- Мышечные ткани. Классификация

Содержание
- 2. МЫШЕЧНЫЕ ТКАНИ (гистогенетическая классификация) Мезодермальная Миотомная – скелетная Целомическая - сердечная Мезенхимная - гладкая Эктодермальная Кожноэктодермальная
- 3. РАЗВИТИЕ МЫШЕЧНОЙ ТКАНИ МИОТОМЫ- СКЕЛЕТНАЯ НЕЙРОЭКТОДЕРМА- МИОНЕЙРАЛЬНАЯ МЕЗЕНХИМНАЯ- ГЛАДКАЯ ВИСЦЕРАЛЬНЫЙ СПЛАНХНОТОМ – МИОКАРД
- 4. Классификация мышечных тканей (по Р.К. Данилову) МЫШЕЧНЫЕ ТКАНИ Мезодермальные Эктодермальные 1. Спланхномезодермальная (висцеральная, сосудистая) 2. Миотомная
- 5. Классификация миоидных тканей (по Р.К. Данилову) Миоидные клетки Мезодермальные Спланхномезенхимные (миофибробласты, миоидные клетки семенных канальцев, миоидные
- 6. МЫШЕЧНАЯ ТКАНЬ ПОПЕРЕЧНОПОЛОСАТАЯ ГЛАДКАЯ СКЕЛЕТНАЯ СЕРДЕЧНАЯ ПРОИЗВОЛЬНАЯ НЕПРОИЗВОЛЬНАЯ
- 7. У высших позвоночных по генетическим признакам И.Н.Борисов выделяет две группы мышечных тканей: целомические (мезодермальные) и нейродермальные.
- 8. КРАТКАЯ ХАРАКТЕРИСТИКА ТРЕХ РАЗНОВИДНОСТЕЙ МЫШЕЧНЫХ ТКАНЕЙ
- 9. СОМАТИЧЕКАЯ (СКЕЛЕТНАЯ) МЫШЕЧНАЯ ТКАНЬ ЛОКОМОТОРНЫЕ НЕЛОКОМОТОРНЫЕ (ВИСЦЕРАЛЬНЫЕ) МЫШЦЫ, ПРИКРЕПЛЯЮЩИЕСЯ К СКЕЛЕТУ: КОСТЯМ, СУХОЖИЛИЯМ. МЫШЕЧНЫЕ ВОЛОКНА, СВЯЗАННЫЕ
- 10. СОСУДИСТО- НЕРВНЫЙ ПУЧОК ЭПИМИЗИЙ ПЕРИМИЗИЙ ЭНДОМИЗИЙ МЫШЕЧНЫЕ ВОЛОКНА МЫШЦА, КАК ОРГАН
- 11. Индивидуальное мышечное волокно окружено плазмолеммой, называемой сарколеммой, которую покрывает базальная мембрана (гликокаликс) толщиной 30-50 нм. Гликокаликс
- 12. ФУНКЦИОНАЛЬНЫЕ АППАРАТЫ МИОСИМПЛАСТА 1. СОКРАТИТЕЛЬНЫЙ (МИОФИБРИЛЛЫ); 2. ПЕРЕДАЧИ ВОЗБУЖДЕНИЯ (САРКОТУБУЛЯРНАЯ СИСТЕМА); 3. ОПОРНЫЙ (ЭЛЕМЕНТЫ ЦИТОСКЕЛЕТА, САРКОЛЕММА,
- 13. Строение поперечнополосатой мускулатуры САРКОПЛАЗМАТИЧЕСКАЯ СЕТЬ И Т-СИСТЕМЫ Гликокаликс (базальная мембрана)
- 14. Каждая миофибрилла содержит около 1500 толстых и 3000 тонких нитей. Поперечная исчерченность скелетного мышечного волокна определяется
- 15. Строение молекулы миозина ГОЛОВКИ МОЛЕКУЛЫ МИОЗИНА АГРЕГАЦИЯ МОЛЕКУЛЫ МИОЗИНА ГОЛОВКИ МИОЗИНА ХВОСТОВЫЕ НИТИ МИОЗИНА ШАРНИРНЫЙ УЧАСТОК
- 16. Актин (спираль из глобулярных молекул) Тропониновый комплекс тропонин образует глобулярный комплекс, состоящий из тропонина I, С,
- 17. А-диск М-линия ядро I-диск Z-линия М-линия Белки М-линии изозим креатиназа (является преобразователем АДФ в АТФ), М-белок,
- 18. УЛЬТРАСТРУКТУРА Z-ЛИНИИ Z-ФИЛАМЕНТЫ Z-МАТРИКС (белок аморфин) АКТИНОВЫЕ МИОФИЛАМЕНТЫ ОПОРНЫЙ АППАРАТ Z ЛИНИЙ АЛЬФА-АКТИНИН, ФИЛАМИН, СПЕКТРИН, ВИНКУЛИН
- 19. МИОФИБРИЛЛА САРКОПЛАЗМАТИЧЕСКАЯ СЕТЬ ТРИАДА Z-ЛИНИЯ САРКОЛЕММА I-диск А-ДИСК ТЕРМИНАЛЬНЫЕ ЦИСТЕРНЫ Т-ТРУБОЧКИ Саркотубулярная система или аппарат передачи
- 20. СХЕМА МЕХАНИЗМОВ ПЕРЕДАЧИ ВОЗБУЖДЕНИЯ И ВЫБРОСА КАЛЬЦИЯ ИЗ САРКОПЛАЗМАТИЧЕСКОЙ СЕТИ В ПОПЕРЕЧНО-ПОЛОСАТОЙ МЫШЕЧНОЙ ТКАНИ
- 21. Связывание Ca2+ тонкими нитями. В покое взаимодействие тонких и толстых нитей невозможно, т.к. миозин-связывающие участки F‑актина
- 22. Са ЗАВИСИМЫЙ МЕХАНИЗМ РЕГУЛЯЦИИ ВЗАИМОДЕЙСТВИЯ АКТИНА С МИОЗИНОМ ТРОПОМИОЗИН ТРОПОНИН АДФ+Ф УЧАСТКИ СВЯЗЫВАНИЯ МИОЗИНА актин
- 23. ПОСЛЕДОВАТЕЛЬНОСТЬ МЫШЕЧНОГО СОКРАЩЕНИЯ: ДЕПОЛЯРИЗАЦИЯ ПОСТСИНАПТИЧЕСКОЙ МЕМБРАНЫ И ГЕНЕРАЦИЯ ПОТЕНЦИАЛА ДЕЙСТВИЯ; РАСПРОСТРАНЕНИЕ ПОТЕНЦИАЛА ДЕЙСТВИЯ ПО ПЛАЗМОЛЕММЕ МЫШЕЧНОГО
- 24. ТИПЫ МЫШЕЧНЫХ ВОЛОКОН ФАЗНЫЕ ТИП I – КРАСНЫЕ, МЕДЛЕННЫЕ, УСТОЙЧИВЫЕ К УТОМЛЕНИЮ, С НЕБОЛЬШОЙ СИЛОЙ СОКРАЩЕНИЯ,
- 25. ТИПЫ МЫШЕЧНЫХ ВОЛОКОН
- 26. РЕГЕНЕРАЦИЯ ПОПЕРЕЧНОПОЛОСАТОЙ МУСКУЛАТУРЫ
- 27. СЕРДЕЧНАЯ (ИСЧЕРЧЕННАЯ) МЫШЕЧНАЯ ТКАНЬ
- 28. СТРОЕНИЕ СЕРДЕЧНОЙ МЫШЕЧНОЙ ТКАНИ кардтомиоцит сосуд вставочный диск кардтомиоцит вставочный диск
- 29. ТИПЫ КАРДИОМИОЦИТОВ СОКРАТИТЕЛЬНЫЕ (РАБОЧИЕ) ПРОВОДЯЩИЕ Р-клетки (пейсмекерские), Промежуточные (переходные), Клетки-волокна Пуркинье СЕКРЕТОРНЫЕ (ЭНДОКРИННЫЕ) Натрий уретический фактор
- 30. МИТОХОНДРИАЛЬНАЯ СИСТЕМА КАРДИОМИОЦИТОВ межмитохондриальный контаткт межмитохондриальный контаткт миофибриллы миофибриллы
- 31. САРКОПЛАЗМАТИЧЕСКИЙ РЕТИКУЛУМ И Т-КАНАЛЫ РАБОЧИХ КАРДИОМИОЦИТОВ СР ТЦ ТЦ- терминальные цистерны СР-саркоплазматический ретикулум Т Т Т-каналы
- 32. ГЛАДКИЕ МИОЦИТЫ
- 33. КЛАССИФИКАЦИЯ ГЛАДКОЙ МУСКУЛАТУРЫ: ПО СКОРОСТИ СОКРАЩЕНИЯ ФАЗНАЯ – в матке, тонкой кишке, портальной вене; ТОНИЧЕСКАЯ –
- 34. Различают контрактильные, секреторные и промежуточные леомиоциты. Контрактильные ГМ в них много элементов гладкой ЭПС, слабо развита
- 35. ГЛАДКИЕ МИОЦИТЫ ПЛОТНЫЕ ТЕЛЬЦА ПЛОТНЫЕ ПЛАСТИНКИ МИКРОФИЛАМЕНТЫ ЯДРО Плотное тельце – аналог Z линий связано с
- 36. К рождению скелетные мышцы составляют около 25 % массы тела. Самыми толстыми оказываются волокна в диафрагме,
- 37. От 3-х до 7 лет количество миофибрилл в волокне увеличивается по сравнению с новорожденным в 15—20
- 38. Наиболее интенсивно она начинает развиваться в старшем школьном возрасте между 15 и 18 годами, когда происходит
- 39. Хорошо пользоваться скелетной нелокомоторной и тонической мускулатурой! Спасибо за внимание!
- 41. Скачать презентацию
 Мир животных, который окружает нас. 4 класс
Мир животных, который окружает нас. 4 класс Презентация Птицы водоёмов и побережий.
Презентация Птицы водоёмов и побережий. Почему осенью осыпаются листья
Почему осенью осыпаются листья Белки. Качественный состав белков
Белки. Качественный состав белков Породы лошадей
Породы лошадей Земноводные
Земноводные Питательные среды для культивирования и их классификация
Питательные среды для культивирования и их классификация Насекомые – вредители сада
Насекомые – вредители сада Направления эволюции
Направления эволюции Формы обучения биологии
Формы обучения биологии Систематика живого мира
Систематика живого мира Селекция растений
Селекция растений Классификация вирусов по Балтимору
Классификация вирусов по Балтимору Поняття про хвороби рослин. Неінфекційні та інфекційні хвороби рослин. Лекція №2
Поняття про хвороби рослин. Неінфекційні та інфекційні хвороби рослин. Лекція №2 Комары. Интересные факты
Комары. Интересные факты Живые изгороди
Живые изгороди Интегративная деятельность организма. Рефлексы
Интегративная деятельность организма. Рефлексы Внутреннее строение лягушки
Внутреннее строение лягушки Внутренняя среда организма, кровь
Внутренняя среда организма, кровь 20190315_tip_ploskie_chervi
20190315_tip_ploskie_chervi Изучение влияния внешних факторов на проращивание семян яблони
Изучение влияния внешних факторов на проращивание семян яблони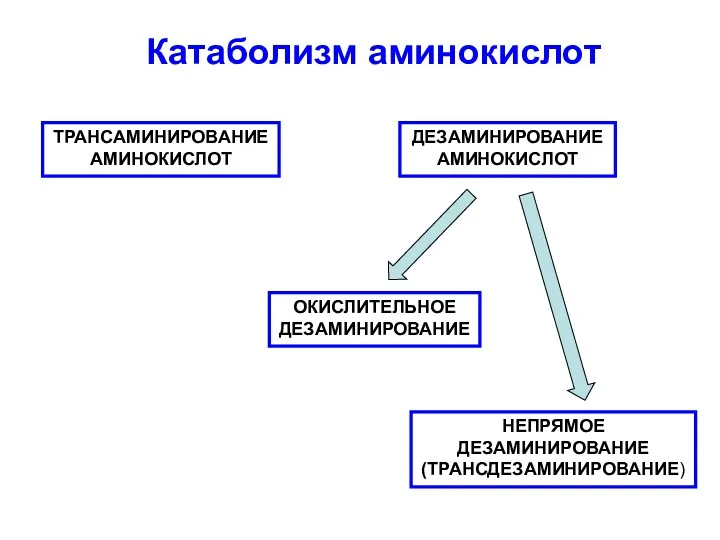 Катаболизм аминокислот
Катаболизм аминокислот Проект Загрязнение воздуха
Проект Загрязнение воздуха Общая Характеристика Царства растения. Низшие растения
Общая Характеристика Царства растения. Низшие растения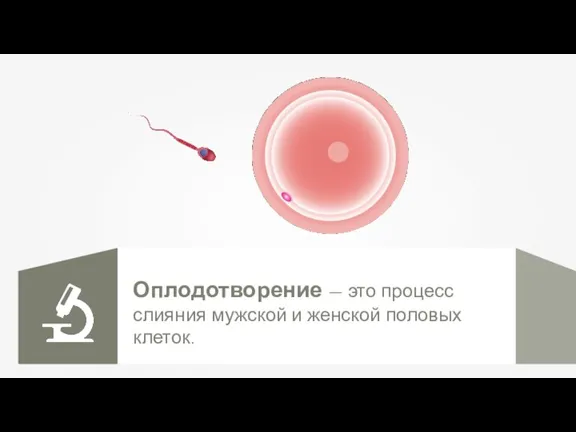 Оплодотворение. Способы оплодотворения
Оплодотворение. Способы оплодотворения Жизнь и гибель динозавров на земле
Жизнь и гибель динозавров на земле Российские учёные-биологи. Их вклад в науку.
Российские учёные-биологи. Их вклад в науку. Первая помощь при повреждении скелета
Первая помощь при повреждении скелета