрилл I-диски и светлая (Н-) зона А-диска становятся тоньше, а тёмные участки
- 57. Трупное окоченение После смерти, в связи с угасанием метаболических процессов, в мышцах быстро снижается концентрация АТФ,
- 58. Типы мышечных волокон По своим физиологическим возможностям и обуславливающим их биохимическим свойствам, мышечные волокна делят на
- 59. Красные и белые волокна
- 60. Типы мышечных волокон Эти волокна в том или ином соотношении содержатся в одной и той же
- 61. Красные мышечные волокна Красные мышечные волокна (волокна I, или медленного типа) Способны к не очень интенсивной,
- 62. Красные волокна В волокнах много миоглобина - белка, запасающего О2 . Отсюда - красный цвет волокон
- 63. Красные волокна В красных волокнах многочисленны липидные включения. Высока активность ферментов окисления - в т.ч. сукцинатдегидрогеназы
- 64. Белые мышечные волокна Белые мышечные волокна (волокна II, или быстрого типа) Белые волокна способны к интенсивной,
- 65. Белые волокна Содержание миоглобина - низкое. Отсюда - светлый цвет волокон. Содержание гликогена - высокое. Анаэробный
- 66. Белые волокна Липидные включения невелики или вообще отсутствуют - поскольку жиры анаэробным способом не разрушаются. Активность
- 67. Миосателлиты миосателлитов выполняют роль камбия: у детей – при росте мышечных волокон у взрослых – в
- 68. Регенерация мышечной ткани При любом виде повреждения мышцы вначале происходит: миграция в повреждённую область нейтрофилов и
- 69. Регенерация Собственно регенерация осуществляется двумя способами - Первый способ: восстановление целостности повреждённых волокон - путём медленного
- 70. Регенерация Однако при значительном повреждении базальной мембраны мышечных волокон полного восстановления прежней структуры обычно не происходит:
- 71. Мышца как орган Если говорить о скелетных мышцах как об органах, то в них обнаруживаются другие
- 72. Мышца как орган Эндомизий - это узкие прослойки рыхлой волокнистой неоформленной соединительной ткани между мышечными волокнами.
- 73. Сердечная мышечная ткань Образует миокард - мышечную оболочку сердца и иннервируется вегетативной нервной системой. Состоит из
- 74. Сердечная мышечная ткань Основной элемент сердечной мышечной ткани – типичные кардиомиоциты Клетки цилиндрической формы, которые стыкуются
- 75. Вставочные диски Места контактов соседних кардиомиоцитов называются вставочными дисками. На световых препаратах они выглядят как тонкие
- 76. Вставочные диски Интердигитации – пальцевидные впячивания клеток друг в друга; Десмосомы – контакты, обеспечивающие более прочное
- 77. Регенерация Плохая из-за отсутствия камбия - миосателлитов или иных камбиальных клеток в сердечной мышечной ткани нет.
- 78. Разновидности кардиомиоцитов Типичные (сократительные, или рабочие) кардиомиоциты, В сердце присутствуют и другие их разновидности: Секреторные (ПНФ),
- 79. Сердечная мышечная ткань Миофибриллы имеют такую же организацию, как и в скелетной мышечной ткани, т.е. образуют
- 80. Т-трубочки и L-система В типичных кардиомиоцитах, как и в мышечных волокнах скелетных мышц, имеются специальные мембранные
- 82. Сердечная мышечная ткань В кардиомиоцитах много миоглобина и липидных капель. Гранул же гликогена относительно мало. В
- 83. Сердечная мышечная ткань Субстратами окисления служат следующие вещества: жирные кислоты, кроме того, – продукты распада последних
- 84. Сравнение скелетной и сердечной Сократительными элементами в обеих тканях являются миофибриллы, которые ориентированы вдоль длиной оси
- 85. Сравнение скелетной и сердечной Между волокнами двух поперечнополосатых тканей существует принципиальная разница: в скелетной мышечной ткани
- 86. Сравнение скелетной и сердечной мышечной ткани В скелетной и сердечной мышечных тканях, помимо АТФ, функцию аккумулятора
- 88. Скачать презентацию

Мышечная ткань
Мышечные ткани – это группа тканей, имеющая различное происхождение и
Мышечная ткань
Мышечные ткани – это группа тканей, имеющая различное происхождение и

Функции мышечной ткани
Выполняют сократительную функцию
Приводят в движение рычаги скелета,
Обеспечивают ритмическую деятельность
Функции мышечной ткани
Выполняют сократительную функцию
Приводят в движение рычаги скелета,
Обеспечивают ритмическую деятельность

Общие свойства мышечных тканей
1. Структурные элементы мышечных тканей (клетки, волокна) обладают
Общие свойства мышечных тканей
1. Структурные элементы мышечных тканей (клетки, волокна) обладают

Общие свойства мышечных тканей
Содержат много митохондрий.
Имеют запасы углеводов в виде
Общие свойства мышечных тканей
Содержат много митохондрий.
Имеют запасы углеводов в виде

Общие свойства мышечных тканей
Сокращение происходит по принципу скольжения толстых и тонких
Общие свойства мышечных тканей
Сокращение происходит по принципу скольжения толстых и тонких

Классификация
Классификация мышечных тканей основана на их признаках:
Строения и функции (морфофункциональная
Классификация
Классификация мышечных тканей основана на их признаках:
Строения и функции (морфофункциональная

Морфологическая классификация
Гладкие
Поперечно-полосатые
Скелетная
Сердечная
Морфологическая классификация
Гладкие
Поперечно-полосатые
Скелетная
Сердечная

Поперечно-полосатые
мышечные ткани
Сердечная мышечная ткань - из миоэпикардиальной пластинки (находящейся
Поперечно-полосатые
мышечные ткани
Сердечная мышечная ткань - из миоэпикардиальной пластинки (находящейся

Гистогенетическая классификация
Соматического типа (миотомы)
Целомического типа (миоэпикардиальная пластинка)
Мезенхимного происхождения (стенки сосудов и
Гистогенетическая классификация
Соматического типа (миотомы)
Целомического типа (миоэпикардиальная пластинка)
Мезенхимного происхождения (стенки сосудов и

Происхождение мышечных тканей
Гладкие (неисчерченные) мышечные ткани
Мезенхимного происхождения - гладкая мышечная
Происхождение мышечных тканей
Гладкие (неисчерченные) мышечные ткани
Мезенхимного происхождения - гладкая мышечная

Гладкая мышечная ткань
Гладкая мышечная ткань образована гладкими миоцитами.
Развивается из мезенхимы
Гладкая мышечная ткань
Гладкая мышечная ткань образована гладкими миоцитами.
Развивается из мезенхимы

Строение гладкого миоцита
гладкие миоциты - клетки веретеновидной и звёздчатой формы (миоэпителиальные
Строение гладкого миоцита
гладкие миоциты - клетки веретеновидной и звёздчатой формы (миоэпителиальные

Строение гладкого миоцита
В гладких миоцитах хорошо выражена гранулярная ЭПС и КГ.
Строение гладкого миоцита
В гладких миоцитах хорошо выражена гранулярная ЭПС и КГ.

Строение гладкого миоцита
Миофибриллы лишены регулярной организации, поэтому у клеток в нет
Строение гладкого миоцита
Миофибриллы лишены регулярной организации, поэтому у клеток в нет

Гладкая мышечная ткань
Несмотря на разное строение, плотные пластинки плазмолеммы и плотные
Гладкая мышечная ткань
Несмотря на разное строение, плотные пластинки плазмолеммы и плотные

Гладкая мышечная ткань
Толстые же (миозиновые) миофиламенты в состоянии покоя распадаются на
Гладкая мышечная ткань
Толстые же (миозиновые) миофиламенты в состоянии покоя распадаются на

Строение гладкого миоцита
Плазмолнмма образуют кавеолы (впячивания) – в них Са2+
Ионы Ca2+
Строение гладкого миоцита
Плазмолнмма образуют кавеолы (впячивания) – в них Са2+
Ионы Ca2+

Гладкая мышечная ткань
Функциональные особенности:
Иннервируется вегетативной нервной системой и потому не
Гладкая мышечная ткань
Функциональные особенности:
Иннервируется вегетативной нервной системой и потому не
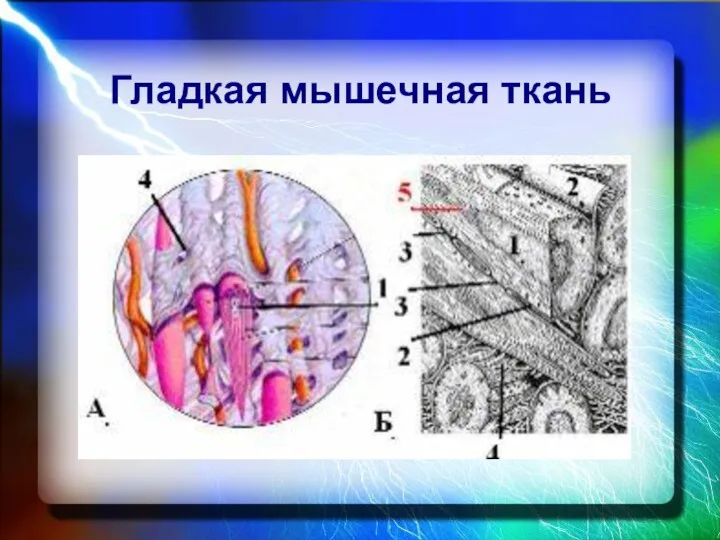
Гладкая мышечная ткань
Гладкая мышечная ткань

Гладкая мышечная ткань
Гладкая мышечная ткань

Механизм сокращения
Под влиянием нервного импульса из внешней среды в клетку с
Механизм сокращения
Под влиянием нервного импульса из внешней среды в клетку с

Механизм сокращения
Толстые миофиламенты внедряются между тонкими – образуются временные миофибриллы.
Миофиламенты перемещаются
Механизм сокращения
Толстые миофиламенты внедряются между тонкими – образуются временные миофибриллы.
Миофиламенты перемещаются

Регенерация и обновление
В обычных условиях новообразования гладких миоцитов не происходит.
Обновление
Регенерация и обновление
В обычных условиях новообразования гладких миоцитов не происходит.
Обновление

Регенерация и обновление
Это происходит двумя способами:
путём гипертрофии (увеличения размеров) миоцитов и
Регенерация и обновление
Это происходит двумя способами:
путём гипертрофии (увеличения размеров) миоцитов и

Скелетная мышечная ткань
Локализация скелетной мышечной ткани:
образует скелетные мышцы.
составляют 25-50% от
Скелетная мышечная ткань
Локализация скелетной мышечной ткани:
образует скелетные мышцы.
составляют 25-50% от

Образование мышечных волокон
Скелетная мышечная ткань - из клеток миотома образуются промиобласты
Образование мышечных волокон
Скелетная мышечная ткань - из клеток миотома образуются промиобласты

Скелетная мышечная ткань
Основной элемент скелетной мышечной ткани – мышечные волокна.
Волокно
Скелетная мышечная ткань
Основной элемент скелетной мышечной ткани – мышечные волокна.
Волокно

Скелетная поперечнополосатая мышечная ткань
Размеры волокон:
Диаметр мышечных волокон – 50-70 мкм,
Скелетная поперечнополосатая мышечная ткань
Размеры волокон:
Диаметр мышечных волокон – 50-70 мкм,

Скелетная поперечнополосатая мышечная ткань
Мышечные волокна отличаются
высокой оксифилией: они интенсивно красятся
Скелетная поперечнополосатая мышечная ткань
Мышечные волокна отличаются
высокой оксифилией: они интенсивно красятся

Скелетная мышечная ткань
В отношении миосимпластов вместо термина "цитоплазма" используется термин "саркоплазма"
Скелетная мышечная ткань
В отношении миосимпластов вместо термина "цитоплазма" используется термин "саркоплазма"

Не надо путать три уровня нитевидных структур в мышечных тканях:
мышечные волокна
Не надо путать три уровня нитевидных структур в мышечных тканях:
мышечные волокна

Поперечная исчерченность
Благодаря особой укладке миофиламентов, миофибриллы имеют поперечную исчерченность: в них
Поперечная исчерченность
Благодаря особой укладке миофиламентов, миофибриллы имеют поперечную исчерченность: в них

Скелетная мышечная ткань
Скелетная мышечная ткань

Поперечная исчерченность
Поперечная исчерченность мышечного волокна обусловлена исчерченностью отдельных миофибрилл:
В каждой миофибрилле
Поперечная исчерченность
Поперечная исчерченность мышечного волокна обусловлена исчерченностью отдельных миофибрилл:
В каждой миофибрилле

Тонкие миофиламенты
Тонкие, или актиновые, миофиламенты образованы глобулярным белком актином.
Примерно 350
Тонкие миофиламенты
Тонкие, или актиновые, миофиламенты образованы глобулярным белком актином.
Примерно 350

Тонкие миофиламенты
Функциональная роль тропонина и тропомиозина состоит в том, что они
Тонкие миофиламенты
Функциональная роль тропонина и тропомиозина состоит в том, что они

Тонкие миофиламенты
Тонкие филаменты прикрепляются к телофрагме (т.е. к белку актинину) с
Тонкие миофиламенты
Тонкие филаменты прикрепляются к телофрагме (т.е. к белку актинину) с

САРКОМЕР
САРКОМЕР

Толстые миофиламенты
Толстые (миозиновые) миофиламенты образованы белком миозином.
Молекула миозина состоит из нескольких
Толстые миофиламенты
Толстые (миозиновые) миофиламенты образованы белком миозином.
Молекула миозина состоит из нескольких

Толстые миофиламенты
Толстые миофиламенты крепятся своей срединной частью к мезофрагме (образованной М-белком).
Длина
Толстые миофиламенты
Толстые миофиламенты крепятся своей срединной частью к мезофрагме (образованной М-белком).
Длина

Саркомер
Саркомер – это участок миофибриллы между двумя соседними телофрагмами.
Саркомер включает
Саркомер
Саркомер – это участок миофибриллы между двумя соседними телофрагмами.
Саркомер включает

САРКОМЕР
САРКОМЕР

Саркомер
Саркомер

Всего в саркомере – около 1400 толстых миофиламентов, что в 4
Всего в саркомере – около 1400 толстых миофиламентов, что в 4

Таким образом, упорядоченность расположения миофиламентов в миофибрилле и миофибрилл в миосимпласте
Таким образом, упорядоченность расположения миофиламентов в миофибрилле и миофибрилл в миосимпласте

Телофрагма
В центре светлого диска располагается телофрагма (видимая как Z-линия) - это
Телофрагма
В центре светлого диска располагается телофрагма (видимая как Z-линия) - это

Мезофрагма
В центре темного диска А находится относительно более светлая Н-зона (шириной
Мезофрагма
В центре темного диска А находится относительно более светлая Н-зона (шириной

Промежуточные филаменты образованы в миосимпластах белком десмином, которые связывают соседние телофрагмы
Промежуточные филаменты образованы в миосимпластах белком десмином, которые связывают соседние телофрагмы

Для передачи возбуждения от плазмолеммы к миофибриллам в миосимпластах существуют специальные
Для передачи возбуждения от плазмолеммы к миофибриллам в миосимпластах существуют специальные

Т-трубочки и L-канальцы
Т-трубочки и L-канальцы

Аккумуляция ионов Са2+ в цистернах
За счет - Са2+-насосов и Са2+-каналов:
Са2+-насос
Аккумуляция ионов Са2+ в цистернах
За счет - Са2+-насосов и Са2+-каналов:
Са2+-насос

Механизм сокращения
При возбуждении мышечного волокна в сарколемме возрастает содержание ионов Са2+.
Механизм сокращения
При возбуждении мышечного волокна в сарколемме возрастает содержание ионов Са2+.

АДФ и фосфат вытесняются актином из связи с миозиновыми головками.
Hапряжённая конформация
АДФ и фосфат вытесняются актином из связи с миозиновыми головками.
Hапряжённая конформация

Размыкание мостиков с помощью АТФ - головки связывают новые молекулы АТФ
Размыкание мостиков с помощью АТФ - головки связывают новые молекулы АТФ

В итоге в саркомерах миофибрилл
I-диски и светлая (Н-) зона А-диска становятся
В итоге в саркомерах миофибрилл
I-диски и светлая (Н-) зона А-диска становятся

Трупное окоченение
После смерти, в связи с угасанием метаболических процессов, в мышцах
Трупное окоченение
После смерти, в связи с угасанием метаболических процессов, в мышцах

Типы мышечных волокон
По своим физиологическим возможностям и обуславливающим их биохимическим свойствам,
Типы мышечных волокон
По своим физиологическим возможностям и обуславливающим их биохимическим свойствам,

Красные и белые волокна
Красные и белые волокна

Типы мышечных волокон
Эти волокна в том или ином соотношении содержатся в
Типы мышечных волокон
Эти волокна в том или ином соотношении содержатся в

Красные мышечные волокна
Красные мышечные волокна
(волокна I, или медленного типа)
Способны
Красные мышечные волокна
Красные мышечные волокна
(волокна I, или медленного типа)
Способны

Красные волокна
В волокнах много миоглобина - белка, запасающего О2 .
Отсюда
Красные волокна
В волокнах много миоглобина - белка, запасающего О2 .
Отсюда

Красные волокна
В красных волокнах многочисленны липидные включения.
Высока активность ферментов окисления -
Красные волокна
В красных волокнах многочисленны липидные включения.
Высока активность ферментов окисления -

Белые мышечные волокна
Белые мышечные волокна (волокна II, или быстрого типа)
Белые волокна
Белые мышечные волокна
Белые мышечные волокна (волокна II, или быстрого типа)
Белые волокна

Белые волокна
Содержание миоглобина - низкое.
Отсюда - светлый цвет волокон.
Содержание гликогена
Белые волокна
Содержание миоглобина - низкое.
Отсюда - светлый цвет волокон.
Содержание гликогена

Белые волокна
Липидные включения невелики или вообще отсутствуют - поскольку жиры анаэробным
Белые волокна
Липидные включения невелики или вообще отсутствуют - поскольку жиры анаэробным

Миосателлиты
миосателлитов выполняют роль камбия:
у детей – при росте мышечных волокон
у
Миосателлиты
миосателлитов выполняют роль камбия:
у детей – при росте мышечных волокон
у

Регенерация мышечной ткани
При любом виде повреждения мышцы вначале происходит:
миграция в повреждённую
Регенерация мышечной ткани
При любом виде повреждения мышцы вначале происходит:
миграция в повреждённую

Регенерация
Собственно регенерация осуществляется двумя способами -
Первый способ: восстановление целостности повреждённых
Регенерация
Собственно регенерация осуществляется двумя способами -
Первый способ: восстановление целостности повреждённых

Регенерация
Однако при значительном повреждении базальной мембраны мышечных волокон полного восстановления прежней
Регенерация
Однако при значительном повреждении базальной мембраны мышечных волокон полного восстановления прежней

Мышца как орган
Если говорить о скелетных мышцах как об органах, то
Мышца как орган
Если говорить о скелетных мышцах как об органах, то

Мышца как орган
Эндомизий - это узкие прослойки рыхлой волокнистой неоформленной соединительной
Мышца как орган
Эндомизий - это узкие прослойки рыхлой волокнистой неоформленной соединительной

Сердечная мышечная ткань
Образует миокард - мышечную оболочку сердца и иннервируется вегетативной
Сердечная мышечная ткань
Образует миокард - мышечную оболочку сердца и иннервируется вегетативной

Сердечная мышечная ткань
Основной элемент сердечной мышечной ткани – типичные кардиомиоциты
Клетки цилиндрической
Сердечная мышечная ткань
Основной элемент сердечной мышечной ткани – типичные кардиомиоциты
Клетки цилиндрической

Вставочные диски
Места контактов соседних кардиомиоцитов называются вставочными дисками.
На световых препаратах
Вставочные диски
Места контактов соседних кардиомиоцитов называются вставочными дисками.
На световых препаратах

Вставочные диски
Интердигитации – пальцевидные впячивания клеток друг в друга;
Десмосомы – контакты,
Вставочные диски
Интердигитации – пальцевидные впячивания клеток друг в друга;
Десмосомы – контакты,

Регенерация
Плохая из-за отсутствия камбия - миосателлитов или иных камбиальных клеток в
Регенерация
Плохая из-за отсутствия камбия - миосателлитов или иных камбиальных клеток в

Разновидности кардиомиоцитов
Типичные (сократительные, или рабочие) кардиомиоциты,
В сердце присутствуют и другие
Разновидности кардиомиоцитов
Типичные (сократительные, или рабочие) кардиомиоциты,
В сердце присутствуют и другие

Сердечная мышечная ткань
Миофибриллы имеют такую же организацию, как и в скелетной
Сердечная мышечная ткань
Миофибриллы имеют такую же организацию, как и в скелетной

Т-трубочки и L-система
В типичных кардиомиоцитах, как и в мышечных волокнах скелетных
Т-трубочки и L-система
В типичных кардиомиоцитах, как и в мышечных волокнах скелетных


Сердечная мышечная ткань
В кардиомиоцитах много миоглобина и липидных капель.
Гранул же
Сердечная мышечная ткань
В кардиомиоцитах много миоглобина и липидных капель.
Гранул же

Сердечная мышечная ткань
Субстратами окисления служат следующие вещества:
жирные кислоты, кроме того, –
Сердечная мышечная ткань
Субстратами окисления служат следующие вещества:
жирные кислоты, кроме того, –

Сравнение скелетной и сердечной
Сократительными элементами в обеих тканях являются миофибриллы,
Сравнение скелетной и сердечной
Сократительными элементами в обеих тканях являются миофибриллы,

Сравнение скелетной и сердечной
Между волокнами двух поперечнополосатых тканей существует принципиальная
Сравнение скелетной и сердечной
Между волокнами двух поперечнополосатых тканей существует принципиальная

Сравнение скелетной и сердечной мышечной ткани
В скелетной и сердечной мышечных тканях,
Сравнение скелетной и сердечной мышечной ткани
В скелетной и сердечной мышечных тканях,
 Устройство увеличительных приборов и правила работы с ними
Устройство увеличительных приборов и правила работы с ними Виды движений в биомеханике. Лекция 5
Виды движений в биомеханике. Лекция 5 Стати собак
Стати собак Ткани. 4 типа тканей
Ткани. 4 типа тканей Введение в физиологию животных
Введение в физиологию животных Гистология органов пищеварения
Гистология органов пищеварения Селекция растений
Селекция растений Природа и сущность языка
Природа и сущность языка Cultural diffusion in humans and animals
Cultural diffusion in humans and animals Тип Хордовые
Тип Хордовые Головоногие моллюски
Головоногие моллюски Функции тонкого и толстого кишечника. Всасывание. Барьерная роль печени. Аппендицит
Функции тонкого и толстого кишечника. Всасывание. Барьерная роль печени. Аппендицит Многоствольные формы
Многоствольные формы Cells. Muscle cells
Cells. Muscle cells Физиология сна и сновидений
Физиология сна и сновидений Модификационная изменчивость
Модификационная изменчивость Загальна характеристика ферментів
Загальна характеристика ферментів Биотехнология в животноводстве
Биотехнология в животноводстве Общая характеристика царства животные
Общая характеристика царства животные Genetic-test. Биометрическое тестирование способностей человека по типам узоров на подушечках пальцев
Genetic-test. Биометрическое тестирование способностей человека по типам узоров на подушечках пальцев Террариум и его обитатели
Террариум и его обитатели Типы клеточной гибели
Типы клеточной гибели Биологические ритмы. Сон и его значение
Биологические ритмы. Сон и его значение Индивидуальное развитие цветковых растений
Индивидуальное развитие цветковых растений Популяция. Генетический состав популяций
Популяция. Генетический состав популяций Викторина о животных
Викторина о животных Профилактика и лечение наследственных заболеваний
Профилактика и лечение наследственных заболеваний Сера и её роль в организме человека
Сера и её роль в организме человека